Behaviours in Attachment-Detachment Cycles of Geckos in Response to Inclines and Locomotion Orientations
2022-06-25WeijiaZONGZhouyiWANGBingchengWANGZhourongZHANGChangYINandZhendongDAI
Weijia ZONG,Zhouyi WANG*,Bingcheng WANG,Zhourong ZHANG,Chang YIN and Zhendong DAI*
1 Institute of Bio-inspired Structure and Surface Engineering,Nanjing University of Aeronautics and Astronautics,Nanjing 210016,Jiangsu,China
2 Civil Aviation College,Nanjing University of Aeronautics and Astronautics,Nanjing 211106,Jiangsu,China
Abstract Geckos can move quickly in various environments by efficiently controlling their complex adhesive toe pads.The locomotion behaviours observed in the attachment-detachment (A-D) cycle of their toe pads in response to changes in their environment should be studied to understand the adaptive behavioural characteristics of such toe pads.The lack of systematic research on the entire A-D cycle,including the release,swing,contact,and adhesion stages,limits the comprehension of the adhesive locomotion mechanism.The A-D cycle of Gekko gecko that facilitates the foot locomotion on inclined and vertical surfaces was investigated to clarify the locomotion behaviours in different stages.Results show that the change trends of foot locomotor angles (yaw and pitch) during the entire A-D cycle remain unchanged in response to various substrates.The bending angles (fore 41°;hind 51°) and contact time percentages (fore 7.42%;hind 7.44%) in the contact stage as well as the forefoot angle ranges (yaw:163.09°;pitch:308.68°) in the A-D cycle also remain constant across all substrates.These invariant foot locomotion behaviours during the swing and contact stages suggest that the foot behaviours are weakly related to the forces acting on the foot,which change according to the environment.Furthermore,the forefoot and hindfoot have different anatomical structure and functional demands,thus,the angle range of forefoot locomotion is larger than that of hindfoot locomotion,and the pitch angle change trend of the forefoot is opposite to that of the hindfoot.The diverse and complex locomotion control of the adhesive toe pads for various environments is reduced by the consistent behaviours in the gecko’s A-D cycle,such as the constant postures in the swing and contact stages.This study provides insight into the adhesive locomotion mechanism of geckos and can facilitate further research on the effective design and control of adhesion robots.
Keywords attachment-detachment cycle,Gekko gecko,inclines,locomotion behaviours,locomotion orientation
1.Introduction
Locomotion results from the interaction forces between the attachment devices of organisms and the external environment(Dickinsonet al.,2000).Some insects (Liuet al.,2019),tree frogs(Endleinet al.,2013),and geckos,have evolved attachment devices with adhesion function to negotiate extremely challenging terrains.Geckos can adapt to various environments and move quite fast on steep slopes or even ceilings (Wanget al.,2015).The extraordinary adhesion ability of geckos results from the hierarchical adhesion structure (Autumn and Peattie,2002;Russell,1975;Zaafet al.,1999) and efficient locomotion control of their complex compliant toe pads (Russell,2002;Songet al.,2020).The adhesion relies not only on the microstructure(setae) but also on the macroscopic structure (toes and toe pads).Many researchers have focused on the adhesion mechanisms of the micro-or mesoscale structures (setae,lamellar)(Autumnet al.,2006;Tianet al.,2006;Tianet al.,2013).Full comprehension of the adhesion-locomotion mechanism also requires an understanding of the dynamic adhesion behaviours of macrostructures in response to locomotion challenges in a complete attachment-detachment cycle,including the stance and swing phases that ensure effective attachment of the microstructures.This includes understanding how the toe pads contact,stand on,and release from the substrate as well as how they swing in air,especially for complex toe pads that respond to varying climbing moves.
The attachment-detachment (A-D) cycle can be divided into contact,adhesion,release,and swing stages,and the stable adhesion stage is directly related to the adhesion forces (Wanget al.,2018).Time (attach/detach time (Wanget al.,2011)) and space variables (toes orientation (Foster and Higham,2012)and angle (Wanget al.,2010)) are introduced to describe the foot locomotion behaviour.Many researchers have studied the adhesion stages.For example,the realisation of the adhesion depends on the distance (Arztet al.,2003),angle (Russell and Higham,2009;Autumnet al.,2000);force (Russell,2002,1981;Autumnet al.,2006) between the setae and the interface,and the toe pad deployment in the adhesion stage (Russell and Higham,2009;Songet al.,2020),although the contact,release and swing stages in the dynamic A-D cycle are often ignored.The locomotion behaviours of the toe pads in an entire A-D cycle and how they meet the adhesion requirements need to be explored.
The adhesion forces of the gecko’s toe pads differ in various environments (Autumn and Peattie,2002;Russell and Oetelaar,2016).The direction or magnitude of the adhesion forces is related to the deployment of toe pads in the adhesion stage(Russell and Higham,2009;Songet al.,2020;Wanget al.,2018).To adapt to various locomotion challenges,geckos’ foot position,orientation (Imburgiaet al.,2019;Russell and Oetelaar,2016;Wanget al.,2018),postures (Songet al.,2020),and locomotion speed (Russell and Higham,2009) alter.This means that the A-D cycle could satisfy a variety of locomotion requirements using an intergraded behavioural strategy,but the relationship between the toe pads adaptive behaviours and locomotion demands is not systematically analysed.
