A New Species of Nidirana (Anura,Ranidae) from Southern Guangxi,China
2022-06-25WeicaiCHENWanxiaoPENGPengLIandYoujunLIU
Weicai CHEN,Wanxiao PENG,Peng LI and Youjun LIU
1 Key Laboratory of Environment Change and Resources Use in Beibu Gulf Ministry of Education,Nanning Normal University,Nanning 530001,Guangxi,China
2 Guangxi Key Laboratory of Earth Surface Processes and Intelligent Simulation,Nanning Normal University,Nanning 530001,Guangxi,China
3 Shiwandashan National Nature Reserve,Fangchenggang 538000,Guangxi,China
Abstract A new music frog,Nidirana shiwandashanensis sp.nov.,is described from Shiwandashan National Nature Reserve,Shangsi County,Guangxi,China.Phylogenetic analysis based on mtDNA sequences indicates that the new species is closest to N.guangxiensis. However,N.shiwandashanensis sp.nov.differs from its congeners based on morphological characters,molecular data,and bioacoustics.The advertisement calls are comprised of 6-8 double notes,which are distinct from its sister taxon,N.guangxiensis.Currently,the new species occurs in the evergreen forest at Shiwandashan National Nature Reserve between 500-600 m a.s.l.
Keywords bioacoustics,morphology, Nidirana,phylogeny
1.Introduction
The genusNidiranaDubois,1992 is distributed in eastern and southeastern Asia (AmphibiaChina,2021;Frost,2021).For a long timeNidiranahas been regarded as a subgenus ofRanaLinnaeus,1758.Based on large-scale species sampling and comprehensive morphology,molecular analysis,and bioacoustics data,Lyuet al.(2017) suggested that the genusNidiranais a distinct genus and a sister taxon to the genusBabina.In recent years,Nidiranaspecies diversity increased dramatically from 7 to 16 (AmphibiaChina,2021;Frost,2021),namelyN.guangdongensisLyu,Wan and Wang,2020;N.guangxiensisMo,Lyu,Huang,Liao and Wang,2021;N.leishanensisLi,Wei,Xu,Cui,Fei,Jiang,Liu and Wang,2019;N.mangveniLyu,Qi and Wang,2020;N.nankunensisLyu,Zeng,Wang,Lin,Liu and Wang,2017;N.occidentalisLyu,Yang and Wang,2020;N.xiangicaLyu and Wang,2020;N.yaoicaLyu,Mo,Wan,Li,Pang and Wang,2019;andN.yeaeWei,Li,Liu,Cheng,Xu and Wang,2020.This high frequency of new species descriptions indicates that the species diversity ofNidiranawas and most like still is substantially underestimated.
Due to conservative phenotypes,Nidiranaspecies from southern China were often treated asNidirana adenopleura(Lyuet al.,2017,2019,2020a,2020b).Currently,there are four recognizedNidiranaspecies in Guangxi,i.e.:N.guangxiensis,N.leishanensis,N.xiangica,andN.yaoica(AmphibiaChina,2021;Lyuet al.,2021),withN.guangxiensisandN.yaoicahaving usually been identified asN.adenopleura(Lyuet al.,2019,2021).In 2021,we surveyed Shiwandashan National Nature Reserve(SWDS),Shangsi County,Guangxi,China,and collected sevenNidiranaspecimens.After analyzing the specimens based on morphology,phylogeny,and bioacoustics,we found that these specimens differed from the knownNidiranaspecies.Herein,we describe the specimens as a new species ofNidirana.
2.Materials and Methods
2.1.SpecimensFour adults,one sub-adult,and two tadpoles were collected in SWDS.All specimens were fixed in 10%formalin for 48 h,then transferred to 75% ethanol,finally deposited in Nanning Normal University.Muscle samples were stored in 100% ethanol for molecular analysis.
2.2.Morphological examinationThe definition of morphological characters refers to Lyuet al.(2021).The following measurements were taken with digital calipers to the nearest 0.1 mm:SVL,snout-vent length (from the tip of snout to posterior margin of the vent);HDL,head length (from the tip of snout to the articulation of the jaw);HDW,head width(head width at the commissure of the jaws);SNT,snout length(from the tip of snout to the anterior corner of the eye);IND,internasal distance (distance between nares);IOD,interorbital distance (minimum distance between upper eyelids);ED,eye diameter (from the anterior corner of the eye to posterior corner of the eye);TD,tympanum diameter (horizontal diameter of tympanum);TED,tympanum-eye distance (from the anterior edge of tympanum to posterior corner of the eye);HND,hand length (from the proximal border of the outer palmar tubercle to the tip of digit III);RAD,radio-ulna length (from the flexed elbow to the proximal border of the outer palmar tubercle);FTL,foot length (from the distal end of the shank to the tip of digit IV);TIB,tibial length (from the outer surface of the flexed knee to the heel).
