麻点豹天牛幼虫取食和爬行的声发射信号特征1)
2022-06-24张少春熊鑫鑫朱代根邓婷婷李明
张少春 熊鑫鑫 朱代根 邓婷婷 李明
(西南林业大学,昆明,650224)(安徽工程大学)
在森林保护中,蛀干害虫幼虫钻蛀树木树干的情况日益严重,其幼虫对活立木造成的破坏几乎是不可逆的。常见的蛀干害虫有天牛类、小蠹虫类、吉丁类等[1],其中天牛类蛀干害虫危害较为严重。目前世界已知天牛种类有35 000多种,我国已有记载的达3 000多种[2]。因此,实现对天牛类蛀虫的早期辨识对于树木病虫害防治具有积极的作用。
早在20纪末,程惊秋[3]用加速传感器,对桔褐天牛幼虫和桑粒肩天牛幼虫的声信号进行了波形和频谱的分析,初步研究了天牛幼虫的声行为特征。21世纪初,韦雪青等[4]总结了昆虫声音监测史,并阐述了害虫声音检测技术的优势。Goerlitz et al.[5]通过特制电容麦克风对昆虫爬行声信号检测,研究了昆虫在不同基质上爬行发出的声音。Chesmore D et al.[6]对早期声学检测设备的改进设计,发现蛀干害虫具有独特的摄食特性,可用此特性识别昆虫的物种。许小芳等[7]在消音装置中采集蠹虫的声信号,并对其除噪,提取有效信号并对其进行频谱分析,得到几种蠹虫声活动的主频。卜宇飞等[8]利用声音侦听技术对星天牛幼虫和光肩星天牛幼虫的4种声行为的特征研究,发现天牛幼虫在取食行为下产生的声信号频率高。信号的频率和能量可以判断出害虫的活动行为。后来,Watanabe H et al.[9]又利用声发射无损检测技术证实了竹粉虱幼虫和成虫在进食阶段都会产生AE信号,并用声发射作为非破坏工具来监测这种昆虫的取食行为。豆春峰等[10]利用AE技术研究了麻点豹天牛幼虫在1 d中的活跃性,发现害虫的信号能量可以反映出其活跃程度,通过害虫的活跃时段对其进行防治。以上研究表明了声发射技术在检测树木钻蛀性害虫方面的巨大潜力,为蛀干害虫的实时动态监测提供了一种行之有效的方法。但国内利用声发射技术检测蛀干害虫的研究较少。
本文以寄生在杨树中的麻点豹天牛幼虫为研究对象,利用AE技术研究其不同生物学行为下的声学特征。主要内容为模拟天牛幼虫的蛀道,并采集天牛幼虫在蛀道中取食和爬行时所产生AE信号,然后对采集到的原始信号进行频谱分析,利用频域特征辨识幼虫的生物学行为,基于AE信号能量统计分析幼虫在不同生物学行为下的活性。对以后研究蛀干害虫提供参考。
1 材料与方法
针对麻点豹天牛对杨树破坏的问题,本研究试验样本(包括天牛幼虫及杨树段)均取自云南省昆明市盘龙区郊区金殿鸣凤山林中。首先,从带有天牛幼虫的杨树段中取出麻点豹天牛幼虫;然后将健康的杨树段样本处理成规则木段,其尺寸为220 mm×20 mm×20 mm,木材含水率为70%~80%。为保证天牛幼虫顺利钻入试验木段,用电钻在规则杨木段一端面钻深为70 mm直径为8 mm的孔,模拟幼虫在该木段的初侵入孔,使天牛幼虫容易进入,以便后面自己钻孔。选取体长32 mm,质量0.4 g,龄期为3龄的麻点豹天牛幼虫,放入已钻好孔洞的规则木段中,用小木棒堵上洞口,在试验过程中,取下小木棒,以便观察幼虫在木段中的生物学行为。为使幼虫适应环境并保持旺盛的生命力,让幼虫的生存环境更接近在活立木中高水分的环境,将其放入恒温恒湿培养箱中2~3 d;设定温度为22 ℃,湿度为90%。
本试验使用北京软岛时代科技有限公司制造的AE信号采集仪器DS5-8B。在试验木段表面布置两个AE传感器,传感器S1距离木段右端50 mm,S1与S2相距60 mm,如图1所示,试验时所用传感器型号为RS-13A,在AE传感器之后配置增益为40 dB的前置放大器,试验过程中每个通道的采样频率设置为2.5MHz。图1和2所示分别为杨树木段示意图及试验实物布置图。图2所示通过端面的模拟蛀道可以实时观测幼虫的生物学行为特征。