The forefoot of the gecko is different from the hindfoot in terms of physiological anatomy and adhesion potential (Russell,1975).The force patterns of the forefoot and hindfoot vary at different slopes (Wanget al.,2015).In addition,the locomotion function of the forefoot and hindfoot on a vertical surface in different directions (Wanget al.,2011) and the response to various inclinations (Birn-Jeffery and Higham,2014) are disparate.Therefore,it is necessary to investigate whether the variations in the physiological structure are related to the distinction in the adaptive behaviours of the A-D cycle of the forefoot and hindfoot.
Here,the A-D cycle ofGecko Gekkowith complex compliant toe pads on substrates at varying inclinations (level,45° and vertical surfaces) and locomotion orientations (uphill and downhill) was investigated to obtain the foot behaviours responding to changing environments.Gecko locomotion is recorded using a motion capture system,and the kinematics analysis is carried out,and then the toe pads’ locomotion behaviours during the A-D cycle for changing locomotion demands are summarized.Additionally,the contrasts and similarities in forefoot and hindfoot behaviours and the relationship between the toe pad structures and adhesion functions are studied.Thus,determination of the behavioural patterns in the A-D cycle of gecko’s toe pads for diverse locomotion demands will further our understanding of the adhesion-locomotion mechanism in the hierarchical adhesion structure.This can provide guidance for the design and control strategy of biomimetic adhesion robots.
2.Materials and methods
2.1.AnimalsExperimental geckos (Gekko gecko) were obtained from Guangxi Province (China).They were housed in a greenhouse with controlled humidity (65%-75%) and temperature (27±2) ℃.Five geckos with similar weight and body size were selected for climbing experiments (N=5;Mass=60.5±5.2 g;Body length=255.5±12.2 mm;Tail length=140.4 ±8.3 mm;Mean±SD).The selected geckos were fed with crickets every day and trained to climb on level,inclined and vertical surfaces for approximately two weeks.
2.2.Experimental protocolThe experimental setup has four parts:a rotatable motion platform with a locomotion channel,motion capture system,three-dimensional (3D) force measuring array and data processing platform (as shown in Figure 1A).The motion capture system mainly comprises four motion capture cameras (Prime 17 W,NaturalPoint,Inc.,Corvallis,OR,USA).The locomotion channel is constructed using acrylic plates and profiles and is 200 mm in width,allowing the geckos to move freely.It has an open end that serves as an entrance;the other end has a dark box to attract geckos.The 3D force platform comprises 24 force sensors with micron-level resolution,each of which can measure the reaction force of a different toe pad (Daiet al.,2011).
Following the marking technology proposed in (Wanget al.,2019) several markers are pasted on the gecko before each trial at the following positions:bregma,cervical spine,middle of spine,caudal vertebrae,left and right shoulders/hip,elbows/knees,wrists/ankles,middle and tip of the tail,and white nail polish was applied on the heel and tip of the middle toe (d3) of the forefoot and hindfoot as marking points (Figure 1C).
The rotatable locomotion platform was set to 0°,45° and 90°.Geckos were trained to climb the 90° and 45° surfaces from top to bottom and bottom to top,respectively.They were placed at the channel entrance and encouraged to move forward by squeezing the bottom of the tail and tapping the back until they climbed into the dark box.The locomotion behaviour of the gecko was recorded using the motion capture system with camera frame rate of 250 FPS.The foot reaction forces were measured using the 3D force measuring array with sampling rate of 500.
2.3.Data analysisTo obtain the x,y and z coordinates of the markers at the bregma,left and right shoulders/hip,cervical spine,caudal vertebrae,elbow/knee,wrist/ankle,heel and tip of the third toe of forefoot and hindfoot,the DLT6 program (Hedrick,2008) heeld on MATLAB 2018 platform(MathWorks Inc.,Massachusetts,USA) was used for digitising the recorded videos.The y,z,and x axes represent the forward and backward motion parallel to the direction of travel,the dorsoventral motion perpendicular to the moving surface,and the mid-lateral motion perpendicular to the y-z plane,respectively.
A global coordinate system (O0) was established,as shown in Figure 1A.The movement direction of the geckos is the positive direction of the y-axis,the x-y plane is parallel to the movement plane,and the direction perpendicular to the x-y plane is the positive direction of the z-axis.According to the Cartesian coordinate system,the x-axis is positive,and the movement direction is perpendicular to the y-z plane.The angle rotation around the x-and z-axis is the pitch and yaw angle,respectively.In addition,distinct coordinate systems (Olf,Orf,Olh,Orh) for the four feet are established (Figure 1C).The origin coordinates are set at the wrist/ankle joint;the x-y plane is parallel to the motion plane.The y-axis direction is consistent with the motion direction;the x-axis direction points to the outside of the body (the right and left foot points to the right and left,respectively).
2.4.Parameters definitionThe A-D cycle comprises the stance and swing phases.The stance phase can be divided into three stages:1) Contact (attach):the incipient period of contact is defined as the moment when the toe pad touches the substrate until the entire toe pad is in full contact with the heel.2) Adhesion:the stable period of contact represents the duration when the whole toe pad is in contact with the substrate and generates thrust.3) Release (detach):the release period of contact refers to the stage where the first toe begins to hyperextend and detach until all the toes leave the substrate (Figure 1E).The time spent in release,swing,contact,and adhesion stages (TR,TS,TC,TA) and the percentage of each of these periods are obtained.
