路边青叶绿体基因组特征与系统进化分析
2022-06-25胡赛文马良丁怡宁毕光耀李贺敏黄岩夏至
胡赛文,马良,丁怡宁,毕光耀,李贺敏,黄岩,夏至
(1.河南农业大学农学院,河南 郑州 450046;2.河南省中药材生产技术推广中心,河南 郑州 450008)
药用植物路边青(GeumaleppicumJacq.)为蔷薇科(Rosaceae)蔷薇亚科(Rosoideae)路边青属(GeumL.),为多年生草本植物[1],是2020版中国药典收录中药材蓝布正的基源植物之一,主要分布在黑龙江、吉林、辽宁、内蒙古等地,野生资源丰富[2]。路边青为常用药中医临床,是中成药复方穿心莲片、感清糖浆[3-5],马兰感寒胶囊、蓝芷安脑胶囊[6]的重要原料药之一。具有益气健脾,补血养阴,润肺化痰等功效[7]。药理学研究表明,中药材路边青具有较好的抗炎[8]、杀菌[9]、抗肿瘤[10]等活性,其中三萜类、鞣质类、黄酮类[11-13]是其主要化学成分。目前,药用植物路边青叶绿体基因组信息和进化分析鲜见报道。
叶绿体是绿色植物进行光合作用的半自主型细胞器,编码着与光合作用等重要生物学过程相关的上百种基因[14],具有序列和结构保守,母系遗传[15-16]等优势。叶绿体基因组作为植物特有的质体基因组[17],一般为典型的双链环状4分体结构[18],对比核基因和线粒体基因,叶绿体基因组基因序列相对具有保守性,但其基因之间的间隔区位点变化幅度大,有较高的分辨率[19-22]。目前,随着基因组重测序技术发展及基因组组装和注释软件如NOVO Plasty[23]等的开发,许多植物的叶绿体基因组已报道,如匾蓄(PolygonumaviculareL.)[24]、三七(Panaxnotoginseng(Burkill)F.H.Chen ex C.Chow &W.G.Huang)[25]、五味子(Schisandrachinensis(Turca.)Bail)[26]、刺五加(Eleutherococcussenticosus(Ruprecht &Maximowicz)Maximowicz)[27]、三叶崖爬藤(TetrastigmahemsleyanumDiels et Gilg)[28]和阳春砂(AmomumvillosumLour.)[29]。本研究采用高通量测序技术,对路边青叶绿体全基因组进行测序,组装和注释,统计路边青叶绿体基因组结构特征,分析了其重复区特征,基因间隔区信息位点的变异特征,准确定位其与蔷薇科部分属的亲缘关系,为中药材路边青的资源保护及可持续利用提供一定的理论基础。
1 材料与方法
1.1 材料
路边青新鲜叶片釆集于河南省卢氏县(33°59′40″N,113°50′05″E),叶片用无菌水冲洗数次,装入取样袋后用硅胶快速干燥,带回实验室置于-80 ℃冰箱备用。样品标本号为XZ-2020-19,本研究取样的蔷薇科的物种的来源信息见表1。

