LncRNA NEAT1在结核分枝杆菌感染肺泡Ⅱ型细胞中表达及作用机制
2022-06-27王玲李君丁国祥青海省第四人民医院呼吸五科青海西宁80000检验科
王玲 李君 丁国祥 (青海省第四人民医院 呼吸五科,青海 西宁 80000;检验科)
肺结核俗称肺痨,是结核病中最常见的一种慢性消耗性传染疾病〔1〕。资料显示,全球每年肺结核新发患者约800万人,年死亡人数约140万人,且其发病率与死亡率呈逐年上升趋势,已成为严重威胁人类健康的重要疾病之一〔2〕。临床显示,肺结核主要是由结核分枝杆菌引起的,但结核分枝杆菌的致病机制至今仍未完全清楚〔3〕。长链非编码RNA(LncRNA)在真核生物体内表达广泛,能够参与机体遗传调节、转录、翻译等生物学过程〔4〕。LncRNA核富集转录体(NEAT)1是一种在细胞核内表达的LncRNA〔5〕。近期研究表明,肺结核患者外周血单个核细胞中LncRNA NEAT1表达上调,且其表达情况与肺结核疾病的进展及转归有关〔6〕。但LncRNA NEAT1在结核分枝杆菌致病过程中的生物学调节机制目前尚鲜有报道,本研究为进一步分析LncRNA NEAT1与结核分枝杆菌感染的过程,通过分析LncRNA NEAT1在结核分枝杆菌感染肺泡Ⅱ型细胞中的表达情况,旨在探讨其作用机制。
1 材料和方法
1.1细胞与主要试剂 肺泡Ⅱ型细胞RLE-6TN(编号为BNCC337708)购自北京北纳创联生物技术研究院;卡介苗(BCG)疫苗冻干粉购自北京生物制品研究所;RPMI1640完全培养液、罗氏培养基购自武汉纯度生物科技有限公司;抗Toll样受体(TLR)4、髓样分化因子(MyD)88、肿瘤坏死因子受体相关因子(TRAF)6、磷酸化(p)-核转录因子(NF)-κB抗体及抗β-actin抗体购自赛默飞世尔科技(中国)有限公司;RNA提试剂盒、蛋白提取试剂盒、反转录试剂盒、qPCR试剂盒购自北京索莱宝科技有限公司。
1.2方法
1.2.1细胞培养和分组 将RLE-6TN细胞复苏后,转入RPMI1640完全培养液中,置于37℃、5%CO2饱和湿度的培养箱中进行培养,待细胞融合度达到90%时,弃去培养液,用1 ml(37℃)磷酸盐缓冲液(PBS)冲洗1次,胰酶消化后,加入完全培养液继续培养,每2~3 d更换1次培养液,取对数期的细胞进行后续实验。
1.2.2细菌的制备 BCG疫苗每支用稀释液进行稀释,接种于罗氏斜面培养基上,培养3 w后,取生长良好的BCG于匀浆器中,加少量生理盐水(0.05% Tween-80)研磨,以RPMI1640完全培养液稀释成菌悬液,调整细菌浓度(5 mg/ml)。
1.2.3细菌感染 细胞取对数生长期细胞用,培养至6 孔板,胰蛋白酶消化后,RPMI1640完全培养液制备细胞悬液,待细胞融合度至50%时,弃去培养液,根据感染复数(MOI),细菌数与细胞数之比的不同,加入终浓度为0.0、0.1、1.0、10.0 MOI重悬菌的RPMI1640完全培养液,每组细胞设置3组重复,将细胞置于(37℃、5%CO2)饱和湿度的培养箱中培养,采用直接涂片抗酸染色镜检,结核杆菌呈亮红色,结核杆菌数目≥3条/视野为细胞培养成功。分别在6、12、24 h时取对照组与重悬菌处理组的RNA、蛋白质和培养液上清,以备后续检测。
1.2.4qPCR检测 NEAT1、miR-146a、TLRs信号通路相关分子及炎症因子白细胞介素(IL)-6、IL-8、肿瘤坏死因子(TNF)-α mRNA的水平取上述各样本提取RNA,检测RNA浓度及纯度,采用反转录试剂盒反转录得cDNA,采用qPCR试剂盒进行NEAT1、miR-146a及TLRs信号通路相关分子mRNA相对定量分析,反应体系20 μl,反应条件为95℃ 10 s、60℃ 20 s、70℃ 10 s,共40个循环,所有样品重复测定3次。以β-actin作为内参(miR-146a内参为U6),采用2-ΔΔCt法分析目的基因的相对表达水平,内参及目的基因的引物序列见表1。
