无患子优树种实表型及子代生长性状变异特征1)
2022-06-24李永祥邵文豪姜景民黄世清张永志
李永祥 邵文豪 姜景民 黄世清 张永志
(中国林业科学研究院亚热带林业研究所,杭州,311400)(安吉县龙山林场)
无患子(SapindusmukorossiGaertn.)是无患子科(Sapindaceae)无患子属落叶乔木树种,在我国东部、南部至西南部亚热带和热带地区呈散生或居群分布[1]。无患子喜光稍耐荫,根系深、抗性强,在低山丘陵和石灰岩山地均可栽植[2],在山地造林、坡地绿化和生态修复等方面具有重要作用[3]。无患子叶片具有吸收SO2、粉尘等空气污染物的功能,秋季叶色金黄,硕果累累,是我国南方地区重要的园林观赏乡土树种。
无患子果皮富含皂苷,种仁油脂丰富[4],是重要的化工原料和生物质能源树种[5],在有关其皂苷化学活性及提取工艺、油脂加工、药理作用与临床应用等[6-9]方面已开展了大量的研究。果实和种子表型作为无患子的主要经济性状已有相关研究报道[10],而关于其种实性状与子代生长性状之间相关性缺少研究。当前,无患子多采用种源或家系混苗造林,苗木规格差异大,生长表现差,成林效果不佳,影响了其在国土绿化和生态廊道建设中的利用。本文通过研究无患子优树无性系种实表型及子代苗期生长性状变异,探讨两者性状之间的相关性,筛选子代生长表现优良的优树无性系,以期为无患子优良种质资源在生产造林中推广应用提供技术支持。
1 材料与方法
1.1 试验地概况
研究地位于浙江省安吉县龙山林场,该地属亚热带海洋性季风气候,光照充足,气候温和,雨量充沛[11],常年平均温度17 ℃,年降水量1 400 mm。
1.2 试验材料
本研究选用的39个无患子优树无性系是从浙江省天台县无患子实生栽培群体中选优无性扩繁而来,各无性系植株已进入正常开花结实期。2018年11月份果实成熟期采摘种子,于室内阴凉通风处储藏。2019年3月份圃地育苗。播种前7 d,用水浸种,每两天换水一次。圃地苗床宽1.2 m,按照行距30 cm、株距15 cm的密度进行点播,每个优树无性系种子播种30行以上。2019年12月份对各优树无性系子代苗生长情况进行调查。
1.3 测定方法
研究采用游标卡尺测量每颗果实的纵径、横径和侧径(精度为0.01 mm);使用高精度电子台秤称量每颗果实的质量(精度为0.01 g)。果实相关数据测量完毕,剥除果皮部分,测量种子纵径、横径和侧径(精度为0.01 mm),并称量种子质量(精度为0.01 g)。
分别从每个优树无性系子代幼苗中随机选取30行,每行中随机选择一个单株,共选择30个子代,利用钢卷尺测量其苗高(精度为0.01 cm),使用电子数显游标卡尺测量其地径(精度为0.01 mm)。
家系遗传力:h2=((MSv-MSe)/MSv)×100%,式中MSv为处理间均方、MSe为处理内均方[12]。
遗传增益:△G=h2Si/X,式中h2为性状的遗传力、Si为家系选择差、X为群体平均数[13]。
2 结果与分析
2.1 无患子不同优树无性系果实性状的变异
