A New Species of the Asian Leaf Litter Toad Genus Leptobrachella (Amphibia,Anura,Megophryidae) from Chongqing City,Southwest China
2022-06-25TaoLUOWeifengWANGDongPENGBoLEIHuaiqingDENGShengnanJIHeqingHUANGandJiangZHOU
Tao LUO ,Weifeng WANG,Dong PENGBo LEI,Huaiqing DENGShengnan JI,Heqing HUANG* and Jiang ZHOU
1 School of life Sciences,Guizhou Normal University,Guiyang 550001,Guizhou,China
2 School of Karst Science,Guizhou Normal University,Guiyang 550001,Guizhou,China
3 Biodiversity Research Center,Chinese Research Academy of Environmental Sciences,Beijing 100012,China
4 Chongqing Academy of Environmental Science,Chongqing 401147,China
Abstract Herein we describe a new species,Leptobrachella yunyangensis sp.nov.,from Yunyang County,Chongqing City,China,based on a combination of molecular and morphological data.Phylogenetic analysis based on the mitochondrial 16S rRNA gene and six nuclear genes indicated that this new species represented an independent evolutionary lineage.The uncorrected genetic distance between the new species and its closest congener species,L.oshanensis,was 5.4 % for 16S rRNA.The new species can be distinguished from its congeners by a combination of the following characters:(1) moderate body size (SVL 28.3-30.6 mm in males);(2) rough dorsal skin,with sparse large granules and tubercles and short longitudinal ridges on the shoulder;(3) distinctly discernible tympanum with a diameter smaller than that of the eye (TMP/EYE ratio:0.51) and a distinct black supratympanic line present;(4)an internasal distance almost equal to interorbital distance(IND/IOD ratio:1.05);(5) flanks with several dark spots arranged longitudinally in two rows;(6) distinctly visible supra-axillary,femoral,pectoral,and ventrolateral glands;(7) bicolored iris,with the upper 1/3 of the iris being copper orange and the lower 2/3 a light silvery grey;(8) relative finger lengths of I < II=IV < III and relative toe lengths of I < II < V < III < IV;(9) absence of webbing and lateral fringes on fingers,and toes with rudimentary webbing and narrow lateral fringes;(10) heels overlapping when the thighs are positioned at right angles to the body;(11) tibiotarsal articulation reaching to the anterior corner of the eye when the leg is stretched forward;(12) ventral surfaces of the throat,chest,and belly greyish white with purple-brown speckling;(13) lacking distinct blackish dorsolateral markings;and (14) transverse dark brown bars on the surfaces of limbs and digits.
Keywords mitochondrial DNA,morphology,new species,nuclear gene,taxonomy
1.Introduction
The genusLeptolalax(Dubois,1983) in the family Megophryidae(Bonaparte,1850),is regarded as closely associated with the genusLeptobrachella(Smith,1925) and has been assigned as a synonym of the genusLeptobrachellabased on a large-scale molecular analysis(Chenet al.,2018).Currently,the genusLeptobrachellacontains 91 species (Frost,2021;Chenet al.,2021a) that are widely distributed from southern China to northeastern India and Myanmar,extending to mainland Indochina,peninsular Malaysia,and the island of Borneo (Feiet al.,2012;Frost,2021).Leptobrachellahas a relatively high diversity of cryptic species,with nearly 59 species described since 2010 (see theLeptobrachellaspecies list in Frost 2021;Chenet al.,2021a).Currently,33 species of this genus are known from China (AmphibiaChina,2021).These areL.alpina(Fei,Ye and Li,1990) andL.bourreti(Dubois,1983) from Yunnan and Guangxi;L.asperaWang,Lyu,Qi and Wang,2020,L.eos(Ohler,Wollenberg,Grosjean,Hendrix,Vences,Ziegler and Dubois,2011),L.feiiChen,Yuan and Che,2020,L.flaviglandulosaChen,Wang and Che,2020,L.nyx(Ohler,Wollenberg,Grosjean,Hendrix,Vences,Ziegler and Dubois,2011),L.niveimontisChen,Poyarkov,Yuan and Che,2020,L.purpurus(Yang,Zeng and Wang,2018),L.pelodytoides(Boulenger,1893),L.tengchongensis(Yang,Wang,Chen and Rao,2016),andL.yingjiangensis(Yang,Zeng and Wang,2018) from Yunnan;L.laui(Sung,Yang and Wang,2014)andL.yunkaiensis(Wang,Li,Lyu and Wang,2018) from southern Guangdong,including Hong Kong;L.liui(Fei and Ye,1990) from Fujian,Jiangxi,Guangdong,Guangxi,Hunan,and Guizhou;L.oshanensis(Liu,1950) from Gansu,Sichuan,Chongqing,Guizhou,and Hubei;L.yeaeShi,Hou,Song,Jiang and Wang,2021 from Sichuan;L.bashaensisLyu,Dai,Wei,He,Yuan,Shi,Zhou,Ran,Kuang,Guo,Wei and Yuan,2020,L.purpuraventraWang,Li,Li,Chen and Wang,2019,L.bijieWang,Li,Li,Chen and Wang,2019,L.suiyangensisLuo,Xiao,Gao and Zhou,2020,L.chishuiensisLi,Liu,Wei and Wang,2020,L.dorsospinaWang,Lyu,Qi and Wang,2020,andL.jinshaensisCheng,Shi,Li,Liu,Li and Wang,2021 from Guizhou;L.ventripunctata(Fei,Ye and Li,1990) from Guizhou and Yunnan;L.mangshanensis(Hou,Zhang,Hu,Li,Shi,Chen,Mo and Wang,2018),andL.wulingensisQian,Xiao,Cao,Xiao and Yang,2020 from southern Hunan and Guihzou,andL.damingshanensisChen,Yu,Cheng,Meng,Wei,Zhou and Lu,2021,L.sungi(Lathrop,Murphy,Orlov and Ho,1998),L.maoershanensis(Yuan,Sun,Chen,Rowley and Che,2017),L.shangsiensisChen,Liao,Zhou and Mo,2019,L.shiwandashanensisChen,Peng,Pan,Liao,Liu and Huang,2021,andL.wuhuangmontisWang,Yang and Wang,2018 from Guangxi (Feiet al.,2012;Sunget al.,2014;Yanget al.,2016;Yuanet al.,2017;Yanget al.,2018;Wanget al.,2018;Houet al.,2018;Chenet al.,2019;Wanget al.,2019,2020;Chenet al.,2018,2020;Luoet al.,2020;Lyuet al.,2020;Liet al.,2020;Qianet al.,2020;Chenet al.,2021a,b;Chenget al.,2021;Liuet al.,2021;Shiet al.,2021).The continual discovery of new species suggests thatLeptobrachellaspecies diversity may be grossly underestimated,and thus extensive field surveys by researchers are required.
During biodiversity surveys in Yunyang County,Chongqing,China,during June 2021,we collected several specimens of an unknownLeptobrachellaspecies.The specimens were assigned to the genusLeptobrachellaon the basis of the following characters:(1) a comparatively small body size,with a snout-vent length less than 60.0 mm;(2) having an elevated inner metacarpal tubercle;(3) having macro-glands on the body (including supraaxillary,femoral,and ventrolateral glands);(4) lacking vomerine teeth;(5) having small tubercles on the eyelids;(6) having a whitish vertical bar on the anterior tip of the snout (Dubois,1983;Matsui,1997,2006;Lathropet al.,1998;Delormeet al.,2006;Daset al.,2010).Subsequent phylogenetic analysis of mitochondrial genes and nuclear gene fragments revealed that these specimens belonged to the genusLeptobrachellaand represented a distinct evolutionary lineage.Combining morphological data and molecular genetic differences,we describe the specimens as a new species.
2.Materials and Methods
2.1.SamplingA total of 12 new specimens were collected in this study.Six specimens were of the undescribed species from Qiyaoshan Nature Reserve and Lianhua Village,Renhe Town,Yunyang County,Chongqing,China;six wereL.alpinafrom Wulian Mountain,Jingdong County,Yunnan,China.All specimens were fixed in 10% buffered formalin and later transferred to 75% ethanol for preservation.The muscle samples used for molecular analysis were preserved in 95% alcohol and stored at -20°C.All newly collected specimens were kept at Guizhou Normal University (GZNU),Guiyang City,Guizhou Province,China.