图1 杨树木段示意图

图2 实物布置图
2 结果与分析
2.1 幼虫取食和爬行行为的AE信号
麻点豹天牛幼虫在蛀食树干过程中会造成木纤维破坏,而纤维破坏时会以AE的方式释放应变能,同时,幼虫在爬行时也会造成木纤维的损伤,进而产生AE信号。因此,AE信号能够客观地反映幼虫的生物学行为活动。本试验主要研究麻点豹天牛幼虫的取食和爬行信号的特征,针对两种行为产生的AE信号进行频谱分析。麻点豹天牛幼虫取食和爬行的原始波形图,如图3和图4所示。

图3 取食原始波形
对比天牛幼虫取食和爬行的AE信号原始波形,爬行行为的AE信号原始波形密集。观测麻点豹天牛幼虫的爬行行为,幼虫先鼓起背部和腹部的步带,再压缩前半部的步带,然后伸长身体;采集幼虫爬行行为的原始AE信号,其波形如图4所示。对比图3中天牛幼虫取食行为的AE信号,天牛幼虫不同行为下信号幅值的最大值存在显著区别,取食行为有两次突发信号的幅值大于0.20 V,爬行行为的最大幅值为0.19 V,取食行为的最大幅值大于爬行行为的最大幅值,虽然爬行行为AE信号密集,但幅值没有显著特征。

图4 爬行原始波形
为进一步探究幼虫在两种生物学行为下的声学特性,本文统计了峰值频率、能量、撞击数等时域参数,为后面对麻点豹天牛幼虫的研究提供基础数据。由于采样率为2.5 MHz,所以原始AE信号的数据间隔可达到纳秒级,虽然时间精度非常高,但是考虑AE事件的持续时间,本文以秒为单位统计相关AE参数指标。
表1为麻点豹天牛幼虫取食时AE信号的基本参数。从表1中可以得到:幼虫取食的峰值频率中62.5%的AE信号频率是51.3 kHz,频率波动范围在(-2.4,2.5)kHz内。从信号的发生时间来看,取食行为AE信号发生的时间间隔主要(占比为75%)分布在13~24 s的范围内,并且有50%的时间间隔是不超过20 s的,最长时间间隔为41 s,平均的间隔为23 s。就能量而言,取食行为最大能量值是21.22×10-6v·s,此外取食行为的能量值都小于10×10-6v·s,其平均值为7.53×10-6v·s;方差是30。能量集中度即能量自由度[11],可以反映试件内部能量的集中度。r增加说明有大量低能量的事件产生,r减小说明有少量高能量事件产生[12-14]。能量集中度和能量成负相关关系,能量集中度的增减也反映着能量的降低与升高。结论:天牛幼虫取食行为的能量差异明显,但是频率一致。
表2是麻点豹天牛幼虫爬行时AE信号的基本参数,从表2中可以得到幼虫爬行的峰值频率中50%的AE信号频率是14.6 kHz,频率波动范围在(-1.2,0.6 kHz)内。通过发生时间可以算出AE信号发生的时间间隔,在幼虫爬行行为中,时间间隔范围在19~29 s的信号占75%,最长时间间隔为44 s,最短的时间间隔为2 s,平均的间隔为22 s。从表中能量列中可以发现,爬行行为最大能量是15.73×10-6v·s,并且在爬行行为中75%的能量小于10×10-6v·s,其平均值为8.22×10-6v·s;方差是20。