The webbing formula follows Savage (1975);tadpole stage was identified following Gosner (1960);sex was determined by examining the nuptial pad.Comparison data were obtained from collected specimens (Appendix 1) and the literature(Boettger,1895;Boulenger,1904,1909;Bourret,1937;Chang and Hsu,1932;Chou,1999;Chuaynkernet al.,2010;Feiet al.,2007,2009;Kuramoto,1985;Liet al.,2019;Lyuet al.,2017,2019,2020a,2020b,2021;Matsui,2007;Schmidt,1925;Weiet al.,2020).
2.3.Phylogenetic analysisMaximum likelihood (ML) and Bayesian inference (BI) methods were used to analyze the phylogenetical relationships based on partial 16S ribosomal RNA gene (16S,~1050 bp) and partial cytochrome oxidase subunit I gene (COI,~640 bp).We referred to Lyuet al.(2017)to isolate and amplify the two mitochondrial DNA fragments.Homologous sequences ofNidiranaspecies,mostly from type localities (Table 1),were downloaded from GenBank for phylogenetic analysis.ML was conducted on the CIPRES science gateway with default settings (https://www.phylo.org/portal2/home.action#).BI was implemented using MrBayes v3.2(Ronquistet al.,2012),and the best-fit model (GTR+I+G) and running parameters following Chenet al.(2021).
2.4.Bioacoustics analysisAdvertisement calls were recorded in the field at the ambient temperature of 21°C with a SONY ICD-TX50 recorder on 10 May 2021.The software Raven Pro 1.6 (Cornell Laboratory of Ornithology,USA) was used to analyze the call recordings as per Köhleret al.(2017).
3.Results
ML and BI analyses of a matrix containing 1775 bp generated identical topologies,consistent with Lyuet al.(2021) (Figure 1).Preliminary phylogenetical trees indicate that specimens of SWDS represent a monophyletic group and are closest toN.guangxiensiswith robust node supports (Figure 1).
Sixteen advertisement calls were recorded in the field.A call group contains 6-8 calls with double notes (Figure 2).Call duration ranges from 1.35 s to 2.11 s (mean 1.66±0.21 s,n=16),whereas call interval ranges from 5.68 s to 13.10 s (mean 7.76± 1.92 s,n=15).The dominant frequencies of double notes are 0.36-2.64 kHz and 0.43-3.25 kHz,respectively.The calls differ from their congeners with regular 6-8 double notes.Nidirana guangxiensiscalls consist of 6-11 rapidly repeated identical and regular single notes (Lyuet al.,2021) (Figure 2).Nidirana daunchina,N.lini,N.nankunensis,N.xiangica,andN.yeaeissue a significantly different first note (Chou,1999;Lyuet al.,2017;Lyu and Wang,2020).Nidirana adenopleura,N.chapaensis,N.guangdongensis,N.leishanensis,N.occidentalis,N.pleuraden,andN.yaoicahave calls without double notes and there are less than six calls in a group (Boulenger 1904;Lyuet al.,2020).Nidirana hainanensisissues 2-4 fast-repeated double-notes (Lyuet al.,2017;Liet al.,2019).
Molecular analyses,bioacoustic data,and morphological evidence (see taxonomic account section) indicate that the specimens from SWDS represent a so far unrecognizedNidiranaspecies,which is described as follows.
Taxonomic account
Nidirana shiwandashanensis sp.nov.(Figure 3)
HolotypeNNU00238,adult male,collected from Shiwandashan National Nature Reserve,Shangsi County,Guangxi,China on 10 May 2021,by WeiCai CHEN.
ParatypeNNU00239 and NNU00605,adult males,NNU00606,adult female,collected from the same locality of holotype.NNU00239 collected on 10 May 2021;NNU00605 and NNU00606 collected on 24 August 2021.WeiCai CHEN collected all specimens.
EtymologyThe species name ‘shiwandashanensis’ refers to the type locality,Shiwandashan National Nature Reserve.We suggest the English name as Shiwandashan Music Frog,and the Chinese name as Shi Wan Da Shan Qin Wa (十万大山琴蛙).