由表1可知,经F值检验,无患子优树无性系果实质量、纵径、横径以及侧径性状在无性系间均差异极显著(P<0.01)。果实质量变异系数为8.34%~24.87%,均值为14.32%,SM9的变异系数最大(24.87%);果实质量均值为5.30 g,SM99的果实质量最大(7.98 g),SM57果实质量的最小(3.21 g),SM99和SM57的果实质量与其它无性系差异显著(P<0.05)。

图1 无患子种实表型性状示意图
果实纵径、横径、侧径变异系数范围分别为2.87%~7.44%、3.16%~11.60%、3.45%~11.37%,均值依次为4.80%、5.38%、5.75%,均明显小于果实质量变异系数均值。SM338果实纵径均值最大(24.06 mm),与其它无性系差异显著(P<0.05);SM410果实横径均值最大(28.36 mm),除SM333外,其它无性系间差异显著(P<0.05);SM99果实侧径均值最大(23.77 mm),除SM333、SM81外,与其它无性系间差异显著(P<0.05);SM67果实纵径和侧径均值最小,分别为17.69、16.62 mm,SM57果实横径均值最小,(19.18 mm),SM67和SM57与其它无性系间差异极显著(P<0.01),表明SM67、SM57无性系果实形态较小,与其它无性系相比有一定程度的分化。
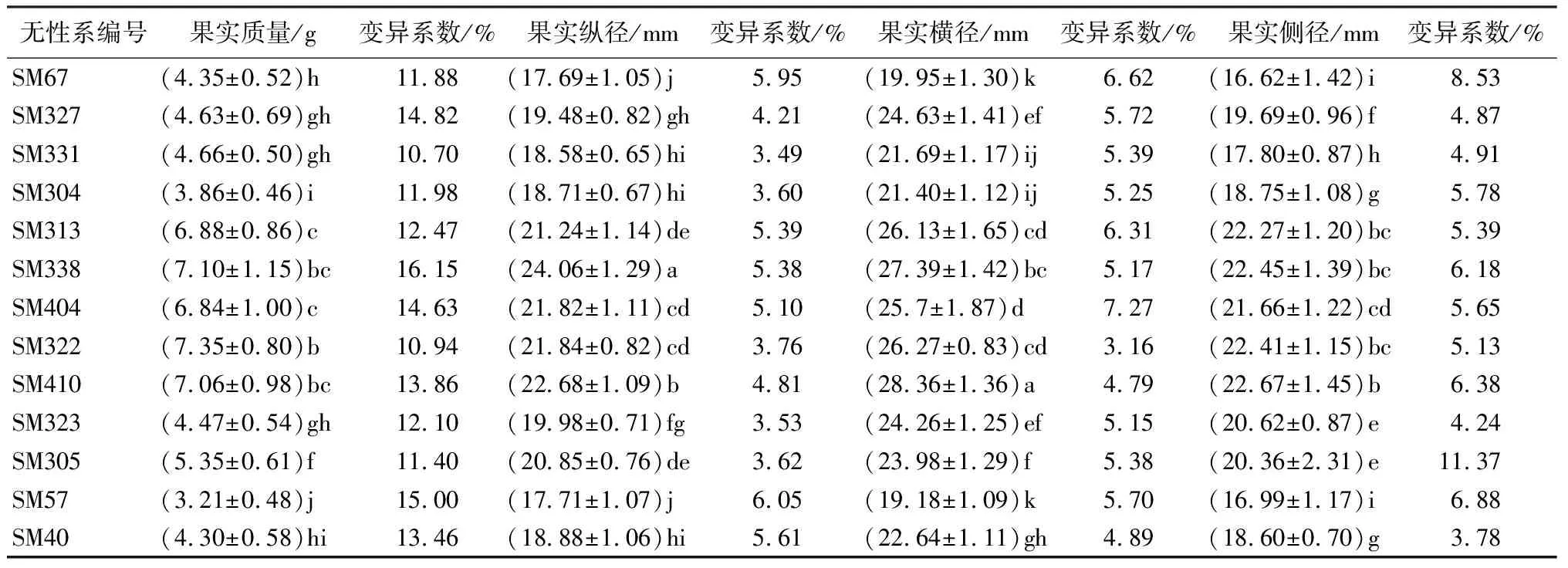
表1 无患子优树无性系果实性状
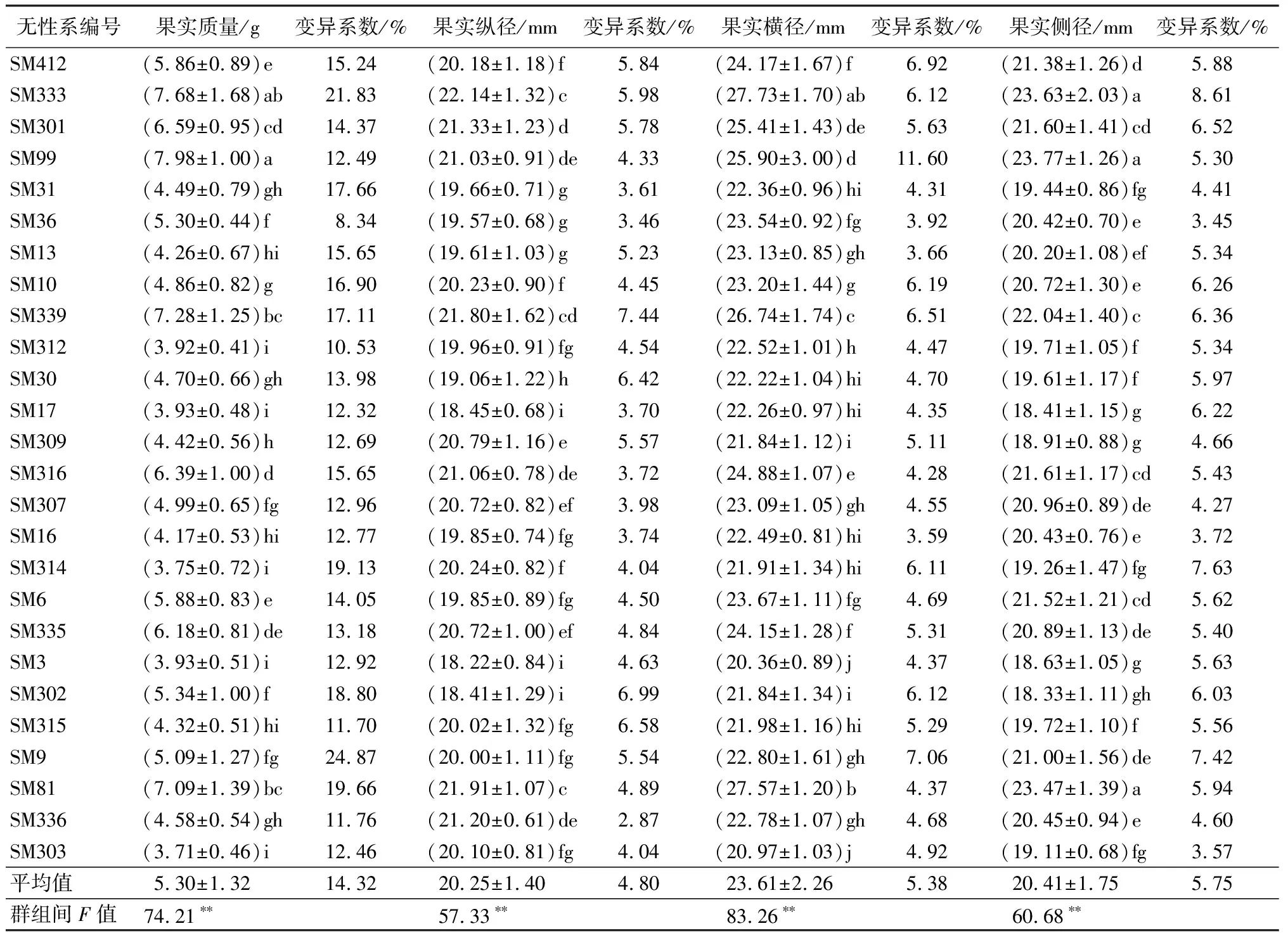
续(表1)
2.2 无患子不同优树种子性状的变异
由表2可知,经F值检验,无患子优树无性系种子质量、纵径、横径以及侧径性状在无性系间均差异极显著(P<0.01)。种子质量变异系数8.19%~20.38%,均值为11.91%,SM412的变异系数最大(20.38%)。种子质量均值为1.82 g,SM99的种子质量最大(2.53 g),除SM333外,与其它无性系间差异显著(P<0.05);SM304的种子质量最小(1.32 g),与其它无性系间差异显著(P<0.05)。种子纵径、横径及侧径的变异系数范围分别为2.36%~8.71%、2.69%~18.04%、2.81%~14.46%,均值依次为4.50%、4.87%、4.85%,均明显小于种子质量变异系数均值。SM333种子纵径及侧径均值最大,分别为16.72、15.38 mm,种子纵径均值与其它无性系间差异显著(P<0.05),SM81种子横径均值最大(17.88 mm),与其它无性系均差异显著(P<0.05),SM67种子纵径、横径及侧径均值最小,与其它无性系间均差异极显著(P<0.01),表明SM67种子形态明显较小,与其它无性系间分化明显。

表2 无患子优树无性系种子性状

续(表2)
2.3 无患子优树无性系种实性状变异的相关性
由表3可知,通过Pearson相关研究无患子优树无性系种实表型性状变异的相关性,果实质量与果实纵径、侧径性状变异相关系数分别为0.40、0.35,具有显著相关性(P<0.05);果实质量与种子质量性状变异相关系数为0.48,具有极显著相关性(P<0.01);种子纵径与果实纵径性状变异相关系数为0.50,具有极显著相关性(P<0.01),与种子横径性状变异相关系数为0.52,具有P<0.001水平的极显著性。表明无患子优树无性系果实质量性状与果实纵径、侧径性状存在显著正相关,与种子质量性状存在极显著正相关;种子纵径与种子横径、果实纵径性状存在极显著正相关。