The following angle definition takes the local coordinate system of each foot (Figure 1C) as the reference coordinate system.
Foot bending angleθ:After the toe pad goes the swing phase,the toe is rolled up when the toe pad touches the substrate and starts to adhere.The angle between the third toe (the line between toe tip and heel) and the positive direction of the y-axis in the y-z plane at the moment that the toe pad touches the substrate is defined as the bending angle (Figure 1D).It is an instantaneous parameter.
Bending angular velocity ω:The rotation speed of the third toe tip around the toe heel during the process from the toe heel to the toe tip contacting the substrate is obtained by dividing the bending angleϑby the contact process timeTC.
Foot pitch angleα:The angle between the third toe (the line between toe tip and heel) and the positive direction of the y-axis in the y-z plane during the process of the entire A-D cycle is defined as the pitch angle (Figure 1D).It is a process parameter.
Foot yaw angleγ:The angle between the third toe and the x-axis positive direction.A positive value indicates that the toe pad points in the positive direction of the y-axis,and a negative value indicates that the toe pad points in the direction opposite to the y-axis direction (Figure 1D).It is a process parameter.
Contact angleβ:The angle between the calf (the line between the wrist/ankle joint and elbow/knee joint) and the positive direction of y-axis in the y-z plane at the moment that the toe pad touches the substrate is defined as the contact angle (Figure 1D).It is an instantaneous parameter.
Initial contact force Fn_initial:This is the force perpendicular to the substrate surface when the toe pad firstly touches the substrate upon collision.It is an instantaneous parameter.
2.5.Statistical analysesAround sixty tests were chosen from more than 200,including 20 trials each of vertical-up,and vertical-down,10 trials each at 45° slope-up,45° slope-down,and horizontal.Three geckos participated in all experiments,and 3-5 trials of each experiment were selected.The other two geckos took part in the experiments of uphill-downhill vertical climbing,and also 3-5 trials of these two experiments were selected.There was no significant difference in the locomotion features between the two geckos and the other three gecko in the uphill-downhill vertical climbing experiments.Only trials in which the geckos moved in a straight path were picked,and force data from the feet that did not overlap were used.Every selected trial had a velocity of±15% of the average speed and at least a complete gait cycle video clip.
SPSS 19.0 (IBM Inc.,NY,USA) was used to analyse the selected data statistically.We used the Kruskal-Wallis test and one-way ANOVA analysis with the Scheffe method for post hoc tests for variables:contact/release time percentage,yaw angle,pitch angle,bending angle,foot contact angle,initial contact force,and bending angular velocity for different slopes(0°,45° and 90°) and motion directions (uphill and downhill).ANOVA with the Welch method and the Games-Howell test was used when the variance homogeneity was not satisfied.A significance level of 0.05 was used throughout.Spearman correlation analysis and generalized linear models was adopted to test the relationship between locomotion speed and time percentage of the contact and release stages.The results of the tests are expressed as the Mean±SD.
3.Results
3.1.Time percentages of different stagesThe four stages,release,swing,contact and adhesion,in an A-D cycle of the climbing gecko are depicted in Figure 2A-D.Figure 2E shows the time percentages of these four stages in vertical (uphill and downhill),45-degree slope (uphill and downhill),and level locomotion.