DiagnosisNidirana shiwandashanensissp.nov.differs from its congeners in the combination of the following characters:medium body size (SVL 46.2-50.8 mm in males;48.3 mm in female);dorsal skin smooth without tubercles;ventral surface near immaculate creamy-white without spots;lateroventral grooves present on all fingers and toes;tibiotarsal articulation reaching eye;a call group containing 6-8 calls with double notes.
Description of holotypeAdult male,SVL 46.2 mm;head length almost equal to width (HDL/HDW=1.06);snout oval,significantly protruding beyond lower jaw;canthus rostralis distinct;loreal region sloping;nostril oval and closer to snoutthan eye,laterally opening;a creamy white longitudinal ridge present,beginning at posterior of nostril along with upper lip and ending at above insertion of arm;supratympanic fold absent;IOD/IND=0.68;eye diameter larger than tympanic diameter (ED/TD=1.36);vomerine teeth oval,closer each other than choana;tongue with a deep notch on posterior;a pair of subgular internal vocal sacs present.Relative finger lengths:II< IV < I < III;tips of fingers slightly dilated with lateroventral grooves;finger webbing and dermal fringes absent;subarticular tubercles prominent and conical;three distinct palmar tubercles;nuptial pad present on finger I.Relative toe lengths:I < II < V <III < IV;tip of toes dilated,forming an arrow-shaped disc with lateroventral grooves;toe webbing formula:I 2 -2½+II2½--3 III 2+-4½-IV 4 -3-V;toes with lateral fringes;subarticular tubercles prominent and oval;inner metatarsal tubercles elongated,but outer metatarsal tubercles indistinct;heels not meeting when thighs held at right angles to body;tibiotarsal articulation reaching eye.Dorsum smooth without tubercles on posterior;hindlimbs with several longitudinal dermal ridges;dorsolateral fold beginning at posterior of eye and ending at above groin and irregular small glands present along its inferior margin;pineal gland distinct;flanks with a single gland each;vent with some small tubercles around (Figure 3).

Figure 1 Bayesian Inference tree illustrating phylogenetic relationships among Nidirana based on the mitochondrial 16S and COI genes.Numbers at nodes are posterior probabilities (left) and maximum likelihood bootstrap values (right) from a separate Maximum likelihood analysis of the same dataset.* represents type locality.

Figure 2 Advertisement calls of Nidirana shiwandashanensis sp.nov. and N.guangxiensis (NNU00213) from the type localities.
Color of holotypeIn life,dorsum light brown with several sparse black spots;pineal gland creamy-white;dorsum medially with a creamy-white line,beginning at pineal gland and ending at vent;presence of a black line beginning at tip of snout,along the inferior margin of dorsolateral fold and ending at groin;tympanum brown with a black marking at posterior;presence of a creamy-white linear gland on upper jaw;maxillary gland creamy-white;flanks with several black spots and a large greyish-yellow glandular area;thigh and tibia with three grey bands;ventral surfaces of throat,chest,belly,and thigh near immaculate creamy white;anterior of base of forelimb with a dark stripe.In preservation,dorsal surface fading to deep grey;black spots displaying darkish black.
VariationsMeasurements of type series are listed in Table 2.Paratypes are similar to holotype in morphology and color pattern.The colorations of transverse bands on thigh and tibia vary from grey to dark.Spots on dorsal surface of thigh are variable,some having more dense spots than others.Black spots on dorsum vary too.Female individual (NNU00606) lacks large greyish-yellow glandular areas on flanks.
TadpoleIn the 40thstage (NNU00600),body length 16.8 mm,tail length 33.9 mm,maximal tail height 10.2 mm,eyes lateral,spiracle sinistral,dorsoposterior directed,dorsal fin arising at the base of the tail,labial tooth row formula:I:1+1/1+1:II (Figure 3F).In life,body brown,tail pale brown with pigmented tiny spots;body and tail grey in preservation.
Ecology and distributionNidirana shiwandashanensissp.nov.was found in the swamp at evergreen forests between 500-600 m a.s.l.,where there was no running water.On 10 May 2021,we heard the advertisement calls in the field but have not found eggs.On 24 August 2021,we revisited the same site and found the tadpole in 40thstage and the sub-adult specimen (Figure 3G,H).We assume that the breeding season ranges from 5 to 7 months.We did not observe the new species has nest construction behavior.So far,we only found this new species in SWDS.Whether it also occurs in the adjacent reserve,Golden Camellia National Nature Reserve,or has an even larger actual range,had to be determined through further fieldwork.