表3 无患子优树无性系种实性状变异的相关性
2.4 无患子优树无性系子代生长性状变异
由表4可知,经F值检验,无患子优树无性系子代苗高地径性状在无性系间均差异极显著(P<0.01)。子代苗高性状变异系数为18.66%~51.87%,均值为32.72%,SM327的变异系数最大(51.87%),SM336的变异系数也相对较大(为50.92%);子代苗高均值为70.34 cm,SM322苗高均值最大(105.17 cm),除SM36、SM331、SM99外,与其它无性系间差异显著(P<0.05),SM336和SM81苗高均值分别为18.13、24.13 cm,明显小于子代苗高均值,除SM336和SM81外,与其它无性系间均差异极显著(P<0.01)。

表4 无患子优树无性系子代生长性状变异
子代地径变异系数为18.51%~50.67%,均值为30.31%,SM336的最大(50.67%),SM67的最小(18.51%);地径均值为11.15 mm,SM331子代地径均值最大(14.60 mm);SM336子代地径均值最小(4.40 mm),与SM304无性系差异显著(P<0.05),除SM304外,与其它无性系间均差异极显著(P<0.01)。表明SM336子代苗期生长较差,与其它无性系间差异显著,出现一定程度的分化。
第二,要想在高等院校“C语言程序设计”课程上机实验教学环节过程中获得良好教学效果,应以业务技能培训和外来人才招聘的方式,建构形成一支专业能力良好且综合素质优秀的“C语言程序设计”课程任课教师队伍,为高等院校各专业学生,提供“C语言程序设计”课程理论知识要素的专业化、深入化的讲解分析,提升实际教学的整体效能。
2.5 无患子优树无性系子代生长与种实性状的相关性
由表5可知,经Pearson相关分析,无患子优树无性系子代苗高与地径性状间存在极显著正相关(P<0.001),两者性状相关系数高达0.91,表明种苗较高的子代其地径也相对较大。本文所测种实表型性状,除种子质量与果实纵径两者性状间存在P<0.01水平极显著正相关,其它种实性状(果实纵径、果实横径、果实侧径、果实质量、种子纵径、种子横径、种子侧径、种子质量)间均存在P<0.001水平极显著正相关,相关系数范围0.48~0.91,果实横径与果实侧径两者性状相关系数最高为0.91。表明对其中一种实性状(如果实质量)进行高产优株选择时,可以通过其它种实性状作为预选的参考和指标。然而,无患子优树无性系各种实表型性状与子代生长性状间均无显著相关性。
2.6 无患子优树无性系子代生长性状遗传力与遗传增益
由表6可知,子代苗高与地径性状遗传力分别为96.09%和93.23%。两者性状遗传力均大于90%,表明无患子优树子代生长性状遗传力水平较高,而地径性状遗传力相对低于苗高性状。