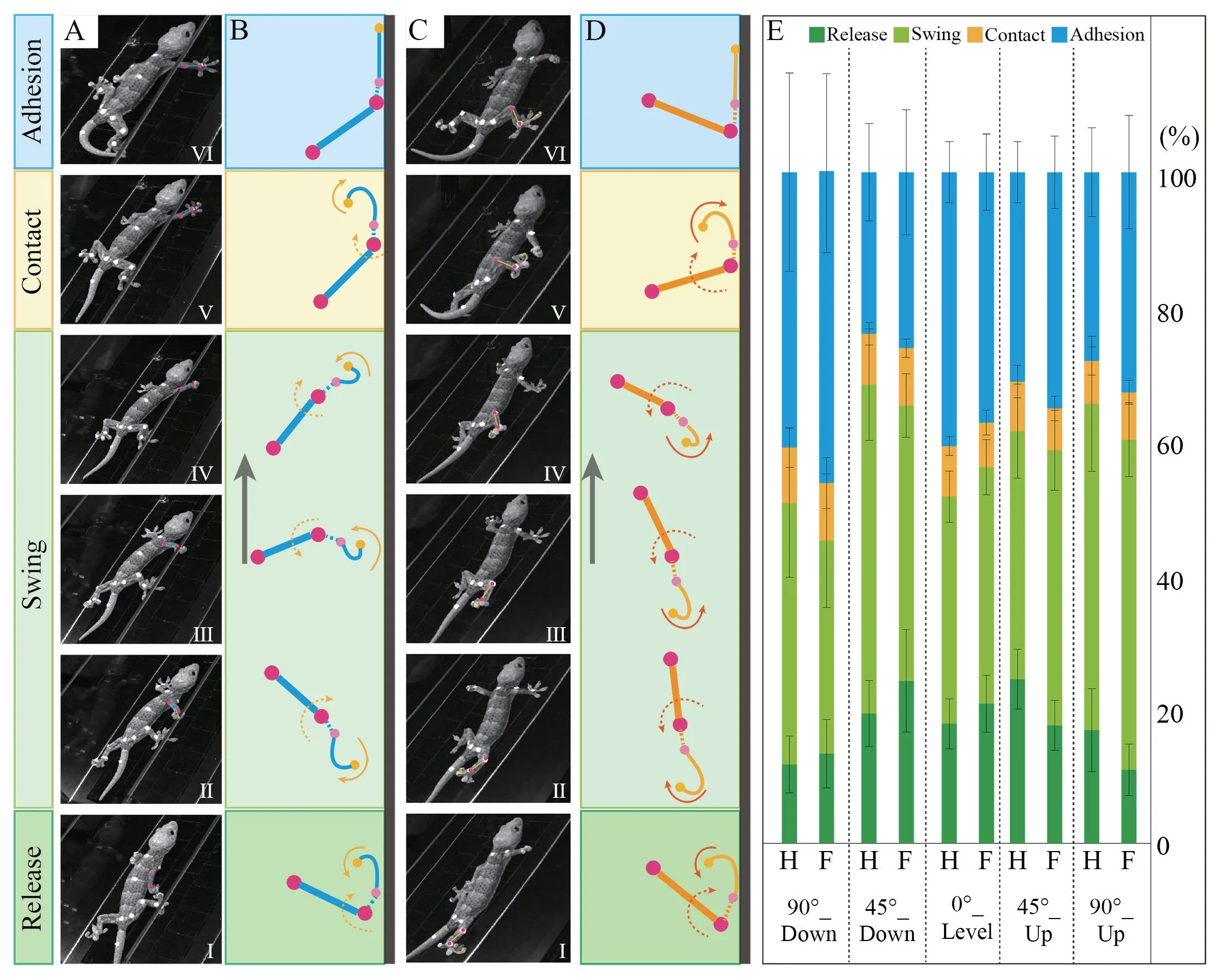
Figure 2 Locomotion postures and time percentages of different stages of forefoot and hindfoot locomotion in an A-D cycle.A-B:Snapshots and the corresponding simplified schematic diagrams (sagital view) of gecko’s forefoot locomotion in an A-D cycle;C-D:Snapshots and the corresponding simplified schematic diagrams (sagital view) of gecko’s hindfoot locomotion in an A-D cycle;E:Time percentage in each stage of an A-D cycle of forefoot and hindfoot in level and uphill and downhill locomotion on 45° and vertical surfaces.(H:represents hindfoot;F:represents forefoot).
Contact stage:The contact time percentages exhibit no significant difference on different slopes and motion directions(Forefoot:P=0.07;Hindfoot:P=0.28).This means that making the whole toe pad contact the substrate in varied locomotion conditions requires approximately the same time percentage.The contact time percentages of the forefoot (7.42%±0.97%) and that of the hindfoot (7.42%±0.61%) also exhibit no significant difference on different substrates (Supplementary Materials Table 6),indicating that the forefoot and hindfoot require similar time percentages for attachment in response to various locomotion conditions.
Release stage:The forefoot and hindfoot release time percentages varied significantly among substrates (Forefoot:P< 0.01;Hindfoot:P< 0.01),demonstrating that both the forefoot and hindfoot modify the release time percentage for detachment when coping with different locomotion conditions.When the gecko climbs downhill on the slope and wall or moves on horizontal plane,the release time percentage of the forefoot is slightly larger than that of the hindfoot,while the opposite is true when climbing up.
3.2.Locomotion posture changing trendFigure 2A-D and Figure 3E-F show the posture changes of the forefoot and hindfoot in a complete A-D cycle,including the adhesion,release,swing and contact stages.The dynamic postures of the forefoot and hindfeet exhibit clear differences during the swing stage:Forefoot toes are rolled up and detached at the beginning of the swing phase when the crus rotates toward the motion direction around the wrist joint.When the forefoot swings in the air following a swift detachment,all of the toes curl up and close together,and the toe pad changes the pitch angle dramatically,rising,then lowering,then rising again,and ultimately dropping to make contact with the substrate.During the swing stage,all toes on the hindfoot bend upward but are not clothed together noticeably.The hindfoot changes the yaw angle perceptibly.Specifically,the hind toe pad swings backward (against the motion direction) firstly,and then it swings forward until it is ready to make contact with the substrate for adhesion.

Figure 3 Yaw and pitch angle changing trend and locomotion postures of forefoot and hindfoot.A-D:Pitch and yaw angle changes during the four stages in the A-D cycle with schematic diagrams of foot motion postures above and below the curve.(n=10;Vertical-down climbing;Mean values of the yaw or pitch angles (Purple line;Variance,gray shadow);The vertical black dotted line is the dividing line of four different stages in the A-D cycle:Adhesion,Release,Swing and Contact);E-F:Schematic diagram of the A-D cycle postures of the (E) forefoot and (F) hindfoot in the frontal and sagital plane (Vertical-down climbing;The dark blue line represents the crus;the light blue line represents the third digits of the toe pad;Solid line arrow indicates digits rotation;Dotted arrow indicates the leg rotation).