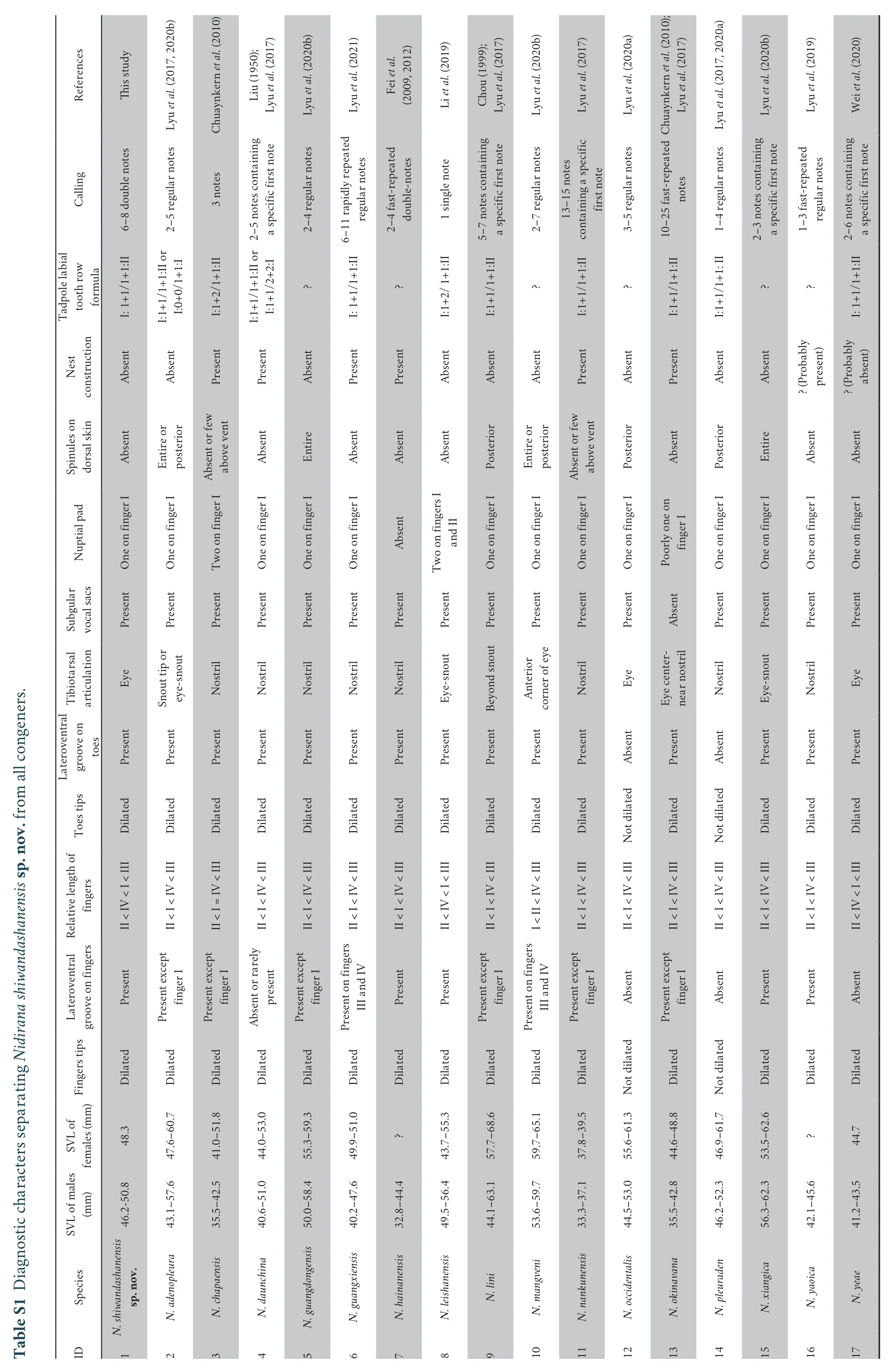
ComparisonA summary of morphological characters is provided in Table S1.Nidirana shiwandashanensissp.nov.differs from its congeners based on the following characters:(1) SVL 46.2-50.8 mm in males;(2) dorsal skin smooth without tubercles;(3) ventral surface near immaculate creamy-white;(4)tibiotarsal articulation reaching eye;(5) relative finger length II < IV < I < III;(6) lateroventral grooves present on all fingers and toes;(7) nuptial pad present on Finger I;(8) a call containing double notes.