1.2.5Western印迹检测TLRs信号通路及炎症相关蛋白的水平 采用蛋白提取试剂盒提取上述样本总蛋白,Lowry法进行蛋白定量,绘制标准曲线,并计算样品实际浓度,用PBS将蛋白稀释至同一浓度,取30 μg蛋白样品进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE),常规转膜,封闭后,加入Ⅰ抗溶液,4℃孵育过夜,第2天室温平衡30 min,用TTBS(0.1% Tween-20)洗膜3次,每次10 min,加入相应Ⅱ抗,在杂交盒中,室温摇床孵育1.5 h,PBS洗膜2次,电化学发光(ECL)显色,Quantity One凝胶成像分析软件拍照并分析各蛋白相对表达量。每组均设3组重复,以β-actin作为内参照。

表1 RT-qPCR引物序列
1.3统计学方法 采用SPSS21.0软件进行方差分析、SNK-q检验。
2 结 果
2.1LncRNA NEAT1在结核分枝杆菌感染肺泡Ⅱ型RLE-6TN细胞中的表达情况 在相同MOI条件下,24 h时RLE-6TN细胞中NEAT1的表达水平显著高于0、6、12 h时(P<0.05),12 h时RLE-6TN细胞中NEAT1的表达水平显著高于0、6 h时(P<0.05),6 h时RLE-6TN细胞中NEAT1的表达水平显著高于0 h时(P<0.05)。在相同时间条件下,10.0 MOI组肺泡Ⅱ型细胞中NEAT1的表达水平显著高于0.0 MOI组、0.1 MOI组、1.0 MOI组(P<0.05),1.0 MOI组的RLE-6TN细胞中NEAT1的表达水平显著高于0.0 MOI组和0.1 MOI组(P<0.05);0.1 MOI组的RLE-6TN细胞中NEAT1的表达水平显著高于0.0 MOI组(P<0.05),见表2。
2.2生物信息学软件预测miR-146a是NEAT1下游的靶基因 SatrBase搜索生物信息学预测结果显示,miR-146a是NEAT1下游的直接靶标,见图1。
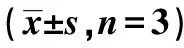
表2 LncRNA NEAT1在结核分枝杆菌感染不同时间点RLE-6TN细胞中的表达情况
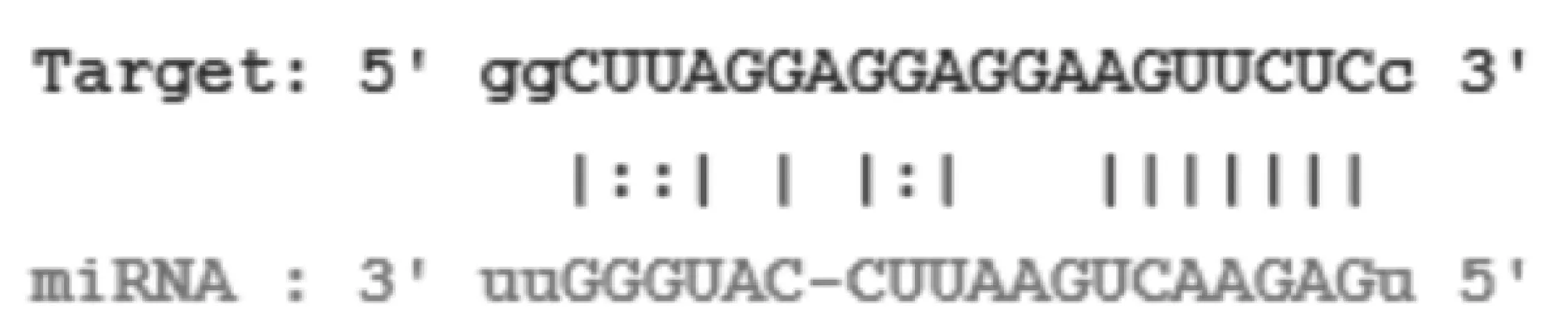
图1 生物信息学软件预测miR-146a是NEAT1下游的靶基因
2.3RT-qPCR检测miR-146a及TLRs信号通路相关分子mRNA的表达情况 以MOI=10.0的结核分枝杆菌感染RLE-6TN细胞,结果显示,与0 h时相比,6、12、24 h 时RLE-6TN细胞中的miR-146a表达水平显著降低(P<0.05),MyD88 mRNA表达水平显著升高(P<0.05),呈时间依赖性;TLR4 mRNA表达水平在6 h、12 h时显著升高(P<0.05),24 h时显著降低(P<0.05),NF-κB mRNA水平变化的差异无统计学意义(P>0.05),见表3。
表3 感染不同时间点miR-146a及TLRs信号通路相关分子的mRNA表达情况
2.4TLRs信号通路相关分子的蛋白水平变化 与0 h时相比,6 h时RLE-6TN细胞中TLR4的蛋白表达水平显著升高(P<0.05),12 h时显著降低(P<0.05),24 h时显著升高(P<0.05);6、12、24 h MyD88、TRAF6、p-NF-κB/NF-κB蛋白水平均显著升高(P<0.05),呈时间依赖性,见图2,表4。

图2 RLE-6TN细胞感染结核杆菌不同时间TLRs信号通路相关分子蛋白水平的变化情况