The locomotion posture of the forefoot and hindfoot did not alter considerably under various locomotion situations,according to the video analysis and the changing trend of the locomotion angles in vertical,45° slope,and horizontal surfaces(Supplementary Materials Figure 1).The downhill locomotion on the vertical surface is chosen to exhibit the trend of the pitch and yaw angle over time during an A-D cycle (Figure 3A-D)as well as the entire process of forefoot and hindfoot postures(Figure 3E-F).The fluctuation trends of the foot yaw angle in various locomotion substrates are similar,with first a decrease and subsequently an increase.The shifting tendency of the forefoot pitch angle is more intricate than that of the hindfoot,and they are close to the opposite.The forefoot pitch angle lowers and then increases,whereas the hindfoot pitch angle increases and then decreases.
3.3.Locomotion posture angles
Yaw angle and pitch angleThe forefoot yaw angle range shows no significant change in different locomotion situations (F(4,45)=1.42,P=0.24),whereas that of the hindfoot has significant differences in different locomotion situations(F(4,22.5)=16.95,P< 0.001),which means that the hindfoot adjusts the rotation range (frontal plane) on different inclines.The forefoot yaw angle range is larger than that of the hindfoot in different locomotion situations except vertical-uphill and horizontal climbing (Figure 3A,Figure 4A and Supplementary Materials Table 6).
There is no difference in the minimum (Fore:-156.92° ±15.70°;K-W,χ2=5.67,df=4,P=0.23;Hind:164.00°±32.46°,F(4,45)=0.03,P=1.00) and the range (Fore:308.68°±20.42°;F(4,45)=2.57,P=0.05;Hind:164.00°±32.46°;K-W,χ2=8.57,df=4,P=0.07) of forefoot and hindfoot pitch angle on different locomotion situations (Figure 4B),indicating that the rotation movement of forefoot and hindfoot in the sagital plane remains nearly unchanged in varied locomotion conditions.Additionally,the pitch angle range of the forefoot is obviously larger than that of the hindfoot (as shown in Figure 4B and Supplementary Materials Table 6).
Foot bending angle and the angular velocityThe bending angles of the forefoot and hindfoot are 40.99°±5.01° and 50.91°±8.21° respectively;and they remain almost unchanged for different inclined plane locomotion,respectively (Fore:F(4,45)=1.26,P=0.30;Hind:K-W,χ2=3.78,df=4,P=0.44).The fore-and hindfoot bending angles show differences in various locomotion substrates (Supplementary Materials Table 9).The forefoot and hindfoot bending angular velocityωhas differences in various locomotion situations (Supplementary Materials Table 8).This means that the rotation speed of the toe tip around the toe heel is adjusted according to different locomotion conditions during the contact stage.ωof the forefoot is smaller than that of the hindfoot in different substrates(Figure 5B;Supplementary Materials Table 9).
Contact angleThe contact angle of the forefoot and hindfoot changes significantly on different locomotion situations(Fore:K-W,χ2=35.29,df=4,P< 0.001;Hind:F(4,21.10)=45.00,P< 0.001,Supplementary Materials Table 8),indicating that gecko adapted the angle between the crus and substrate at the moment that foot contacting the substrate to different locomotion situations.The forefoot contact angle differs from that of the hindfoot in different locomotion situations (Figure 5C;Supplementary Materials Table 9),and it is approximately half of that of the hindfoot during level walking.It also shows that the magnitude relation of the contact angle between the forefoot and hindfoot is related to the locomotion directions.Specifically,the forefoot contact angle (43.10°±6.33°) is larger than that of the hindfoot (32.21°±7.12°) when moving upwards vertically,whereas it is smaller (26.26°±3.05°) than that of the hindfoot (43.65°±5.98°) when moving downwards.Furthermore,the forefoot contact angle (17.12°±2.69°) is smaller than that of the hindfoot (50.88°±3.05°) when moving uphill on a 45° slope,whereas it (24.99°±5.60°) is larger than that of the hindfoot (7.46°±5.10°) when moving downhill.
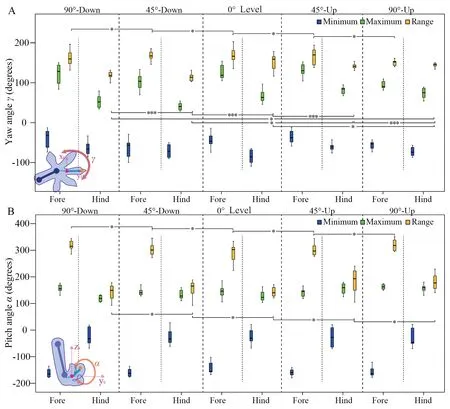
Figure 4 Maximum,minimum,and range of the yaw and pitch angles in the swing stage of forefoot and hindfoot when moving in the vertical plane (uphill and downhill),45° inclined plane (uphill and downhill) and horizontal plane.A:Yaw angle;B:Pitch angle.(n=10;Significance:* represents P ≥ 0.05,** represents 0.001 ≤ P < 0.05,*** represents P < 0.001).