2.2.DNA extraction,PCR,and sequencingGenomic DNA was extracted from muscular tissue using a DNA extraction kit from Tiangen Biotech Co.,Ltd.(Beijing,China).Seven tissue samples used for molecular analysis were amplified and sequenced for one mitochondrial gene and six nuclear genes:the partial 16S ribosomal RNA gene (16S rRNA),brain-derived neurotrophic factor(BDNF),sodium/calcium exchanger 1 (NCX),neurotrophin 3(NTF3),recombination activating gene 1 (RAG1),rhodopsin(RHOD),and solute carrier family 8 member 3 (SLC8A3)(Primer sequences are in Table S1).PCR amplifications were performed in a 20 μl reaction volume with the following cycling conditions:an initial denaturing step at 95°C for 5 min,35 cycles of denaturing at 95°C for 1 min,annealing at 50-57°C for 1 min,and extending at 72°C for 1 min,followed by a final extending step at 72°C for 10 min.PCR products were purified with spin columns.The purified products were sequenced with both forward and reverse primers using a BigDye Terminator Cycle Sequencing Kit according to the guidelines of the manufacturer.The products were sequenced on an ABI Prism 3730 automated DNA sequencer at Chengdu TSING KE Biological Technology Co.Ltd (Chengdu,China).All newly obtained sequences have been submitted to GenBank (Table 1).
2.3.Phylogenetic analysesFor the molecular analysis,we used 276 sequences from 79 species of the genusLeptobrachellafor phylogenetic analysis,including 52 sequences newly amplified and sequenced from nine muscle tissue samples and 224 sequences downloaded from GenBank.In addition,we followed the methods of Chenet al.(2018) and selectedLeptobrachium huashen(Fei and Ye,2005),Leptobrachiumchapaense(Bourret,1937),andMegophrys major(Boulenger,1908) as outgroups and downloaded 21 sequences from GenBank (Table 1).

For the phylogenetic analysis,we constructed two sequence matrices for reconstructing the phylogenetic tree,i.e.,mitochondrial 16S rRNA and the combined sequences of six nuclear genes.All of the sequences were assembled and aligned using the MUSCLE(Edgar,2004) module in MEGA 7.0 (Kumaret al.,2016) with default settings.Alignments were checked by eye and revised manually if necessary.Trimming with the gaps partially deleted was performed in GBLOCKS 0.91b (Castresana,2000).Phylogenetic analysis of the two sequence matrices was performed using maximum likelihood (ML) and Bayesian inference (BI).The bestfit nucleotide substitution models for the sequence supermatrices were selected in PartitionFinder 2.1.1 (Lanfearet al.,2016) using the Bayesian information criterion (BIC).The results indicated GTR+I+G as the best-fitting nucleotide substitution model for the 16S rRNA sequence matrix,and K80+I+G (from SLC8A3 and RAG1),HKY+I+G (from NTF3,RHOD,and NCX),and K80+I (from BDNF) as the best-fitting nucleotide substitution models for the combined nuclear gene sequence matrix.Phylogenetic analysis using maximum likelihood (ML) and Bayesian inference (BI)methods was implemented in IQ-tree 2.0.4 (Nguyenet al.,2015)and MrBayes 3.2.1 (Ronquistet al.,2012),respectively.The ML analysis was run using the best-fit model for each partition with 2000 ultrafast bootstrap (UFB) replicates (Minhet al.,2013),and was performed until a correlation coefficient of at least 0.99 was reached (Hoanget al.,2018).For BI analyses,two independent runs were performed with four Markov chains (three heated chains and a single cold chain) based on the best-fit partitioning schemes and corresponding substitution models selected.Starting from a random tree,each run consisted of a total of 1 × 107generations,sampled every 1000 generations.Convergence was assessed by the average standard deviation of split frequencies (ASDSF) being less than 0.01,and the effective sample sizes (ESS) being greater than 200 in Tracer 1.7.1 (Rambautet al.,2014).Nodes in the trees were considered well supported when Bayesian posterior probabilities(BPP) were ≥ 0.95 and ML ultrafast bootstrap (UFB) values were≥ 95%.Finally,genetic distance between species via the uncorrectedP-distance model for the 16S rRNA gene was estimated using MEGA 7.0.
2.4.Morphological and morphometric analysesMorphometric data were taken from six of the most well-preserved adult specimens (Table 2).Measurements were recorded to the nearest 0.1 mm using digital calipers following the methods of Feiet al.(2009) and Rowleyet al.(2013).These measurements were as follows:SVL=snout-vent length (from tip of snout to vent);HDL=head length (from tip of snout to rear of jaws);HDW=head width (head width at the commissure of the jaws);SNT=snout length (from tip of snout to the anterior corner of the eye);EYE=eye diameter (diameter of the exposed portion ofthe eyeballs);IOD=interorbital distance (minimum distance between upper eyelids);IND=internasal distance (distance between nares);UEW=upper eyelid width (measured as the greatest width of the upper eyelid);NEL=nostril-eyelid length(distance from nostril to eyelid);TMP=tympanum diameter(horizontal diameter of the tympanum);TEY=tympanumeye distance (distance from anterior edge of the tympanum to posterior corner of the eye);HND=hand length (distance from distal end of radioulna to tip of phalanx of finger III);LAHL=length of the lower arm and hand (distance from tip of the third finger to elbow);LW=lower arm width (maximum width of the lower arm);HLL=hindlimb length (distance from tip of fourth toe to vent);THL=thigh length (distance from vent to knee);TIB=tibia length (distance from knee to heel);TW=maximal tibia width;FOT=foot length (from proximal edge of the inner metatarsal tubercle to the tip of the fourth toe);TFL=length of foot and tarsus (distance from the tibiotarsal articulation to the distal end of toe IV);IPTL=inner palmar tubercle length (measured as maximal distance from proximal to distal ends of the inner palmar tubercle);OPTL=outer metacarpal tubercle length (measured as maximal diameter of the outer metacarpal tubercle);and IMTL=inner metatarsal tubercle length (taken as maximal length of the inner metatarsal tubercle).
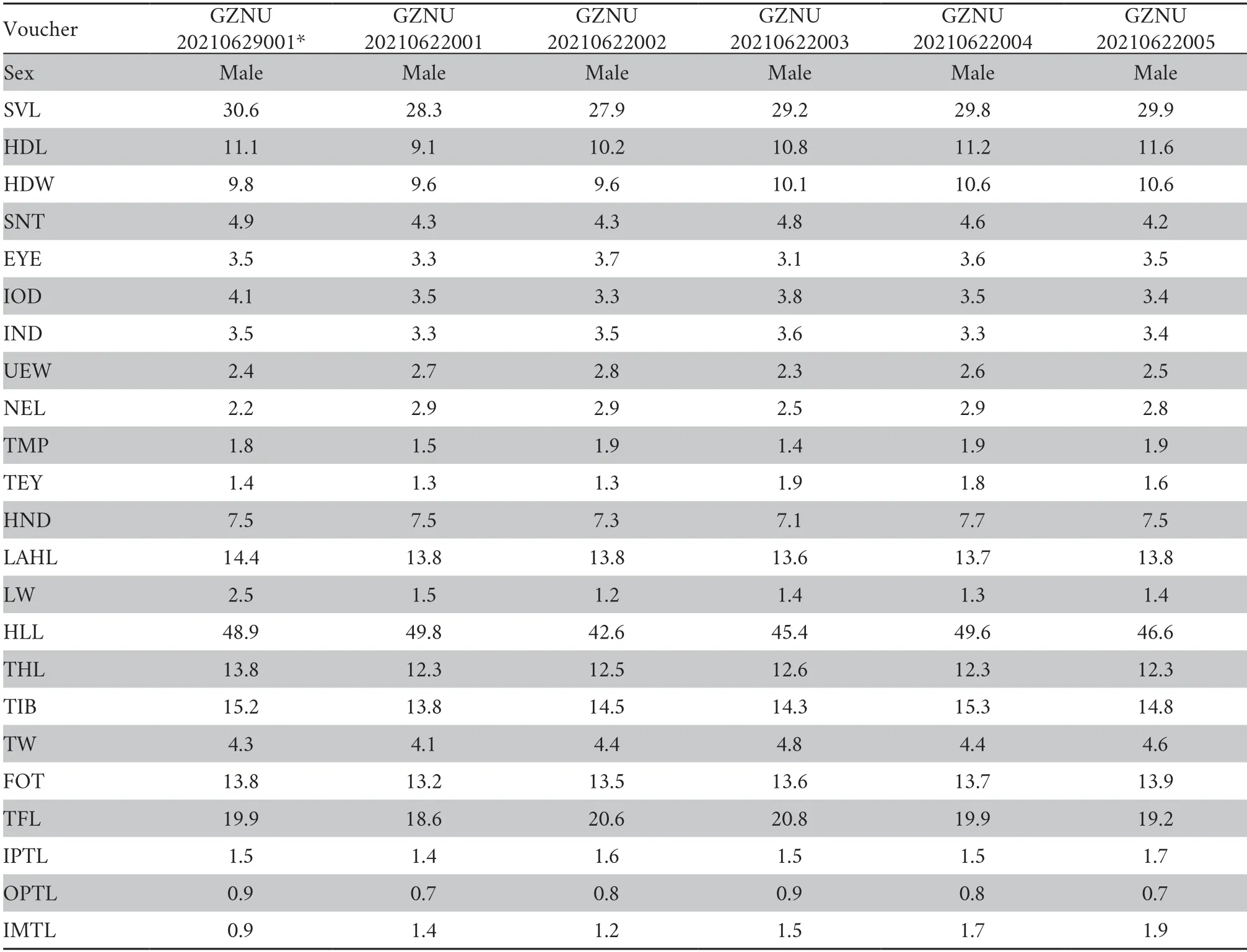
Table 2 Measurements of the adult specimens of Leptobrachella yunyangensis sp.nov.All units in mm.See abbreviations for the morphological characters in the Materials and Methods section.*for the holotype.