表1 蔷薇亚科样品的来源Table 1 The source of samples of Rosoideae

续表 Continuing table
1.2 DNA的提取和高通量测序
DNA样品提取采用北京天根生化植物DNA提取试剂盒,高通量测序送至华大生物科技公司(北京)。试验方法和数据分析参照胡赛文等[24]的方法。
1.3 数据的拼接和注释
原始数据的组装釆用NOVO Plasty[23]程序,选取Geumrupestre(GenBank登录号:MG262388)为参考基因组进行基因注释,基因注释方法和步骤参考胡赛文等[24]的方法,最后获得路边青叶绿体基因组GenBank登录号为OM461318。
1.4 IR边界的收缩和扩张分析
利用在线工具IR scope (https://irscope.shinyapps.io/irapp/) 绘制蔷薇亚科7个属9个物种植物的叶绿体基因组IR边界对比图[30]。对比蔷薇亚科9种植物叶绿体基因组的IRA/IRB、LSC和SSC的序列特征差异。
1.5 基因间隔区位点变异分析
叶绿体基因间隔区(非编码区)DNA序列进化速率较快,在近缘物种间提供较多的系统发育信息位点[31]。本研究选取蔷薇亚科25种植物的叶绿体基因间隔区,比对分析,利用软件[32]统计这些基因间隔区信息位点变异的百分率,筛选出高变异率的基因间隔区。
1.6 系统发育分析
从NCBI数据库,选取公开发布且注释完全的25个蔷薇亚科物种的叶绿体全基因组数据(表1),以蔷薇科近缘的李亚科(Prunoideae)郁李(Cerasusjaponica)为外类群,系统发育树构建参照胡赛文等[24]的方法。
2 结果与分析
2.1 路边青叶绿体基因组的结构特征
测序原始reads为13 564 918条。经组装和注释后路边青叶绿体基因组为共价闭合的双链环状分子(图1),长155 911 bp,具有4分区域结构,其中1个LSC、1对IR和1个SSC,长度分别为85 384、26 119、18 298 bp。IR区域的GC碱基值最高,为42.7%,LSC和SSC均较低,分别为36.8%、30.7%。路边青叶绿体基因组共包括132个基因,非重复基因112个,其中86个编码蛋白基因(PCG)、8个rRNA基因与37个tRNA基因。

图1 路边青叶绿体基因组物理图谱Fig.1 Physical map of Geum aleppicum Jacq.chloroplast genome
2.2 叶绿体全基因组特征比较分析