4.Discussion
4.1.Structure-function relationship in different limbs“Structural and functional foundations” has been proposed as one of the promising areas for future work in terrestrial locomotion (Blob and Higham,2014).The anatomical structure and geometry of joints often reflect their locomotion and function.Single limb function has been studied through the assessment of both forelimbs and hindlimbs (Foster and Higham,2012;Wanget al.,2014).Additionally,the physiological structures of the forefoot and hindfoot have some similarities,which may be closely related to the requirements and functions of A-D locomotion.Firstly,the forefoot and hindfoot are capable of closed adhesion (Imburgiaet al.,2019).For instance,the propulsive function (braking/driving) of forefoot and hindfoot can be exchanged on the substrates of 0-180 degrees(Wanget al.,2015),which is similar to some adhesion insects,like ants (Wöhrlet al.,2017).Furthermore,the adhesion function and the physiological properties of toe pads result in the similar forefoot and hindfoot behaviours in different stages of the A-D cycle.The contact and release stages are crucial for adhesion.In the contact stage,for the quick generation and maintenance of adhesion,phalanges,blood,and other tissues inside the toes work together to spread the rolled up toes and apply normal and tangential force (Russell,1981);and in the release stage,the hyperextension bebaviour allows for easy and rapid detachment with a very small peeling force.The compliant structure of the toes allow the forefoot posture (close together movement) in the swing stages,which is likely to help reduce the resistance during swing stage.Likewise,the skeletal structure of the hindlimb leads to the posture of the hindfoot in the air,causing the large changes in the frontal plane but small changes in the sagital plane.Besides,geckos (Wanget al.,2018;Songet al.,2020)and tree frogs (Endleinet al.,2013;Endleinet al.,2017) adjust the deployment of the distributed toe pads to adapt to the force requirements of different environments.Distributed toes enable the foot to augment the normal adhesion force by coupling the reverse toes (Songet al.,2016).By contrast,during the swing stage,the forefoot bends and collects all toes,exhibiting the curling posture (Figure 3E),whereas the hindfoot does not have such obvious curling state (Figure 3F).It is conducive to preventing the adhesion until necessary.Furthermore,the bending angle of the forefoot is much less than that of the hindfoot in various locomotion situations (Figure 5B),which might be due to the smaller forefoot compared with the hindfoot (Russell,1975).Moreover,due to the segmented toe bone and bent phalanges (Russell,1981),the initial pitch angleαinitialdetermined by the positions of the toe tip and heel is always not zero (Supplementary Materials Table 9).
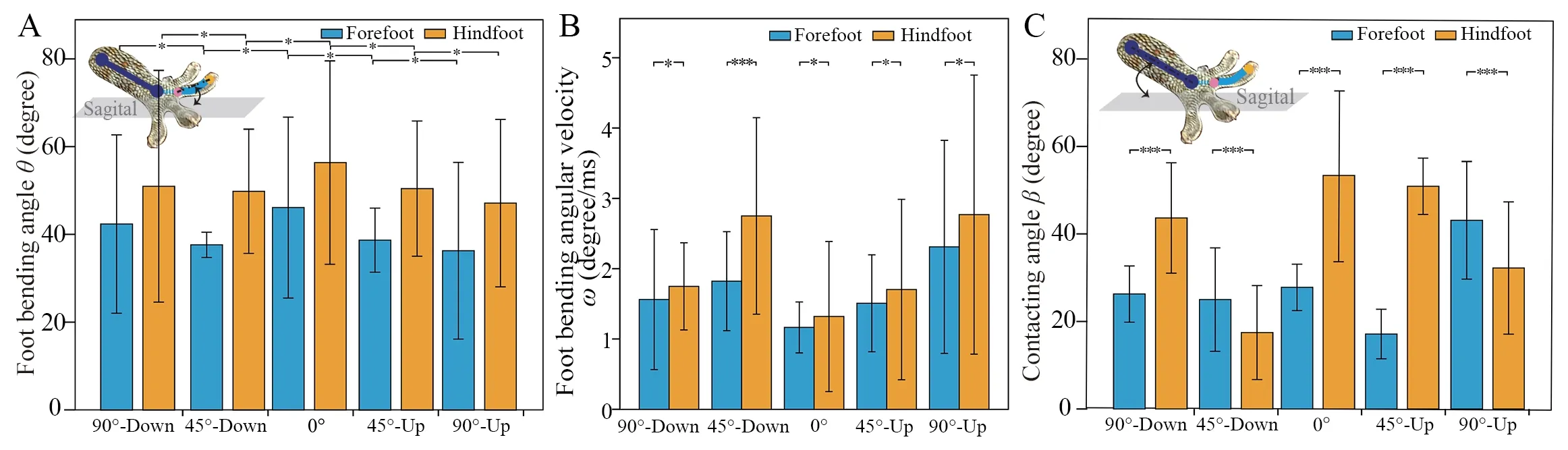
Figure 5 Mean locomotion angles in the contact moment of forefoot and hindfoot.A:Bending angles of the forefootand hindfoot when moving in the vertical plane (up and down),45° inclined plane (uphill and downhill) and horizontal plane;B:Forefoot and hindfoot bending angular velocity in the vertical and 45° inclined plane (uphill and downhill) and horizontal plane;C:Forefoot and hindfoot contact angles in the vertical and 45° inclined plane (uphill and downhill) and horizontal plane.(n=10;Significance:* represents P ≥ 0.05,** represents 0.001 ≤ P < 0.05,*** represents P < 0.001).
Experimental results reveal that the forefoot and hindfeet have distinct locomotion postures and the forefoot pitch and yaw angle ranges in the swing process are much larger than those of the hindfoot (Figure 3 and 4),which is related to the muscle tissue included in the forelimb that can rotate between the radius and ulna (see Figure 2 in (Russell and Bels,2001)) as well as the more sophisticated skeletal system of the forelimb relative to the hindlimb (as shown in Figure 3 in (Russell,1975)).Because the hindlimbs experience significantly greater movement alterations than the forelimbs (Birn-Jeffery and Higham,2014),our findings also show that the change in forefoot movement behaviour throughout an A-D cycle is smaller than the change in hindfoot movement behaviour.Concretely,on varied substrates,the hindfoot adjusts the rotation range (frontal plane),although the forefoot does not;hence the hindfoot yaw angle range is obviously wider during uphill locomotion than during downhill locomotion.