To reduce the impact of allometry,the correct value from the ratio of each character to SVL was calculated,and then all data were log-transformed (log10) for the subsequent morphometric analyses.Principal component analyses (PCAs) of size-corrected measurements and simple bivariate scatterplots were used to explore and characterize the morphometric differences between the undescribed species andL.oshanensis.Mann-WhitneyUtests were conducted to determine the significance of differences in morphometric characters between the undescribed species andL.oshanensisin males.All statistical analyses were performed using SPSS 21.0 (SPSS,Inc.,Chicago,IL,USA),and differences were considered statistically significant atP< 0.05.
Sex was determined by the presence of internal vocal sac openings,and the presence of eggs in the abdomen through external inspection.Comparative morphological data for species of the genusLeptobrachellawere obtained from the literature(Table 3).Owing to the high likelihood of undiagnosed diversity within the genus (Rowleyet al.,2016;Yanget al.,2016),where available we relied on examination of topotypic material and/or original species descriptions.We also examined the type and/or topotype materials forL.alpina,L.suiyangensis,andL.wulingensis(Appendix).
3.Results
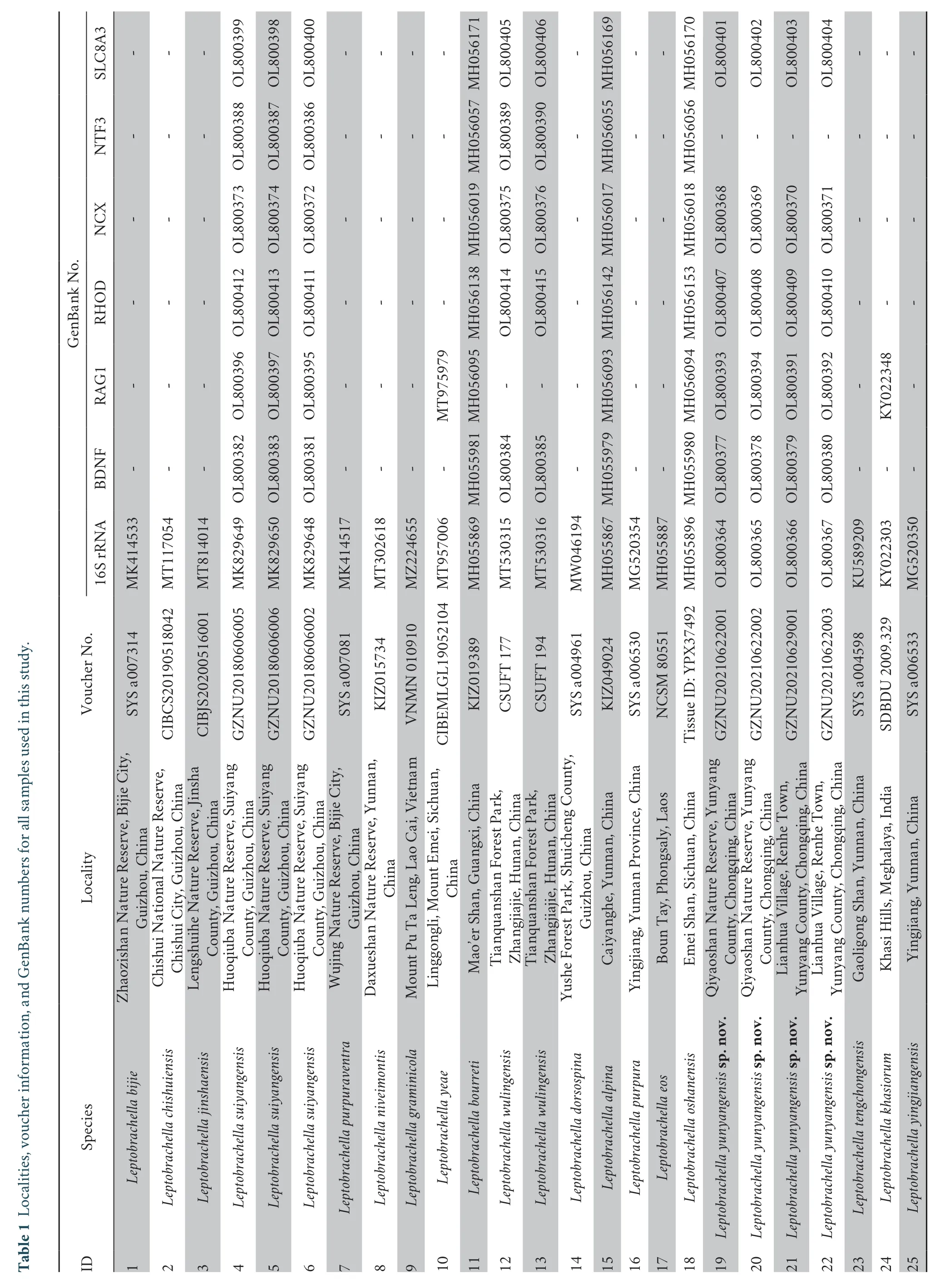
3.1.Phylogenetic analyses and genetic divergencePhylogenetic trees from maximum likelihood (ML) and Bayesian inference (BI) were reconstructed based on DNA sequences of the mitochondrial 16S rRNA gene with a length of 532 base pairs (bp).In the 16S rRNA gene trees,twoLeptobrachellapopulations from Yunyang County,Chongqing,China,clustered into a separate evolutionary lineage and were highly supported (1.00 in BI and 99 % in ML;Figure 1).
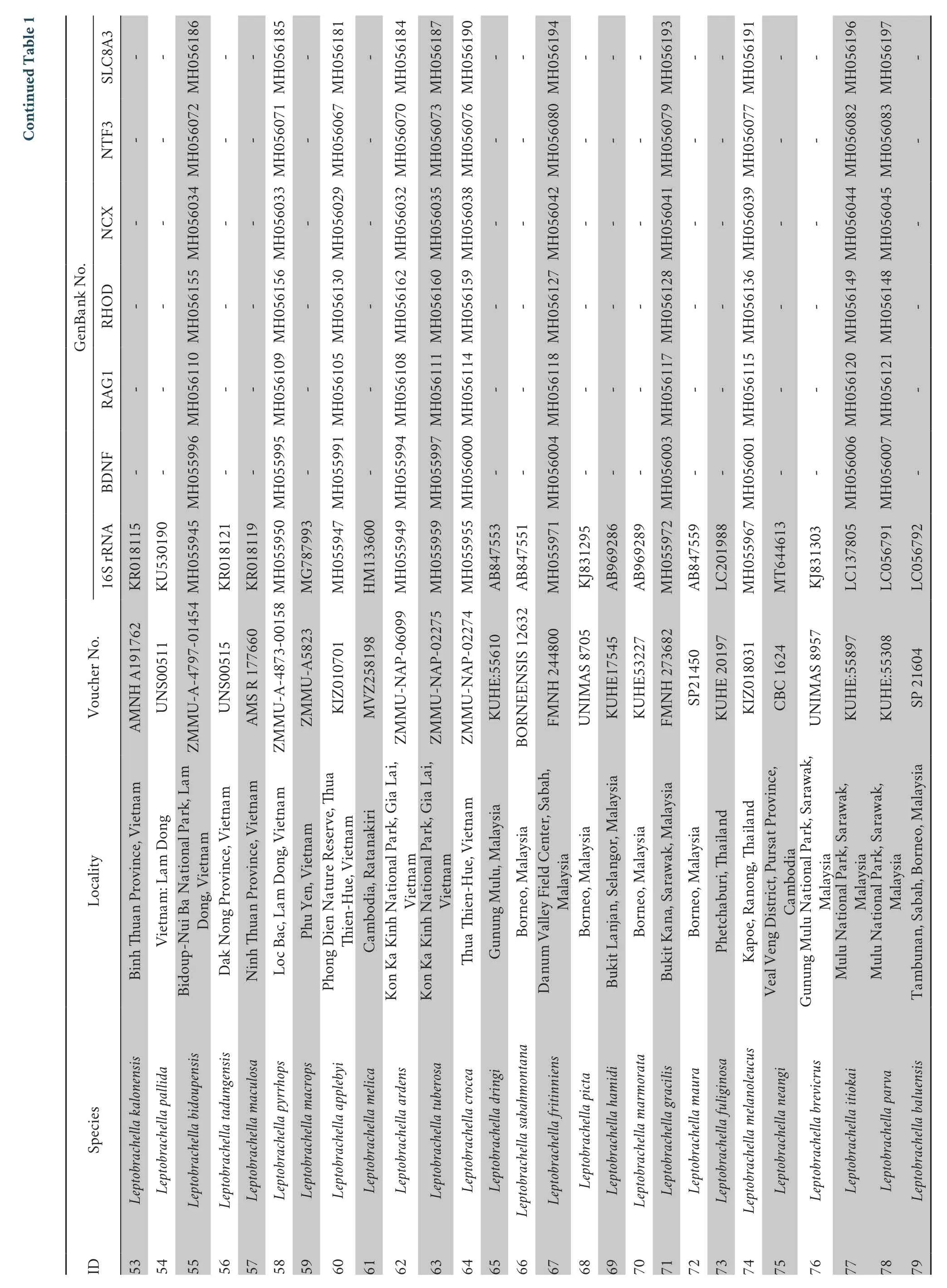
The analysis of the combined six nuclear loci (3594 bp in total length) (BDNF:426 bp;RHOD:321 bp;NCX:426 bp;SLC8A3:540 bp;RAG1:1035 bp;NTF3:567 bp) well resolved the phylogenetic relationships between the major clades of the genusLeptobrachella(Figure 2).These topologies differed from the matrilineal tree due to the lack of sufficient species.However,the two populations of the new species clustered into a highly independent evolutionary lineage that formed a sister clade withL.oshanensis(1.00 in BI and 100 % in ML;Figure 2).