基于25个蔷薇亚科物种的叶绿体全基因组特征(表2),进行比较分析。路边青及24种近缘植物叶绿体基因组的序列长度范围为147 835~160 007 bp,最短的叶绿体基因组是东方草莓,最长的叶绿体全基因组为棣棠花。路边青的叶绿体全基因组长度为155 911 bp,介于蔷薇亚科24个近缘种的叶绿体基因组长度范围之间。25个蔷薇亚科植物叶绿体全基因组GC含量范围为36.6%~37.6%。其中GeummacrophyllumWilld.的GC含量最低(36.6%),野草莓、东方草莓的GC含量最高(37.6%)。路边青叶绿体基因组的GC含量为36.8%,介于蔷薇亚科24个物种叶绿体基因组GC含量之间。
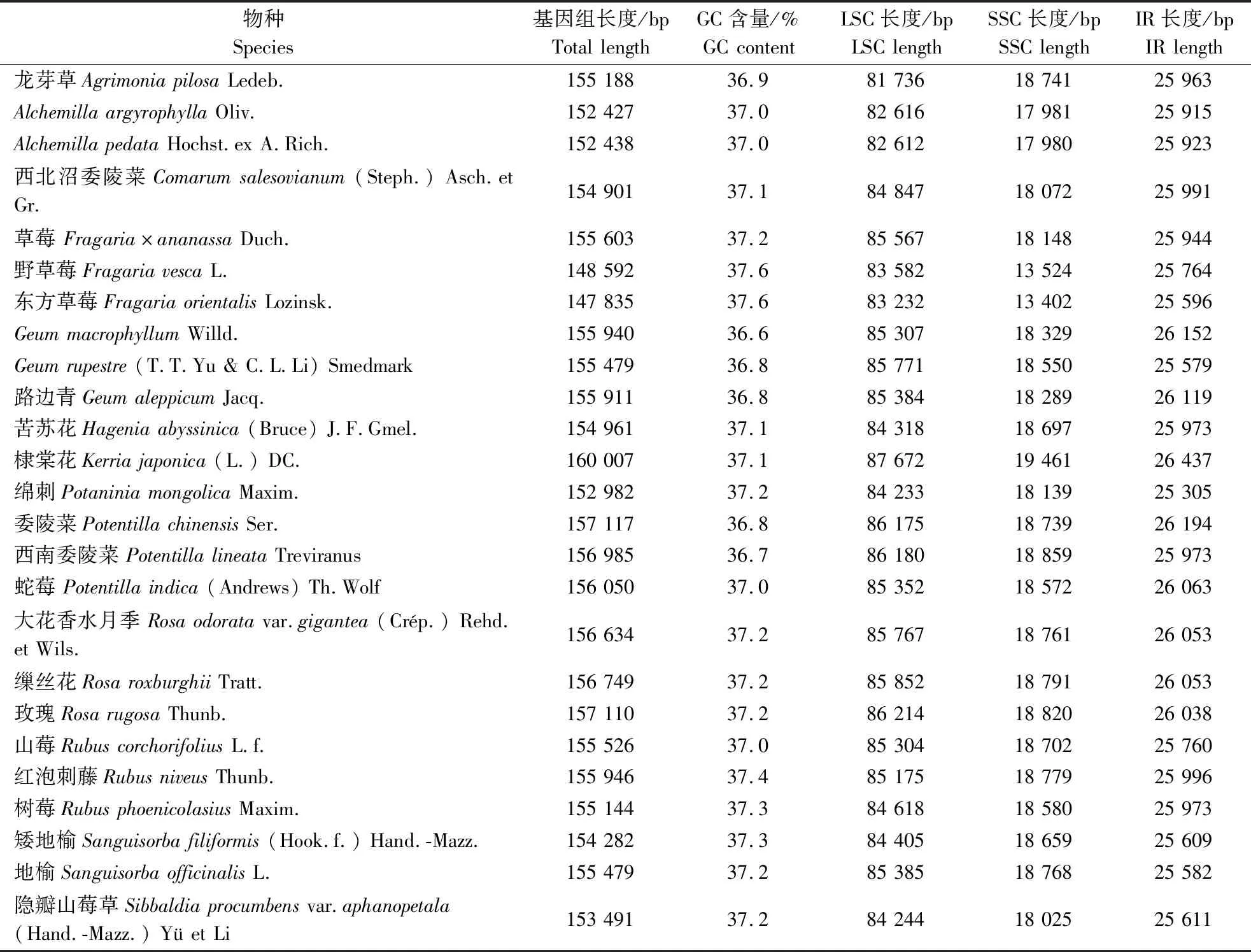
表2 25种蔷薇亚科植物叶绿体基因组的特征Table 2 Chloroplast genome characteristics in 25 species of Rosoideae
2.3 IR边界的特征
通过比较路边青和8个蔷薇亚科物种cpDNA的IR与SC边界区域基因的分布状况,发现这9个蔷薇亚科植物的叶绿体基因组长度差异较小(分布在151 144~156 986 bp之间),其序列整体上高度相似,在JLB边界,其左侧基因均为rps19(位于LSC区),右侧基因均为rpl2(位于IRb区)。其中,rps19基因距JLB边界碱基长度范围为0~22 bp,路边青为10 bp,位于9个物种的扩增范围之内。在JSB边界,地榆、树莓、玫瑰、GeummacrophyllumWilld.、路边青、草莓、龙牙草其侧翼基因完全相同,均为ycf1基因和ndhF基因(位于SSC区),但ycf1向SSC区的扩张程度不同,分别为1、11、16、10、21、11和0 bp。而西南委陵菜、Geumrupestre(T.T.Yu &C.L.Li) Smedmark的ndhF基因向IRb的扩增范围分别为36、91 bp。在JSA边界,所选的7个属9个物种均被ycf1基因横跨,他们在SSC和IRa区向ycf1的扩张程度变异范围较小,分别为4 424~4 638和1 066~1 189 bp。路边青在SSC和IRa区扩张程度分别为4 507和1 115 bp,均位于9个物种的变异范围之内。在JLA边界,其左侧基因均为rpl2(位于IRa区),右侧基因均为trnH(位于LSC区)。其中,西南委陵菜和龙牙草的trnH基因在SSC区的扩张变异程度均为3 bp,而树莓、地榆等7个物种的扩增范围为1~36 bp。路边青的trnH基因在SSC区扩张扩张变异程度为6 bp,位于9个物种的扩增范围之内。
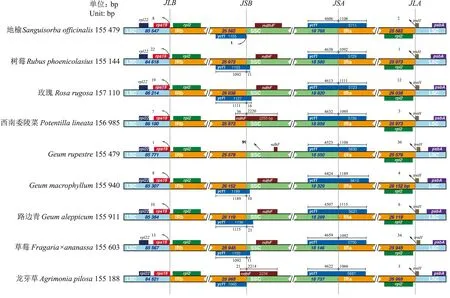