4.2.Intercoordination between adhesion status and limbfoot locomotionAdhesion function relies on the A-D cycle of the attachment devices.The posture and deployment of the toe pads in the adhesion stage is closely related to the adhesion force (Imburgiaet al.,2019;Wanget al.,2010;Songet al.,2020).As an A-D cycle is a dynamic process that includes the release,swing,contact,and adhesion stages,the intermediate contact process from swing to adhesion stage is essential for the stable adhesion.At the moment of initial contact process,there is often a collision forceFn_initial(the force between the foot and substrate at the moment of contact) (Wanget al.,2011) which may cause adhesion failure.TheFn_initialvaries on different substrates(Figure 6B) which is similar to the results on a vertical surface in different locomotion directions (upward,downward,left to right) (Wanget al.,2011).Theoretically,the changes in footmovement result in changes in force;however,the locomotion postures in the swing and contact stages of the forefeet and hindfeet on various substrates have no significant variation,including the angle changing trend,the bending angleϑ,and the contact time percentage.It indicates thatFn_initialhas little connection with the toe pad’s movement.Comparing theFn_initialof forefoot and hindfoot (Figure 6B) with the corresponding foot contact angle (Figure 5A) on different substrates,it was found that the contact angle of the forefoot and hindfoot and their contact force have the same magnitude relationship;that is,if the forefoot contact angle is greater than that of the hindfoot,theFn_initialof the forefoot is also larger than that of the hindfoot.The contact angle changes significantly with various locomotion situations (Supplementary Materials Table 10).The relationship between the foot contact angle and functional requirements of the forelimb and hindlimbs also changes.Fn_initialmeasured in our experiments is similar to the forces obtained by Wang’s team (Wanget al.,2011).Our results show that the magnitude-relationship between theFn_initialof the forefoot and hindlimbs and contact angle on the vertical and 45° when climbing upward and downward is exactly the opposite.Fn_initialis adjusted as the environment changes,however,the movement postures and angles of the fore-and hindfeet do not change.It can be deduced that the changes in the foot reaction force are primarily caused by the movements of the limbs and body,with little relation to forefoot and hindfoot movement patterns.However,the physiological anatomy of the forefeet and hindfeet,and the demands for adhesion,may account for some invariance.

Figure 6 The comparison of the contact angle,bending angles and Fn_intial of forefoot and hindfoot.A:Schematic diagrams of the contact and bending angles of the forefoot and hindlimbs and feet of the gecko on different inclinations and motion directions (Blue lines:foreleg/foot;Orange lines:hindleg/foot;⊕ represents the COM;black dotted line indicates the body);B:Initial contact force between the forefoot and hindfoot and the substrates in five locomotion conditions (Fn_intial of level locomotion refer to (Wang et al.,2015).Significance:* represents P ≥ 0.05,** represents 0.001 ≤ P < 0.05,*** represents P < 0.001.
The overall kinematics of geckos and other animals were analysed to develop models for horizontal locomotion (Chenet al.,2006;Full and Koditschek,1999).The entire kinematic analysis for the toe pad,leg,and body is beneficial in modeling adhesion animals for uphill-downhill climbing.The forefoot and hindfoot bending anglesϑdo not change,andϑof the forefoot is always smaller than that of the hindfoot,whereas the leg contact angleβchanges with changing locomotion;when travelling on 45° slope,βof the limb above the centre of mass(COM) is smaller than that below the COM,and when moving in a vertical planeβof the limbs above the COM is larger than that below the COM (Figure 6A).This overall model shows that when dealing with environmental changes,the legs often experience more obvious changes than the toe pads which is because the realization of the adhesion function puts forward certain requirements on the adhesion devices’ movements,such as the appropriate magnitude and direction with respect to the setal shaft of the perpendicular and parallel preloads(Autumn and Peattie,2002;Russell,2002).Wanget al.proposed that the force patterns of the gecko’s forelimb and hindlimb alter when it travels on different inclinations (Wanget al.2015).We also discovered a changing threshold between the 45° and 90° slopes,and changes in the contact status of the foreleg and hindleg.The adaptive regulatory behaviours of the gecko when it travels in a variety of substrates is particularly important for adhesion climbing robots in engineering applications,as it can guide the control strategies for the robot climbing under varied circumstances.
4.3.Enlightenment for Bionic Adhesion MechanismWhether the adhesion is regulated actively or passively has long been a point of contention,and the research on this problem can be used as an inspiration for the design and control of bionic adhesion mechanism.Geckos require a specified amount of time for attachment (contact) (Autumnet al.,2006;Wanget al.,2011),and the location arrangement of the limb segments and toes are affected by the loading orientation passively(Russell and Oetelaar,2016).Our results show that the contact time percentage and bending angle of forefoot and hindfoot do not change with changes in locomotion demands (Figure 2E),while the release (detachment) time percentages change(Supplementary Materials Table 5).So it can be speculated that the contact stage depends more on the physiological structure of the adhesion mechanism,not on the locomotion environment.From our results,the contact process is likely to be passive as well;however,the release stage is probably to be adjusted actively in response to the changing locomotion requirements,which is similar to the locomotion speed of gecko that will be adjusted to adapted to different inclinations (Zaafet al.,2001).Furthermore,the changing trend and range (yaw and pitch angles) do not change obviously with the change of substrate in the swing stage of the forefoot and hindfoot,indicating that both forefoot and hindfoot have the fixed locomotion patterns in the swing process.Besides,the bending angular velocity might relate to the locomotion demands since it differs with the changing substrates (Supplementary Materials Table 10).A relationship between bending angle,angular velocity,and contact force may be one of the heels to verify the active/passive adhesion further.