The smallest pairwise genetic divergence between the population from Yunyang County,Chongqing,China,and 78 species of the genusLeptobrachellawas 4.8% (vs.L.bourreti)to 20.4% (vs.L.kecil).These levels were similar or higher to the divergence among recognized congeners;for example,1.9%betweenL.bijieandL.chishuiensisand 2.9 % betweenL.asperaandL.feii(Table S2).In conclusion,the genetic divergence,matrilineal tree,and nuclear gene tree were all highly supportive of the twoLeptobrachellapopulations in Yunyang County,Chongqing,China,as an independent evolutionary lineage.
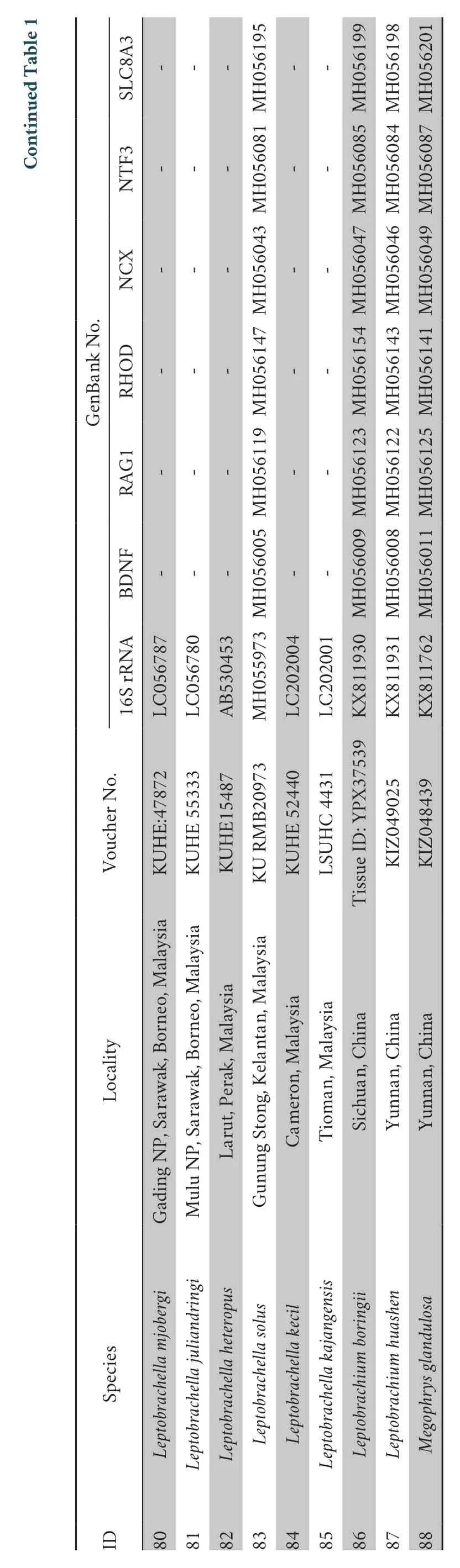
3.2.Morphological analysesThe results of the Mann-WhitneyUtests indicated that males of the new species differed significantly fromL.oshanensisbased on several morphometric characters,including SNT,IOD,IND,NEL,LW,TIB,TW,and FOT (allP-values < 0.05;Table 4).For the male group,PCA extracted three principal component axes with eigenvalues greater than two,and the percentages of explained variance of the first three principal components were 31.18%,15.91%,and 12.25%,respectively,with a cumulative percentage of 60.33%(Table S3).There were 13 major morphological characteristics in the first three principal components that were mainly distributed in the head and hind limbs (Table S3).The first two principal components explained 48.08% of the total variation in male morphology.In the PCA plot composed of PC 1 and PC 2,the first principal component axis could separate the new species fromL.oshanensis(Figure 3) and was based on SNT,IOD,IND,TEY,TIB,and TW.The second component axis was based on UEW,HND,and LAHL.Based on the genetic and morphological differences,we describe the specimens here as a new species.
Table 3 References for morphological characters for congeners of the genus Leptobrachella.

Continued Table 3
3.3.Taxonomy accounts
Leptobrachella yunyangensis sp.nov.Luo,Deng and ZhouTable 2;Figures 4-6
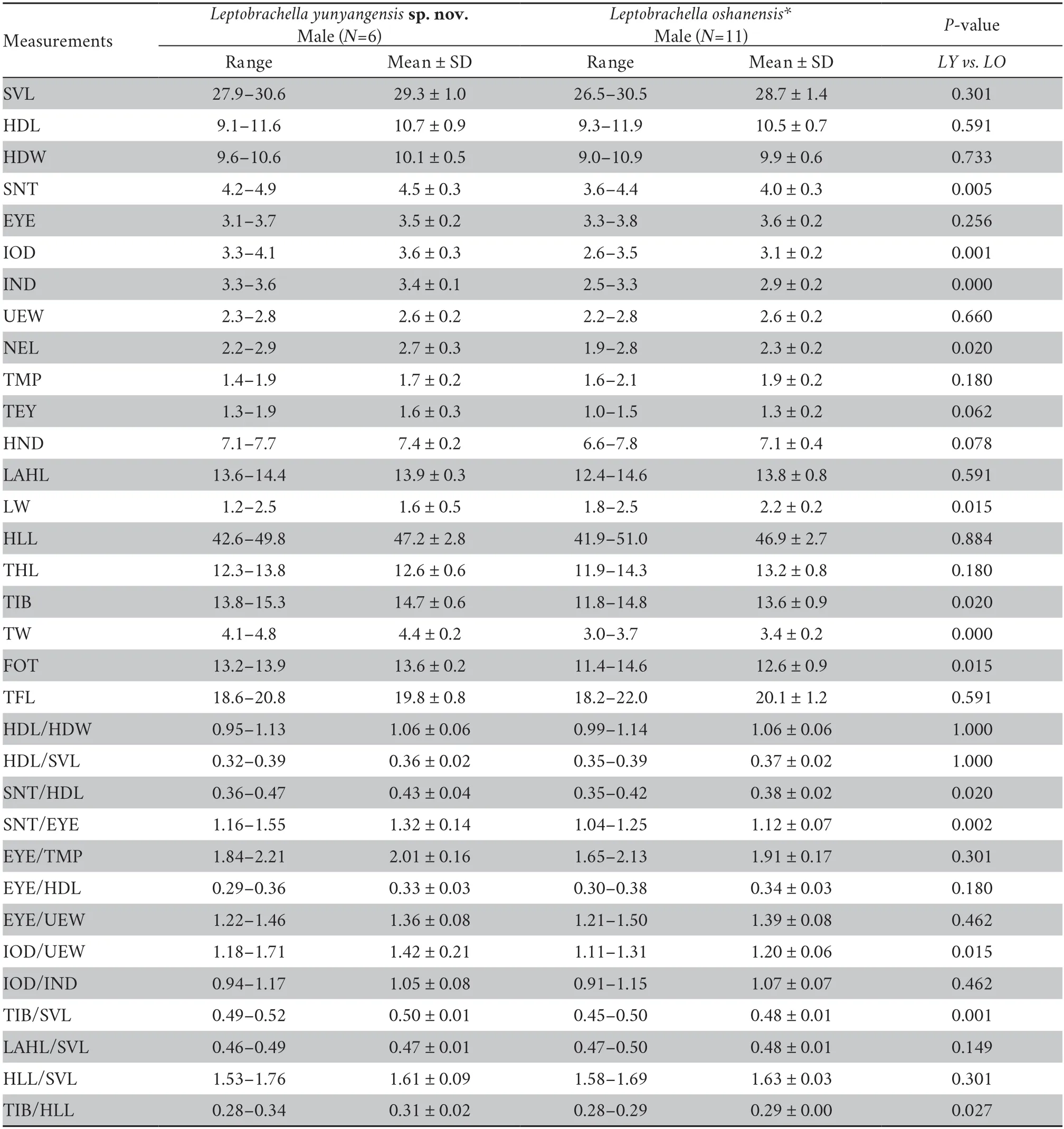
Table 4 Morphological comparison of Leptobrachella yunyangensis sp.nov.(LY) and L.oshanensis (LO).All units in mm.The P-values were from Mann-Whitney U tests.The significance level was set at P < 0.05.Morphometric characters are explained in the Materials and Methods section.