图2 路边青及8个蔷薇亚科植物的叶绿体全基因组边界图Fig.2 Border regions of chloroplast genome of Geum aleppicum Jacq.and eight Rosoideae plants chloroplast genome
2.4 叶绿体基因组基因间隔区信息位点分析
叶绿体基因组非编码区序列相对编码序列有更快的进化速率。不同进化速率的基因或序列片段适合不同分类阶元的系统进化分析[17]。本研究基于蔷薇亚科13个属25种植物的统计结果显示(表3),变异位点变化范围为3.8%~26.0%,最高的为rps4-trnT-UGU基因间隔区,其变异位点百分率为26.0%。超过20%有14个,分别为ccsA-ndhD、cemA-petA、ndhE-ndhG、petA-psbJ、petN-psbM、psbE-petL、psbM-trnD-GUC、rbcL-accD、rps4-trnT-UGU、trnE-UUC-trnT-GGU、trnF-GAA-ndhJ、trnS-GGA-rps4、trnT-UGU-trnL-UAA、ycf4-cemA。这些进化速率较高的叶绿体基因片段,可以作为下一步蔷薇科物种进化关系的理想分子标记。
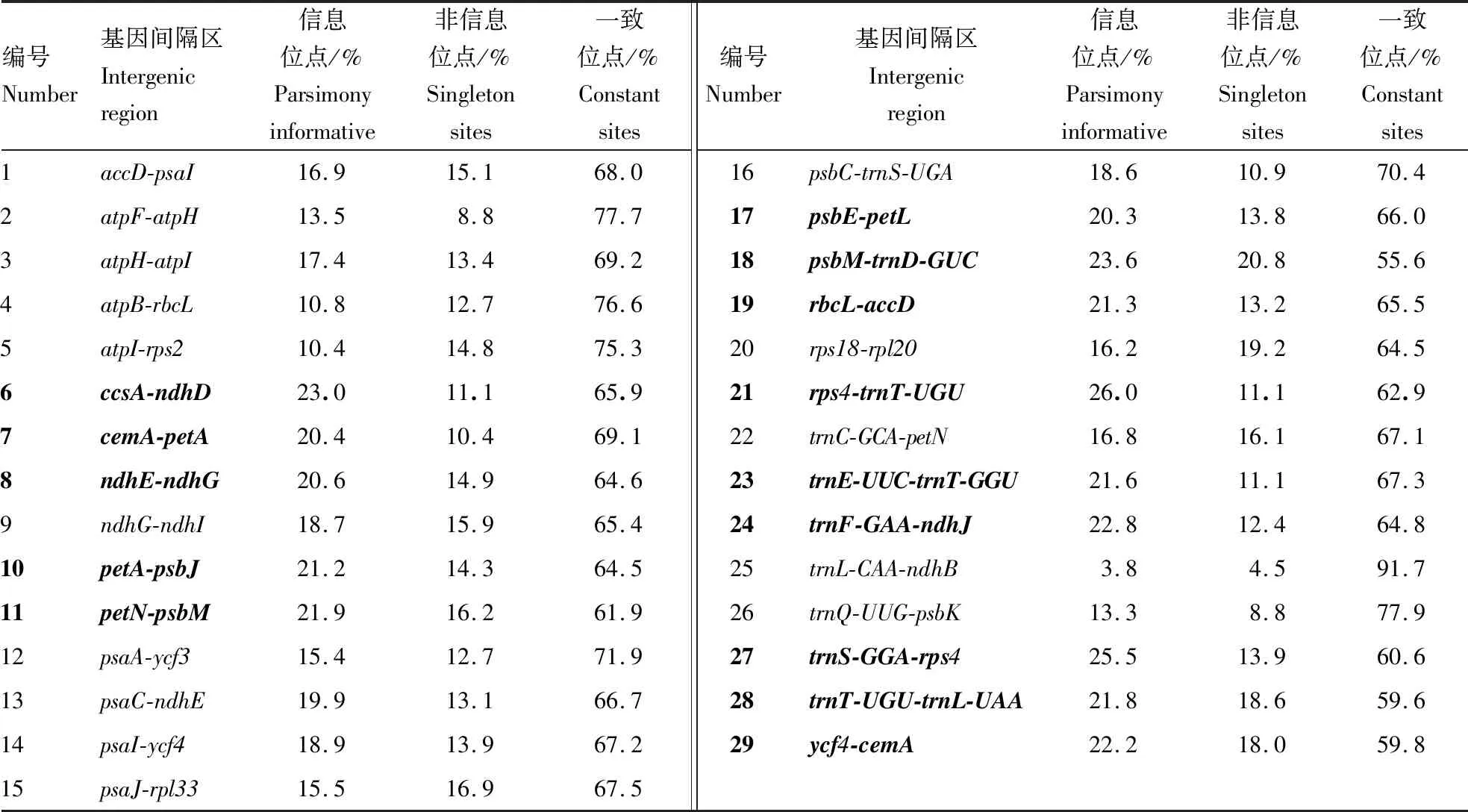
表3 取样的蔷薇亚科植物叶绿体基因间隔区变异信息Table 3 The variation information of chloroplast intergenic region in sampled Rosaceae
2.5 系统发育分析
叶绿体基因组序列进行多重比对后,系统发育树(最大似然法和贝叶斯法)(图3)结果显示,取样类群主要为两大分支,分支Ⅰ包括路边青属的GeummacrophyllumWilld.、Geumrupestre(T.T.Yu &C.L.Li) Smedmark和路边青3个种,构成单系分支;具有100%支持率。分支Ⅱ包括悬钩子属3个种以及蔷薇亚科其他10个属的植物,具有90%支持率。在分支Ⅱ内,分支A包括地榆属(SanguisorbaL.)、苦苏花属(HageniaJ.F.Gmel.)、龙牙草属(AgrimoniaL.),支持率为100%;分支B包括羽衣草属(AlchemfaL.)、沼委陵菜属(ComarumL.)、山莓草属(SibbaldiaL.)、绵刺属(PotaniniaMaxim.)、委陵菜属(PotentillaL.)、蔷薇属(RosaL.)构成单系分支,具有100%支持率。在分支B内,蔷薇属、委陵菜属、草莓属(FragariaL.)的单系性都得到100%支持率。绵刺属与草莓属聚在一支,具有100%支持率。羽衣草属、沼委陵菜属与山莓草属聚在一支,具有100%支持率。
3 结论与讨论