表5 无患子优树无性系种实性状及子代生长相关性
由表7可知,子代苗高与地径性状可以良好的反应子代幼苗生长情况,分别以苗高和地径性状最大均值的前10%、20%和30%为入选率估算遗传增益。子代苗高性状以10%入选率选择时,入选无性系分别为SM322、SM36、SM331、SM99,其遗传增益最大为39.97%;入选率为20%时,入选无性系新增SM312、SM412、SM67、SM333,遗传增益为35.10%;入选率为30%时,入选无性系新增SM333、SM307、SM303、SM6,遗传增益为28.90%。子代地径性状以入选率10%选择时,入选无性系为SM331、SM322、SM312、SM323,其遗传增益也最大为27.51%;入选率为20%时,入选无性系新增SM412、SM99、SM36、SM67,遗传增益为25.59%;入选率为30%时,入选无性系新增SM13、SM6、SM16、SM303,遗传增益为23.66%。在入选率为10%、20%和30%的情况下,子代苗高、地径性状均可获得较大的遗传增益(大于20%),但遗传增益随入选率的不断增大而减小,两者性状遗传增益平均每增加10%入选率分别降低5%和2%遗传增益,入选数增加10%左右。

表6 无患子优树无性系子代生长性状方差分析

表7 不同选择强度的遗传增益
3 讨论
3.1 无患子优树无性系种实性状变异
无患子优树无性系种实质量性状变异较大,而种实纵径、横径及侧径性状变异相对较小,表明无患子种实质量性状存在较大遗传差异,种实形态(纵径、横径及侧径)性状分化较为稳定,与刘济铭等[14]研究的无患子属种质资源种实性状变异结论相似,与张毅等[15]对文冠果种实性状变异规律研究结论也相似。主要原因是种实形态性状发育较快,在一定时间内均可达到成熟时形态;而由于种实在母树的生长位置不同、养分供给分配不均、环境影响差异,导致种实内部的营养物质、水分等内含物质量和比例差异较大,使得种实质量性状变异大于其形态性状变异。果实质量与果实纵径、横径性状存在显著正相关,与种子质量性状亦存在显著正相关;种子纵径与种子横径及果实纵径性状存在极显著正相关。
3.2 子代生长变异及遗传力、遗传增益估算
无患子优树无性系子代生长性状变异大,且苗高性状变异略大于地径性状,两者变异系数均值分别为30.31%和32.72%。表明不同无性系间子代生长分化差异明显,具有显著遗传多样性,目标优良品种的选育潜力大。
遗传力是树木生长性状的主要遗传参数[16],而遗传力和遗传增益作为林木性状变异特点及子代性状受遗传控制程度的指标,可以此为依据确定良种选育方法和制定培育方案[17]。无患子优树无性系子代苗高与地径性状遗传力均高于90%,表明子代苗期生长状况主要受遗传基因控制,受环境和其它因素影响较小,与赵阳等[18]对文冠果种子及苗期生长性状地理种源变异研究结论相似。苗高性状大于其地径性状,在进行优良子代选育时,可以优先考虑子代苗高性状,以地径性状作为参照。