HolotypeGZNU20210629001,adult male collected by Tao Luo on 29 June 2021 from the Lianhua Village,Renhe Town,Yunyang County,Chongqing,China (108.614642°N,31.01610562°E,908 m.a.s.l.).
ParatypesGZNU20210622001,adult male collected from Qiyaoshan Nature Reserve,Yunyang County,Chongqing,China,by Tao Luo on June 22,2021;GZNU20210622002-005,four adult males collected from Qiyaoshan Nature Reserve,Yunyang County,Chongqing,China (108.78942255°N,30.6896741°E,839 m.a.s.l.),by Tao Luo and Dong Peng.
EtymologyThe specific epithetyunyangensisis in reference to the type locality,Lianhua Village,Renhe Town,Yunyang County,Chongqing,China.For the common name,we suggest“Guishi Leaf Litter Toad”,and for the Chinese name “Gui Shi Zhang Tu Chan (贵师掌突蟾)”.The year 2021 marks the 80th anniversary of the founding of Guizhou Normal University,which has made outstanding contributions to the training of teachers in Guizhou Province and the promotion of zoological research in China,and I would like to express my heartfelt wishes to Guishi,which comes from the abbreviation of Guizhou Normal University.
DiagnosisLeptobrachella yunyangensissp.nov.can be distinguished from its congeners by a combination of the following characters:(1) moderate body size (SVL 28.3-30.6 mm in males);(2) rough dorsal skin,with sparse large granules and tubercles and short longitudinal ridges on the shoulder;(3) distinctly discernible tympanum with a diameter smaller than that of the eye (TMP/EYE ratio:0.51) and a distinct black supratympanic line present;(4) an internasal distance almost equal to interorbital distance (IND/IOD ratio:1.05);(5)flanks with several dark spots arranged longitudinally in two rows;(6) distinctly visible supra-axillary,femoral,pectoral,and ventrolateral glands;(7) bicolored iris,with the upper 1/3 of the iris being copper orange and the lower 2/3 a light silvery grey;(8) relative finger lengths of I < II=IV < III and relative toe lengths of I < II < V < III < IV;(9) absence of webbing and lateral fringes on fingers,and toes with rudimentary webbing and narrow lateral fringes;(10) heels overlapping when thethighs are positioned at right angles to the body;(11) tibiotarsal articulation reaching to the anterior corner of the eye when the leg is stretched forward;(12) ventral surfaces of the throat,chest,and belly greyish white with purple-brown speckling;(13) lacking distinct blackish dorsolateral markings;and (14)transverse dark brown bars on the surfaces of limbs and digits.Description of the holotypeGZNU20210629001 (Figures 4 and 5),adult male.Body size moderate,SVL 30.6 mm.

Figure 1 Bayesian inference (BI) tree based on mitochondrial 16S rRNA.In this phylogenetic tree,Bayesian posterior probabilities (BPP)from BI analyses/ultrafast bootstrap supports (UFB) from ML analyses are listed beside the nodes.The symbol “-” represents a value below 0.60/60.The scale bar represents 0.08 nucleotide substitutions per site.The numbers at the tips of branches correspond to the ID numbers in Table 1.
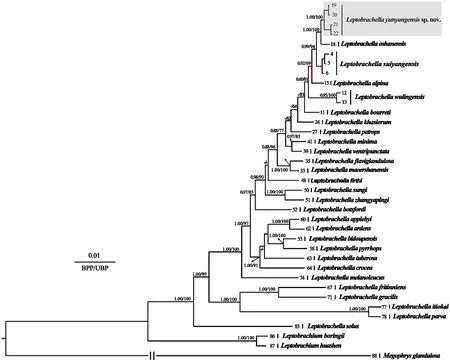
Figure 2 Bayesian inference (BI) tree based on combined nuclear data (six genes).BPP from BI analyses/UFB from ML analyses are listed beside the nodes.The symbol “-” represents values below 0.60/60.The scale bar represents 0.01 nucleotide substitutions per site.The numbers at the tips of branches correspond to the ID numbers in Table 1.

Figure 3 Plots of the first principal component (PC1) versus the second (PC2) for Leptobrachella yunyangensis sp.nov.and L.oshanensis from a principal component analysis.
Headlength slightly wider than head width (HDL/HDW ratio:1.13);snout short,rounded,and projecting beyond the lower jaw in dorsal view,longer than eye diameter (SNT/EYE ratio:1.40);nostril rounded,distinct,and closer to the eye than tip of the snout (NEL/ SNT ratio:0.45);canthus rostralis gently rounded;loreal region slightly concave;interorbital space flat and interorbital distance slightly longer than internarial distance(IOD/IND ratio:1.17);pineal ocellus absent;pupil vertical;eyes large,with eye diameter slightly equal to internarial distance(IND/EYE ratio:1.00);tympanum distinct,rounded,and slightly concave,with a diameter smaller than that of the eye and larger than the tympanum-eye distance (TMP/EYE ratio:0.51,TEY/TMP ratio:0.78);upper margin of tympanum in contact with supratympanic ridge;distinct light-black supratympanic line present;vomerine teeth absent;vocal sac openings slit-like,paired,located posterolaterally on floor of the mouth in close proximity to the margins of the mandible;tongue cordiform,deeply notched behind;supratympanic ridge distinct,extending from posterior corner of the eye to the supra-axillary gland.
Forelimbsslender and comparatively short,length of lower arm and hand 47.06% of snout-vent length;tips of fingers rounded,slightly swollen;relative finger lengths I < II=IV < III;nuptial pad absent;subarticular tubercles absent;inner palmar tubercle large,rounded,and separated from the smaller,round outer palmar tubercle;inner metacarpal tubercles longer than outer metacarpal tubercles (IPTL/OPTL ratio:1.67);absence of webbing and lateral fringes on fingers.
Hindlimbsslender,tibia slightly longer than thigh length(TIB/THL ratio:1.10) and 45.10% of snout-vent length;heels overlapping when thighs are positioned at right angles to the body;relative toe length I < II < V < III < IV;tips of toes round and slightly swollen;subarticular tubercles absent;toe webbing rudimentary;narrow lateral fringes present on all toes;inner metatarsal tubercle present and large,oval,outer metatarsal tubercle absent;tibiotarsal articulation reaching to anterior corner of the eye when the leg is stretched forward.
Dorsal skin rough,with sparse large granules and tubercles;short longitudinal ridges on the shoulder;conical spines on the flanks;ventral skin smooth;pectoral gland and femoral gland milky-white,distinctly visible,and oval;long longitudinal skin ridges on dorsal surface of the limbs;dense tiny granules present on ventral surface of thigh and tibia;femoral gland situated on posteroventral surface of the thigh,closer to knee than to vent,oval;supra-axillary glands long,oval,and raised;ventrolateral glands distinctly visible and raised,forming an incomplete line.