2.5RLE-6TN细胞炎症因子mRNA表达情况 与0 h时相比,6、12、24 h时RLE-6TN细胞中炎症因子IL-6、IL-8、TNF-α mRNA表达水平均显著升高(P<0.05),呈时间依赖性,见表5。
表4 感染不同时间点TLRs信号通路相关蛋白表达情况
表5 感染不同时间点RLE-6TN细胞中炎症因子IL-6、IL-8和TNF-α mRNA水平的比较
3 讨 论
肺结核是全球范围内最重要的传染疾病,肺结核的发病机制十分复杂,其中结核分枝杆菌是引起肺结核患者发病的首要因素〔7〕。结核分枝杆菌是一类典型的胞内寄生菌,其感染过程中涉及固有性免疫与适应性免疫,与抗原递呈、细胞因子释放等过程的机体免疫应答及炎症反应密切相关〔8〕。肺泡Ⅱ型细胞又称为颗粒肺泡细胞,约占肺泡总数的75%左右,具有增殖、分泌肺表面活性物质、维持肺泡内外液体平衡及免疫调节等过程〔9〕。近年来研究发现,肺泡Ⅱ型细胞在多种生理及病理条件下,能够分泌多种细胞因子、炎症因子,参与炎症反应过程,与肺结核的发病过程密切相关〔10〕。
LncRNA结构与mRNA相似,在真核生物体内表达广泛,能够参与许多生物学过程及细胞活动〔11〕。谭杨等〔12〕研究表明,巨噬细胞在结核分枝杆菌感染后,筛选出多种差异表达的LncRNAs,但未对这些LncRNA在结核病中的具体生物学机制进一步研究。NEAT1是LncRNA中的一种,能够参与机体内的免疫调节〔13〕。Morchikh等〔14〕研究表明,NEAT1与HEXIM1能形成多亚基复合物,参与调节DNA介导的先天免疫反应。本研究结果提示NEAT1表达上调可能参与结核分枝杆菌对肺泡Ⅱ型细胞感染过程。但具体生物学机制尚不清楚。
本研究结果表明miR-146a是NEAT1下游的直接靶标。miR-146a是最早证实与炎症相关的miRNAs之一〔15〕。以上结果提示结核分枝杆菌感染肺泡Ⅱ型细胞的生物学调节机制可能与NEAT1对miR-146a的调控有关。孙波等〔16〕研究表明,胃黏膜组织中miR-146a可能通过TLR4/NF-κB信号通路介导炎性反应。TLRs是天然防御系统的一个重要组成部分,其介导的炎症信号通路在心肌损伤及心肌炎等一系列炎症反应中具有关键作用〔17〕。马渝等〔18〕研究表明,miR-146a在脓毒症患者肺组织内表达上调,可抑制TNFα释放,减轻炎症反应。杨汀等〔19〕研究表明,miR-146a的作用靶点为TRAF6和IRAK-1的3′-UTR区,其能够下调IRAK1和TRAF6水平。本研究结果提示TLR4可能在早期仅具有细胞识别作用,但整体上结核分枝杆菌感染RLE-6TN细胞能够促进TLR4的表达。结核分枝杆菌感染细胞后,RLE-6TN细胞中NF-κBmRNA及蛋白表达变化无统计学意义,而其活化形式p-NF-κB蛋白水平显著升高,呈时间依赖性,与TRAF6、MyD88 mRNA及其蛋白变化趋势一致,提示结核菌感染细胞后可能会激活TLRs-NF-κB信号通路。本研究与郭茂等〔20〕研究结果一致。郭茂等〔20〕研究表明,右美托咪定处理脓毒症小鼠外周血单核细胞后,细胞中miR-146a呈高表达,可明显抑制小鼠炎症因子TNF-α、IL-6水平,其机制可能与抑制TLR4-NF-κB信号通路中重要接头蛋白TRAF6、IRAK-1表达有关。提示结核分枝杆菌感染RLE-6TN细胞过程中可能通过激活TLR4-NF-κB信号通,促进促炎因子IL-6、IL-8、TNF-α表达,导致感染发生。
综上,LncRNA NEAT1在结核分枝杆菌感染肺泡Ⅱ型细胞中表达上调,NEAT1可能通过靶向miR-146a,参与调控TLRs信号通路活化及诱导炎症因子的表达。然而LncRNA NEAT1在结核分枝杆菌感染肺泡Ⅱ型细胞中的具体生物学调节机制还不清楚,仍需进一步进行研究。