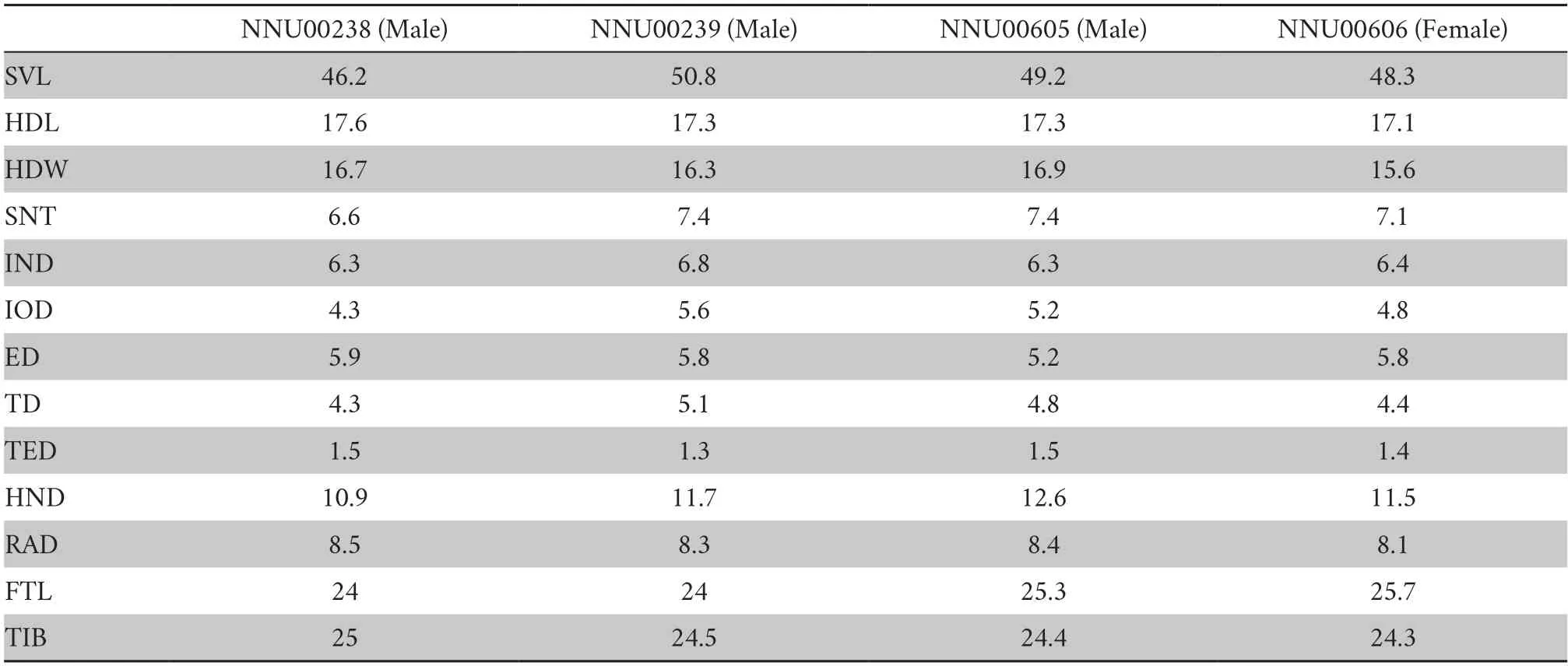
Table 2 Measurements of Nidirana shiwandashanensis sp.nov.(in mm).Abbreviations defined in Materials and Methods.
Phylogenetically,N.shiwandashanensissp.nov.is closest toN.guangxiensis(Figure 2).However,N.shiwandashanensissp.nov.differs fromN.guangxiensisby relative fingers length II < IV < I< III (vs.II < I < IV < III);tibiotarsal articulation reaching eye (vs.tibiotarsal articulation reaching nostril);absent dermal fringes on fingers (vs.broad lateral fringes on inner sides of Fingers II,III and IV but absent on Finger I);lateroventral grooves present on all fingers (vs.lateroventral grooves present on Fingers III and IV);toe webbing formula:I 2 -II2½--3 III 2+-4½-IV 4 -3-Vsmooth dorsal skin without tubercles (vs.rough dorsal skin with dense granules);a call containing 6-8 double notes (vs.6-11 rapidly repeated identical and regular notes) (Figure 2).