Coloration of holotype in life (Figure 4)Dorsum greyish brown,with distinct darker brown markings on sides and scattered with irregular light greyish brown pigmentation and yellowish brown spots;a very indistinct,light brown inverted triangular pattern between the anterior corners of the eyes,without a dark brown W-shaped marking between the axillae;tympanum bicolored,with the upper 2/3 of the tympanum dark brown and the lower 1/3 light orange;light brown between the posterior corner of the eyes and the tympanum;sparse,small,light brown granules and small dark brown patches present on the dorsum of the body and limbs;transverse dark brown bars on dorsal surface of limbs and digits;distinct dark brown spots on the flanks from groin to axilla arranged longitudinally in two rows;elbow and upper arms with distinct light orange coloration;iris bicolored,with the upper 1/3 of the iris copper orange and the lower 2/3 light silver gray.
Ventral surface of the throat,chest,and belly greyish white with purple-brown speckling;supra-axillary gland,femoral,pectoral,and ventrolateral glands white;ventral surface of limbs and lower lip dark grey-purple,scattered with small greyish white spots,creamy white tubercles,and small granules.
Coloration of holotype in preservation (Figure 5)Dorsum of body and limbs fade to dark brown except for upper arm;upper arm surface copper brown;skin on dorsal becoming more smooth;transverse bars on limbs becoming more distinct,and dorsolateral markings,longitudinal skin ridges and spots on back becoming indistinct.Ventral surface of limbs and surface of throat light brown,surface of abdomen greyish white,dark patches on chest,abdomen,and flanks becoming more distinct.Supra-axillary,femoral,pectoral,and ventrolateral glands greyish white.
VariationMeasurements of the type series are shown in Table 2.All the male paratypes match the overall characters of the holotype,except that the ventral surface of the throat,chest,and belly is greyish brown in the holotype GZNU20210629001 (vs.light purple grey in the paratype GZNU20210622001) (Figure 6);tibiotarsal articulation with the same color as the thigh and tibia surfaces (vs.brown in the paratype GZNU20210622001);ventral surface of limbs dark purple grey (vs.violet blue in the paratype GZNU20210622001).

Figure 4 Morphological features of the live adult male holotype GZNU20210629001 of Leptobrachella yunyangensis sp.nov.A:dorsolateral view;B:dorsal view;C:ventral view;D:ventral view of the hand;E:ventral view of the foot.Photos A to E were taken at around 9:00 am.

Figure 5 Morphological features of the preserved adult male holotype GZNU20210629001 of Leptobrachella yunyangensis sp.nov.A:dorsal view;B:ventral view;C:ventral view of hand;D:ventral view of foot.

Figure 6 Morphological features in life.Leptobrachella yunyangensis sp.nov.,paratype GZNU2021062200.A:dorsal view;B:dorsolateral view;C:ventral view;D:ventral view of the hand;E:ventral view of the foot.Photos A to E were taken at around 9:00 am.
Sexual dimorphismAdult males with a comparatively large single subgular vocal sac,and absent nuptial pads and spines.
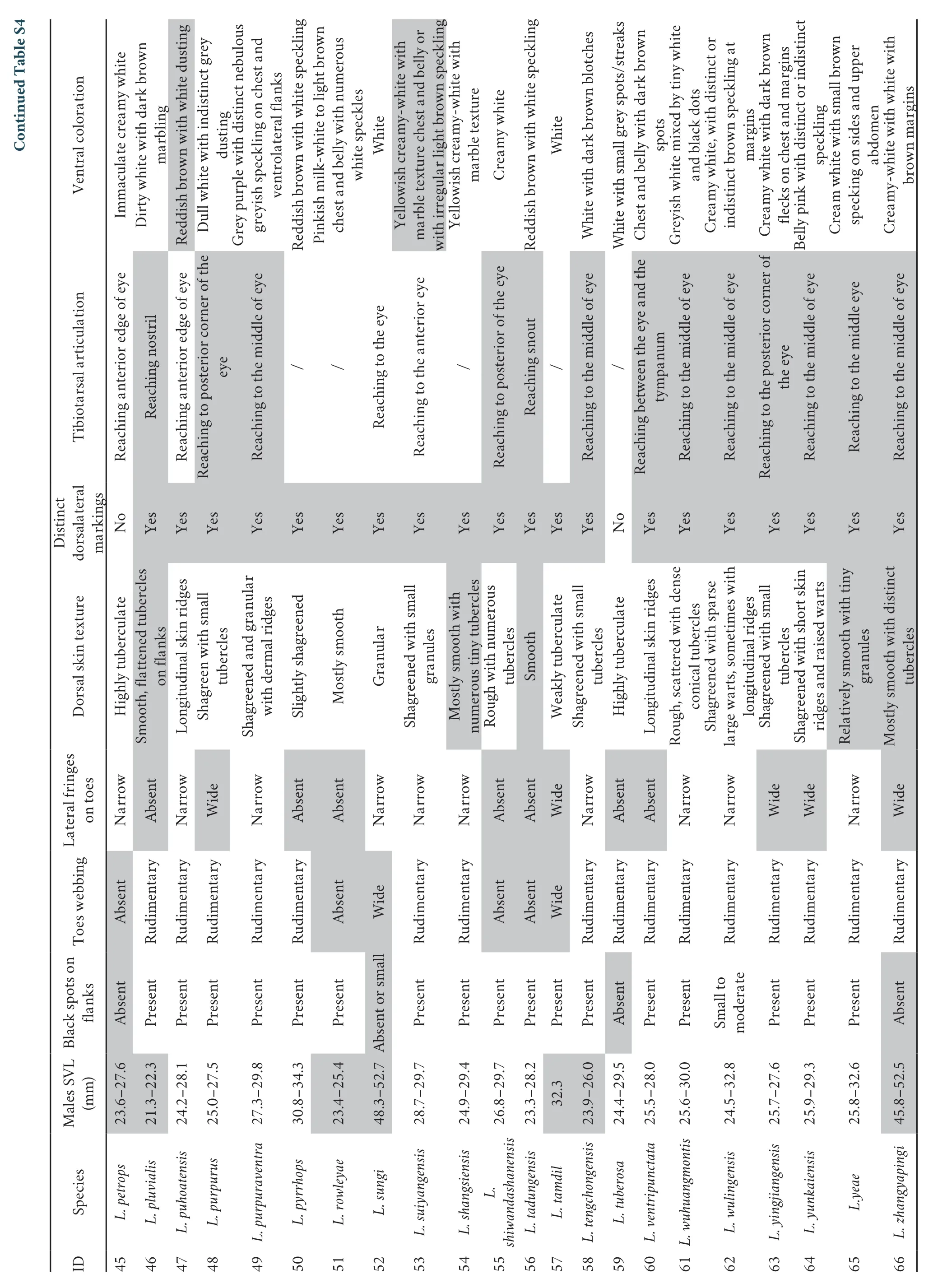
ComparisonComparative information on the new species with 63 recognized species of the genusLeptobrachellanorth of the Kra Isthmus is given in Table S4.
From the 26 known congeners of the genusLeptobrachellaoccurring south of the Kra Isthmus,by the presence of supraaxillary and ventrolateral glands,Leptobrachella yunyangensissp.nov.can easily be distinguished fromL.arayai,L.dringi,L.fritinniens,L.gracilis,L.hamidi,L.heteropus,L.kajangensis,L.kecil,L.marmorata,L.melanoleuca,L.maura,L.picta,L.platycephala,L.sabahmontana,andL.sola(vs.lacking supra-axillary and ventrolateral glands in the latter) by having a significantly larger body size (SVL 24.3-30.6 mm in males).Leptobrachella yunyangensissp.nov.thus differs from the smallerL.baluensis(14.9-15.9 mm in males),L.brevicrus(17.1-17.8 mm in males),L.bondangensis(17.8 mm in a single adult male),L.fusca(16.3 mm in a single adult male),L.itiokai(15.2-16.7 mm in males),L.juliandringi(17.0-17.2 mm in males),L.mjobergi(15.7-19.0 mm in males),L.natunae(17.6 mm in a single adult male),L.parva(15.0-16.9 mm in males),L.palmata(14.4-16.8 mm in males),andL.serasanae(16.9 mm in a single adult female).