遗传增益的大小受遗传力和选择强度的影响,以苗高和地径性状最大均值的10%、20%和30%为入选率估算遗传增益,结果表明3种不同入选率均可获得较大的遗传增益(大于20%),这与姚淑均[19]对滇楸优树及其子代苗期性状遗传变异研究结果相似。依据不同的育种目标确定相应入选率[20],能够获得较大的遗传增益。由于子代苗高性状遗传力大于地径性状,当入选率为10%和20%时,苗高性状遗传增益均大于30%,因此,在进行无患子优良子代选育时,可以选择该两种入选率估算遗传增益作为选育指标。
3.3 种实形态与子代生长的相关性
无患子优树无性系各种实性状间均存在极显著正相关,表明种实在发育过程中,种子表型性状随果实的变化而变化,且种实形态变化较为稳定,大致呈椭球形或圆球形;因为种实属于生殖器官,主要受自身遗传因素影响,分化稳定[21]。根据各种实性状间均存在极显著正相关,对其单一性状改良,能够同时影响其它相关性状变化,在性状选择上提供不同的改良方向和选择标准[22],这与白鑫磊等[22]研究文冠果表型性状的相关性及主成分分析结论相似。在种实性状与其子代生长性状相关方面,除子代苗高与地径性状间存在极显著正相关,种实表型性状与子代生长性状间均无显著相关性,这与刘娟等[23]对圆齿野鸦椿种子性状变异及子代苗生长表现研究结论一致,与曹莉等[24]对不同家系枫杨种子特性及苗期生长相关性研究结论一致,表明子代幼苗形态一致,苗型较好,但不能通过种实性状进行早期选择或者预测子代生长;子代苗期生长优劣受遗传和环境因素影响较大,而非由种实性状差异决定。
3.4 优良子代选择
林木子代苗高和地径是反应子代幼苗生长状况的重要指标[25],大多数研究对不同种源初选是根据子代苗期生长表现[26],也有研究对优树家系进行选择时通过优树子代苗期生长表现进行[27]。无患子优树无性系种实性状与子代生长性状无显著相关性,难以通过种实表型性状进行优良子代的选择。遗传力和遗传增益可以作为子代生长性状受遗传控制程度的指标,本文无患子优树子代生长性状遗传力高,10%和20%入选率时遗传增益较大,综合考虑遗传参数与生长变异,能够更准确的选育优良子代。
通过对无患子优树无性系子代生长性状及变异大小分析,获得无患子优良子代的初步选择。并综合遗传力和遗传增益大小,SM322、SM36、SM331、SM312优树无性系子代生长表现最好,与群体具有显著差异,可以作为最优子代进行优良种质资源的保存或利用,且苗高与地径性状变异相对较小,能够稳定遗传优良性状,达到整齐、美观的造林效果;SM336、SM81、SM304优树无性系子代生长表现较差,遗传分化较大,在优良子代选择中可以将其淘汰。
4 结论
本研究中,无患子优树种实质量、种实形态、子代苗高地径等性状在无性系间差异极显著(P<0.01),存在丰富的遗传变异,种实质量性状比形态性状变异大,子代生长性状比种实表型性状大,变异来源主要在各无性系间。各种实表型性状间均存在极显著正相关(P<0.01),当对其中单一性状进行选择时,可以结合其它性状作为参照。无患子各优树无性系子代苗高与地径性状呈极显著正相关(P<0.01),而子代生长性状与种实性状间无显著相关性,一般较难通过种实性状来预测或筛选优良子代,然而可以通过子代苗期生长结合遗传参数估算,最终选择出子代生长表现优良的优树无性系SM322、SM36、SM331、SM312。