Figure 3 A-H:Nidirana shiwandashanensis sp.nov.A:dorsal view;B:lateral view;C:ventral view;D:ventral view of foot;E:ventral view of hand;F:labial tooth row formula;G:tadpole in the 40th stage;H:sub-adult.
By its medium body size (SVL 46.2-50.8 mm in males),N.shiwandashanensissp.nov.differs fromN.nankunensis(vs.SVL < 38 mm in males);by relative finger lengths II < IV < I< III,N.shiwandashanensissp.nov.differs fromN.leishanensisandN.yeae(vs.II < IV < I < III),N.chapaensis(II < I=IV < III),andN.mangveni(vs.I < II < IV < III);by lateroventral grooves present on all fingers (vs.absentN.yeae,N.occidentalisandN.pleuraden;present all but finger IN.chapaensis,N.adenopleura,N.guangdongensis,N.lini,N.nankunensis,andN.okinavana);by lateroventral grooves present on all toes (vs.absent on all toes inN.occidentalisandN.pleuraden);by tibiotarsal articulation reaching eyes (vs.beyond the tip of snout inN.lini;at nostril inN.adenopleura,N.chapaensis,N.daunchina,N.guangdongensis,N.hainanensis,N.nankunensis,N.pleuraden,andN.yaoica);by spinules absent on posterior dorsal skin (vs.present inN.adenopleura,N.lini,N.mangveni,N.occidentalis,N.pleuraden,andN.xiangica);by single nuptial pad present on Finger I (vs.absent inN.hainanensis;two part on Finger I inN.chapaensis;present on Fingers I and II inN.leishanensis) (Table S1).
4.Discussion
Comprehensive analyses,including morphology,DNA sequences,and bioacoustics,strongly support identifying the herein described specimens from SWDS as a new species in the genusNidirana.Based on the mtDNA analysis,N.shiwandashanensissp.nov.andN.guangxiensisare sister taxa.Although the two species have similar morphological characters,N.shiwandashanensissp.nov.andN.guangxiensisdisplay clearly distinct bioacoustics and nest construction behavior.The advertisement calls of the new species consist of 6-8 calls with double notes,whereas the calls ofN.guangxiensiscontain 6-11 rapidly repeated identical and regular single notes(Figure 2).Additionally,N.shiwandashanensissp.nov.has not to nest construction behavior,other thanN.guangxiensis.
Up to date,there are five recognizedNidiranaspecies in Guangxi,displaying impressive species diversity.Our discovery ofN.shiwandashanensissp.nov.demonstrates that previous studies treating GuangxiNidiranapopulations solely asN.adenopleurawere erroneous (Liu and Hu,1962;Feiet al.,2009;Moet al.,2014;Zhang and Wen,2000).Lyuet al.(2021) and our results both indicate thatNidiranaspecies have relatively narrow ranges and that rivers,and mountains contribute to the speciation ofNidiranaspecies.For instance,Xijiang Rive separatesN.shiwandashanensissp.nov.andN.guangxiensis,whereas Hongshuihe Rive separatesN.guangxiensisandN.yaoica.Nowadays,noN.adenopleurahave been found in northern,southern,eastern,and central Guangxi.WhetherN.adenopleuraoccurs in western Guangxi needs to be investigated further.
AcknowledgmentsThe authors are grateful to the staff of Shiwandashan National Nature Reserve for assistance during fieldwork and two anonymous reviewers for improving the manuscript.Additionally,the authors would like to express their gratitude to EditSprings (https://www.editsprings.cn/) for the expert linguistic services provided.This work was supported by the National Natural Science Foundation of China(32060116) and Guangxi Natural Science Foundation,China(2020GXNSFDA238022).
Appendix
杂志排行

Asian Herpetological Research的其它文章
- A New Species of the Asian Leaf Litter Toad Genus Leptobrachella (Amphibia,Anura,Megophryidae) from Chongqing City,Southwest China
- A New Cyrtodactylus Species (Reptila:Gekkonidae) from Nan Province,NorthernThailand
- Spatial Patterns and Drivers of Chinese Lizard Richness among Multiple Scales
- Behaviours in Attachment-Detachment Cycles of Geckos in Response to Inclines and Locomotion Orientations
- Factors Influencing Home Ranges of the Qinghai Toad-headed Lizard(Phrynocephalus vlangalii) on the Dangjin Mountain,Gansu