By the moderate body size of the male (SVL 28.3-30.6 mm),Leptobrachella yunyangensissp.nov.differs from the largerL.damingshanensis(33.6-34.4 mm in males),L.eos(33.1-34.7 mm in males),L.nahangensis(40.8 mm in one male),L.neangi(35.4-36.3 mm in females),L.sungi(48.3-52.7 mm in males),L.tamdil(32.3 mm in one male),L.zhangyapingi(45.8-52.5 mm in males);from the smallerL.alpina(24.0-26.4 mm in males),L.applebyi(19.6-22.3 mm in males),L.ardens(21.3-24.7 mm in males),L.aspera(22.4 mm in one male),L.bashaensis(22.9-25.6 mm in males),L.bidoupensis(18.5-25.4 mm in males),L.crocea(22.2-27.3 mm in males),L.feii(21.5-22.8 mm in males),L.flaviglandulosa(23.0-27.0 mm in males),L.graminicola(23.1-24.6 mm in males),L.isos(23.7-27.9 mm in males),L.khasiorum(24.5-27.3 mm in males),L.laui(24.8-26.7 mm in males),L.maculosa(24.2-26.6 mm in males),L.melica(19.5-22.7 mm in males),L.mangshanensis(22.2-27.8 mm in males),L.murphyi(23.2-24.9 mm in males),L.niveimontis(22.5-23.6 mm in males),L.pallida(24.5-27.7 mm in males),L.pluvialis(21.3-22.3 mm in males),L.rowleyae(23.4-25.4 mm in males),andL.tengchongensis(23.9-26.0 mm in males).
For 15 species from the same lineage,i.e.,L.oshanensis,L.eos,L.purpurus,L.alpina,L.dorsospina,L.wulingensis,L.bourreti,L.yeae,L.graminicola,L.niveimontis,L.purpuraventra,L.suiyangensis,L.jinshaensis,L.chishuiensis,andL.bijie,Leptobrachella yunyangensissp.nov.could be easily distinguished by several characters.The new species differs fromL.eos(SVL 33.1-34.7 mm in males),L.alpina(SVL 24.0-26.4 mm in males),L.graminicola(SVL 23.1-24.6 mm in males),andL.niveimontis(SVL 22.5-23.6 mm in males) by moderate body size in the male (SVL 28.3-30.6 mm);fromL.eosby the presence of black spots on the flanks(vs.their absence),narrow lateral fringes on the toes (vs.wide),and light grayish creamy white interspersed with light brown spots (vs.creamy white);fromL.oshanensisandL.jinshaensisby rudimentary webbing on the toes (vs.lacking);fromL.purpurusandL.graminicolaby narrow lateral fringes on the toes (vs.wide) and indistinct dorsolateral markings (vs.distinct);fromL.dorsospinaby rough skin with sparse large warts and with short longitudinal ridges (vs.relatively smooth skin,some with small warts),indistinct dorsolateral markings (vs.distinct),and tibiotarsal articulation reaching to the anterior corner of the eye when the leg is stretched forward (vs.reaching to the posterior corner of the eye);fromL.wulingensisby indistinct dorsolateral markings (vs.distinct),tibiotarsal articulation reaching to anterior corner of the eye when the leg is stretched forward (vs.reaching to the middle of the eye),and light grayish creamy white interspersed with light brown spots (vs.creamy white with distinct or indistinct brown speckling at margins);fromL.bourretiandL.yeaeby rough skin with sparse large warts and with short longitudinal ridges (vs.relatively smooth skin) and indistinct dorsolateral markings (vs.distinct);fromL.purpuraventra,L.suiyangensis,L.chishuiensis,andL.bijieby indistinct dorsolateral markings (vs.distinct),tibiotarsal articulation reaching to anterior corner of the eye when the leg is stretched forward (vs.reaching to the middle of the eye or reaching the tympanum).
Leptobrachella yunyangensissp.nov.differs fromL.aerea,L.botsfordi,L.crocea,L.isos,L.pallida,L.petrops,L.tuberosa,andL.zhangyapingibythepresence of black spots on flanks (vs.absent in the latter).
Leptobrachella yunyangensissp.nov.differs fromL.ardens,L.kalonensis,L.shiwandashanensis,andL.tadungensisby rudimentary webbing on toes (vs.lacking toe webbing in the latter) and fromL.pelodytoides,L.sungi,andL.tamdil(vs.wide toe webbing in the latter).
Leptobrachella yunyangensissp.nov.differs fromL.lateralis,L.macrops,L.minima,L.namdongensis,L.nyx,L.pyrrhops,andL.ventripunctatabynarrow lateral fringes on toes (vs.lacking in the latter) and fromL.murphyi,L.yingjiangensisandL.yunkaiensis(vs.wide lateral fringes in the latter).
Leptobrachella yunyangensissp.nov.differs fromL.applebyi,L.ardens,L.bidoupensis,L.kalonensis,L.melica,L.minima,L.nahangensis,L.namdongensis,L.shangsiensis,andL.tadungensisby the dorsal skin being rough with numerous tubercles and short longitudinal ridges (vs.dorsum smooth);fromL.bourreti(vs.dorsum relatively smooth with small warts),L.fuliginosa(vs.dorsum smooth with fine tubercles),L.mangshanensis(vs.dorsum nearly smooth),L.nokrekensis(vs.dorsum tubercles and longitudinal folds),andL.pelodytoides(vs.dorsum with small,smooth warts).
Leptobrachella yunyangensissp.nov.differs fromL.damingshanensis,L.dorsospina,L.maoershanensis,L.puhoatensis,L.purpuraventra,L.wuhuangmontis,andL.wulingensisby lacking distinct blackish dorsolateral markings (vs.distinct blackish dorsolateral markings in the latter).
Currently,only one species of the genusLeptobrachella,namely,L.oshanensis,is distributed in Chongqing City,China,and this species can be distinguished by a combination of the following morphological characters.Leptobrachella yunyangensissp.nov.differs fromL.oshanensisby having rudimentary webbing and narrow lateral fringes on the toes (vs.lacking),dorsal skin shagreened with numerous tubercles and short longitudinal ridges (vs.dorsum smooth with few glandular ridges),tibiotarsal articulation reaching to anterior corner of the eye when the leg is stretched forward (vs.reaching to the middle of the eye),and lacking distinct blackish dorsolateral markings(vs.distinct blackish dorsolateral markings).The mean values of SNT,IOD,IND,NEL,TIB,TW,FOT are significantly greater than those ofL.oshanensisfor males of the new species (Table 4).In addition,in males the mean values of SNT/HDL,SNT/EYE,IOD/UEW,TIB/SVL,and TIB/HLL were significantly larger inLeptobrachella yunyangensissp.nov.than inL.oshanensis(P-values < 0.05;Table 4).
Distribution and ecologyLeptobrachella yunyangensissp.nov.is only known from the type locality of Lianhua Village,Renhe Town,Yunyang County,Chongqing,China and Qiyaoshan Nature Reserve,Chongqing,China,at elevations of 947-1200 m.Individuals of this new species are mostly found in grasses and shrubs near streams.In areas where the new species was found,Leptobrachella yunyangensissp.nov.is sympatric withPanophrys baolongensis(Ye,Fei and Xie,2007),Rana omeimontis(Ye and Fei,1993),Odorrana hejiangensis(Deng and Yu,1992),Polypedates braueri(Vogt,1911),Hyla annectans(Jerdon,1870),andFejervarya multistriata(Hallowell,1860).These species were often found in the same streams asLeptobrachella yunyangensissp.nov.Males were not heard calling during the field survey from June 20 to 29,2021.
4.Discussion
Phylogenetic analyses based on mitochondrial 16S rRNA and six nuclear genes suggested that the specimens collected in this study belonged to the genusLeptobrachellabut were distinct from all previously described species for the molecular marker mitochondrial 16S rRNA,the minimum distance betweenLeptobrachella yunyangensissp.nov.and its congeners was 4.8% (vs.L.bourreti),greater than the current threshold of 3.0% difference between amphibian species (Fouquetet al.,2007).In fact,this is much greater than the genetic divergence between the recognized species of the genusLeptobrachella(Table S2).Morphologically,Leptobrachella yunyangensissp.nov.has a number of features to distinguish it from congeners;for example,the presence of supra-axillary and ventrolateral glands,medium body size,the presence of black spots on the flanks,lacking distinct dorsolateral markings,and narrow lateral fringes on the toes (see the Comparison section for details).Thus,the combined morphological and genetic divergence data support the validity ofLeptobrachella yunyangensissp.nov.as a new species.

Table S1 Primers used in PCR and sequencing in this study.