药用植物路边青作为常用民族药材之一,临床应用范围广。通过对比分析路边青与蔷薇亚科其他24个种的叶绿体基因组,显示路边青与蔷薇亚科其他6个属植物的叶绿体基因组各边界基因类型一致,揭示了路边青与蔷薇亚科其他物种的叶绿体基因组序列存在一定保守性。蔷薇科由于频繁的杂交、孤雌生殖和可能的快速辐射分化等因素的影响,该科的亚科、族、属分类范畴仍存在很大争议,主要分支间系统发育关系难以解析,是系统发育重建最困难的植物类群之一[33]。本研究结果表明,路边青与与蔷薇亚科其他6个属叶绿体基因组序列存在一定保守性,可以用来解决蔷薇科属间系统发育框架。

分支上部数值表示ML分析的BS分析对该分支的支持强度(>50%)。The numerical value at the top of the branch indicates the support strength of the branch by the BS analysis of the ML analysis (>50%).图3 最大似然法构建叶绿体基因组分子系统树Fig.3 Construction of chloroplast genome molecular phylogenetic tree by maximum likelibood method
本研究比较分析了蔷薇亚科24个物种的叶绿体基因组中29个共有的基因间隔区,筛选出变异位点超过20%的叶绿体间隔区序列共计有14个,分别为ccsA-ndhD、cemA-petA、ndhE-ndhG、petA-psbJ、petN-psbM、psbE-petL、psbM-trnD-GUC、rbcL-accD、rps4-trnT-UGU、trnE-UUC-trnT-GGU、trnF-GAA-ndhJ、trnS-GGA-rps4、trnT-UGU-trnL-UAA、ycf4-cemA。这些基因间隔区在蔷薇亚科的属间和种间提供了丰富的信息位点,叶绿体基因片段结合双亲遗传的核基因片段联合分析,为蔷薇亚科植物属下物种间亲缘关系构建提供有效的分子标记。
分子系统树结果表明,蔷薇亚科各属的单系都得到100%支持。路边青与同属的GeummacrophyllumWilld.和Geumrupestre(T.T.Yu &C.L.Li) Smedmark.三者构成一单系分支,具有100%的支持率,药用植物路边青隶属于蔷薇亚科的路边青属,与该属的GeummacrophyllumWilld.和Geumrupestre(T.T.Yu &C.L.Li) Smedmark.亲缘关系较近。本研究组装了蔷薇亚科药用植物路边青的叶绿体全基因组,对比分析了蔷薇亚科药用植物如委陵菜、龙牙草和地榆等叶绿体基因组特征,为蔷薇亚科药用植物的种质资源保护、遗传多样性分析和开发利用奠定理论基础。