The coordination mechanism between geckos’ different toes and the regulation of the adhesion force during the adhesion stage has been proposed (Songet al.,2020;Songet al.,2020).This paper has carried out a systematic investigation of the A-D cycle of geckos’ complex distributed toe pads and found that the foot locomotion postures are consistent in the swing and contact stages,with the exception of the release and adhesion stage.Nowadays,lots of adhesion robots have been devised,however,most of them do not work as effectively as real geckos(Chattopadhyay and Ghoshal,2018).To improve the robot’s climbing performance and adaptation to varied surroundings,here we proposed that a climbing robot control method held on the A-D cycle focusing active and adjustable control on the adhesion and release stages and employing an adaptable passive mode in the swing and contact stages.Therefore,in addition to the attempt of active detachment mechanism(Unveret al.,2006),a set of control methods with “active-passive control cooperation mode” for the complete A-D cycle,that is“Passive contact -Active adhesion and release -Passive swing”,and its application in bionic adhesion mechanism are worthy of further exploration.That is to say,the motion control of the robot foot during the contact and swing process may be reduced.It also implied that the best control is frequently the simplest.Additionally,the invariance of the A-D cycle across locomotion conditions remind us that the control strategy for gecko-inspired robot adhesion foot should be universal for various environments.
Both the kinematics of the A-D cycle of geckos’ adhesion devices,including the time percentage of each stage,locomotion posture,and angle changing trend (which were studied in this paper) as well as the entire dynamics of the A-D cycle (which should be tested and explored in future studies using more samples) are of great importance in understanding the A-D locomotion mechanism.This understanding will facilitate the development of the control strategy of a bionic robot with a complex adhesion mechanism.Comparative experimental tests on adhesion mechanisms of other animals,such as insects,can be conducted.Such test can help obtain more data for statistical analysis and for understanding the behavioural strategies of the A-D cycle of various adhesion devices to provide a broader reference for adhesive locomotion mechanism.
5.Conclusion
Our work investigates the A-D cycle of the gecko’s complex compliant toe pads for level and uphill-downhill locomotion on 45° and vertical surfaces.The foot bending angles (fore 41°,hind 51°),contact time percentages (fore 7.42%;hind 7.44%),and the forefoot angle ranges (yaw:163.09°;pitch:308.68°) remain constant across all experimental locomotion conditions.The foot yaw and pitch angles’ fluctuation trends remain unchanged in response to various substrates.Additionally,the forefoot locomotor angle variation ranges are larger than those of the hindfoot,and the forefoot pitch angle shifting tendency is the inverse of that of the hindfoot.Furthermore,the foot contact angle and initial contact force changed with the changing substrates,whereas the foot bending angle and contact time percentage remained the same,suggesting that the foot reaction force patterns are more related to limb/body movement than toe pad movement during the swing or contact stage,namely,the adhesion function is unaffected by the adaptive behaviours in the swing and contact stages.It indicates that the diverse and complex locomotion control of the adhesion toe pads for various environments is reduced by the adaptive behaviours in the A-D cycle,such as the constant postures in the swing and contact stages,and it advances the comprehension of the adhesive locomotion mechanism,reflects the structural and adaptative strategy of attachment devices for changing locomotion requirements,and provides insight into the design and control of adhesion robots.
AbbreviationsA-D:attachment-detachment;R:release stage;S:swing stage;C:contact stage;A:adhesion stage;TR,TS,TC,TA:the duration of four stages;COM:centre of mass;SD:standard deviation.
Availability of data and materialAll data generated or analysed during this study are included in Supplementary Materials,which can be downloaded from the website https://pan.baidu.com/s/16oRIX_WP6kNL7-MyTGCvFw?pwd=ofie.
AcknowledgementsWe thank Zhenglong QU and Xiaokang MA for participating in the experiments.We also appreciate Yi SONG,Jiwei YUAN,Donghao SHAO,and Tao SUN for reading the manuscript.This work was supported by the National Key R&D program of China (2019YFB1309600)and National Natural Science Foundation of China (51975283).
杂志排行

Asian Herpetological Research的其它文章
- A New Species of the Asian Leaf Litter Toad Genus Leptobrachella (Amphibia,Anura,Megophryidae) from Chongqing City,Southwest China
- A New Cyrtodactylus Species (Reptila:Gekkonidae) from Nan Province,NorthernThailand
- A New Species of Nidirana (Anura,Ranidae) from Southern Guangxi,China
- Spatial Patterns and Drivers of Chinese Lizard Richness among Multiple Scales
- Factors Influencing Home Ranges of the Qinghai Toad-headed Lizard(Phrynocephalus vlangalii) on the Dangjin Mountain,Gansu