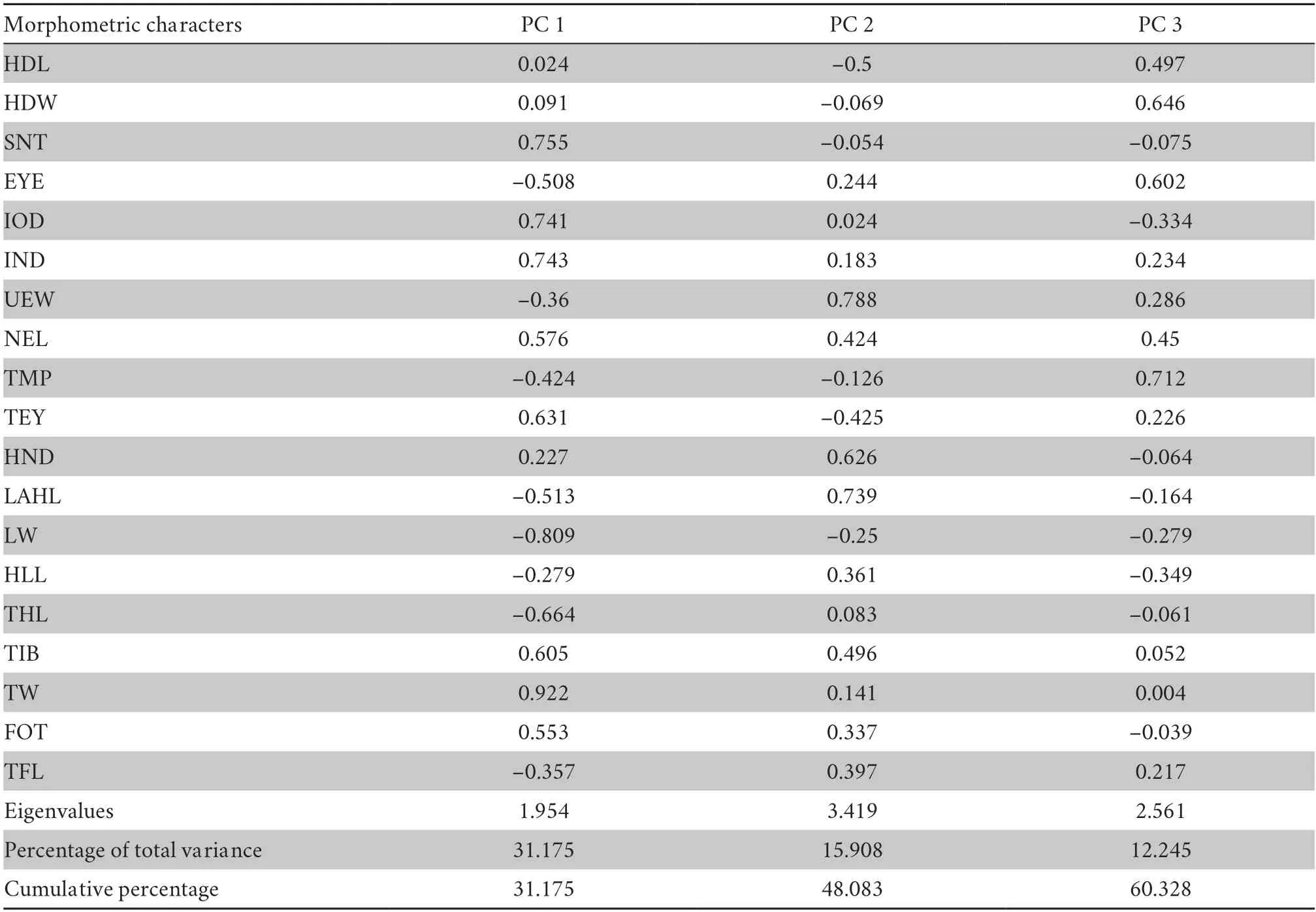
Table S3 Variable loadings for principal components with eigenvalues greater than 1 from morphometric characters corrected by SVL.


In the past decade,24 new species of the genusLeptobrachellahave been discovered in southern China (Sunget al.,2014;Yanget al.,2016;Yuanet al.,2017;Yanget al.,2018;Wanget al.,2018;Houet al.,2018;Chenet al.,2019;Wanget al.,2019;Wanget al.,2020;Chenet al.,2020;Liet al.,2020;Luoet al.,2020;Lyuet al.,2020;Qianet al.,2020;Chenet al.,2021a,b;Chenget al.,2021;Shiet al.,2021),and the discovery of these new species has deepened our understanding of their diversity and evolutionary history.The work of Chenet al.(2018) provides a good framework for the phylogenetic study of the genusLeptobrachella.Unfortunately,this large-scale phylogenetic study included only one sample from Nanchuan,Chongqing,China,and the limited sample size may have led to an underestimation of species diversity.The discovery site ofLeptobrachella yunyangensissp.nov.is located in the transition zone of biodiversity extending from the mountains of southwest China to the Qinba Mountains,and the discovery of the new species provides important additional material for understanding the detailed evolutionary history and biogeographic patterns of the genus.The continued field surveys are important for understanding and conserving the biodiversity of the region.In addition,because this group is highly dependent on forest habitat and is temperature sensitive,it is particularly important to protect its macroecologydependent microhabitats.Therefore,in the context of global warming,there is an urgent need for a comprehensive,systematic,and in-depth survey of the impacts of climate change on terrestrial vertebrates to provide a basis for rational decisions regarding amphibian conservation (IPCC,2014).
AcknowledgementsThis research was supported by the programs of the Guizhou Province World-Class Discipline Construction Program Project (Qianjiao Keyan Fa [2019]125),the Biodiversity Survey and Assessment in Chongqing Counties (2021-090),the Postgraduate Education Innovation Programme of Guizhou Province (Qianjiaohe YJSKYJJ [2021]091),the Strategic Priority Research Program B of the Chinese Academy of Sciences (CAS) (No.XDB31000000),the National Animal Collection Resource Center,China,the Application of Amphibian Natural Antioxidant Peptides as Cosmetic Raw Material Antioxidants (QKZYD [2020]4002).We thank Peifei Luo and Wei Jia for their help during specimen collection.We thank Prof.Daode Yang and Mr.Tianyu Qian for providing the nuclear gene data ofLeptobrachella wulingensis.We thank Accdon (www.accdon.com) for its linguistic assistance during the preparation of this manuscript.
Appendix
Specimens examined
Leptobrachella alpina(N=5):China:Yunnan Province:Jingdong County:Mt.Wuliang (type locality):GZNU20160623002-623006.Leptobrachella suiyangensis(N=8):China:Guizhou Province:Suiyang County:Huoqiuba Nature Reserve (type locality):
GZNU20180606007 (holotype),GZNU20180606001-606006,GZNU20180606008.
Leptobrachella wulingensis(N=2):China:Guizhou Province:Shibing County:Mt.Yuntai (topotype locality):GZNUSB20170501,GZNUSB20170503.
Frost D.R.,Grant T.,Faivovich J.,Bain R.H.,Haas A.,Haddad C.F.B.,de Sá R.O.,Channing A.,Wilkinson M.,Donnellan S.C.,Raxworthy C.J.,Campbell J.A.,Blotto B.L.,Moler P.,Drewes R.C.,Nussbaum R.A.,Lynch J.D.,Green D.M.,Wheeler W.C.2006.
The amphibian tree of life.Bull Am Mus Nat Hist,(297):1-370
Simon C.,Frati F.,Beckenbach A.,Crespi B.,Liu H.,Flook P.1994.Evolution,weighting and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers.Ann Entomol Soc Am,87(6):651-701
Vieites D.R.,Min M.S.,Wake D.B.2007.Rapid diversification and dispersal during periods of global warming by plethodontid salamanders.Proc Natl Acad Sci USA,104(50):19903-19907
Mauro D.S.,Gower D.J.,Oommen O.V.,Wilkinson M.,Zardoya R.2004.Phylogeny of caecilian amphibians (Gymnophiona) based on complete mitochondrial genomes and nuclear RAG1.Mol Phylogenet Evol,33:413-427.
Fu J.Z.,Weadick C.J.,Bi K.2007.A phylogeny of the high-elevation Tibetan megophryid frogs and evidence for the multiple origins of reversed sexual size dimorphism.J Zool,273(3):315-325
Shimada T.,Matsui M.,Yambun P.,Sudin A.2011.A taxonomic study of Whitehead’s torrent frog,Meristogenys whiteheadi,with descriptions of two new species (Amphibia:Ranidae).Zool J Linn Soc,161(1):157-183
Santos J.C.,Cannatella D.C.2011.Phenotypic integration emerges from aposematism and scale in poison frogs.P Natl Acad Sci USA,108(15):6175-6180
杂志排行

Asian Herpetological Research的其它文章
- A New Cyrtodactylus Species (Reptila:Gekkonidae) from Nan Province,NorthernThailand
- A New Species of Nidirana (Anura,Ranidae) from Southern Guangxi,China
- Spatial Patterns and Drivers of Chinese Lizard Richness among Multiple Scales
- Behaviours in Attachment-Detachment Cycles of Geckos in Response to Inclines and Locomotion Orientations
- Factors Influencing Home Ranges of the Qinghai Toad-headed Lizard(Phrynocephalus vlangalii) on the Dangjin Mountain,Gansu