表1 取食行为AE信号基本参数
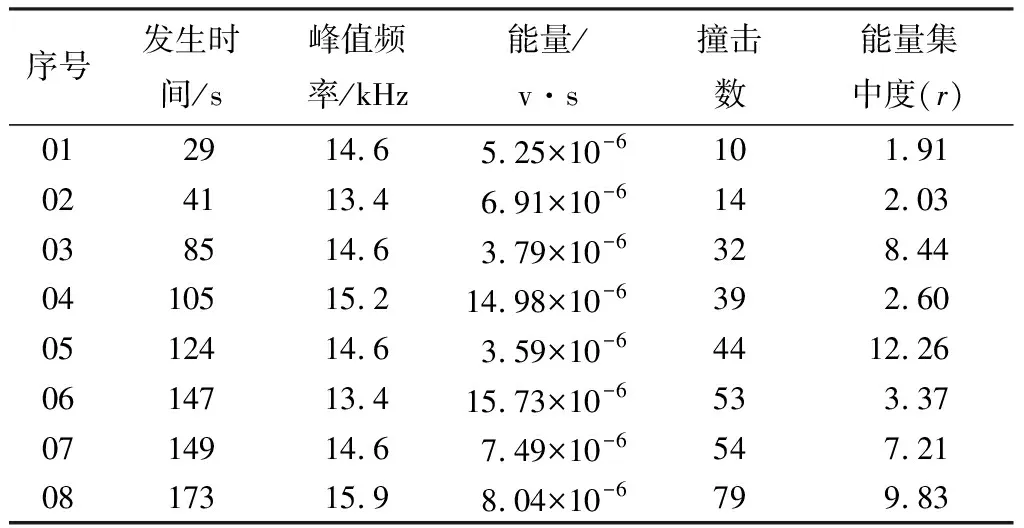
表2 爬行行为AE信号基本参数
对比表1和表2中发生时间的间隔,不论是在幼虫的取食行为还是爬行行为,幼虫行为的平均时间间隔差异不大,取食行为的时间间隔平均值是23 s,爬行行为时间间隔平均值是22 s。因此,从幼虫行为的时间间隔上无法分辨其行为特征。通过对AE信号撞击数的分析,取食行为的平均撞击数为26,爬行行为的平均撞击数为41,结合取食和爬行行为的能量集中度,反映出天牛幼虫取食和爬行行为的能量变化,但能量集中度没有明显的特征,故不能作为分辨天牛幼虫行为的依据。结论:对比取食行为的能量方差值,幼虫爬行行为的能量值变化集中,爬行行为的能量差异明显,但是频率一致。因此时间间隔和能量以及能量集中度均不适合作为辨识天牛幼虫行为的依据。可以用频率分辨天牛幼虫的行为学特征。
试验所采集的天牛幼虫取食和爬行的AE信号为低频信号,经过FFT变化分析频谱特性,为了方便观察,画出0~180 kHz的频谱图。图5为幼虫取食和爬行行为的频谱图,从图中可以看出幼虫取食和爬行行为的频谱图差异显著,图5中爬行行为对应的图形显示,频谱图峰值在15 kHz附近出现,幼虫爬行行为的频谱范围为14~50 kHz,但主频率在14.7 kHz。取食行为下的频率集中,频谱图为单峰值,取食行为的频谱图中显示其主频率为51.3 kHz。

图5 幼虫取食和爬行行为的频谱
2.2 幼虫取食和爬行的AE信号能量分析
对试验采集到的取食和爬行的原始AE信号进行能量的处理与分析,对采集到的原始信号的瞬时能量通过时域分析计算,得到对应时间内的能量值,并分别画出取食和爬行信号的能量图,如图6所示。前面已经计算了爬行行为能量方差为20,取食行为能量方差是30,爬行行为能量范围小于取食行为的能量范围。
取食行为的AE波形(图3)和能量变化(图6)客观反映了幼虫的取食行为。观察取食行为的第18秒左右,有明显的AE信号产生,图6对应的能量图中也有明显增大,该现象说明幼虫的取食是一个连续缓慢的啃食过程,随着啃食强度的增加,木材纤维断裂使应变能集中释放。在20~40 s期间,出现了高于1 V的剧烈AE信号,但对应的能量变化平缓;与之相反,在随后的40~60 s阶段,AE信号幅值降低,而对应的能量却突发增加,该现象说明对于木纤维而言,幼虫的取食过程是能量不断积累后的集中释放过程,具体而言,幼虫在取食时,不断啃咬木材纤维,最终导致木纤维断裂而释放较大应变能。

图6 天牛幼虫AE信号能量图
总而言之,通过对图3和图6的对比分析发现,幼虫在缓慢啃食时会产生大量低幅值(小于0.1 V)AE信号,随着啃食强度的增加,AE信号的幅值将发生显著变化(大于1 V),但是能量的变化通常滞后于AE信号的改变,因为能量的突变是一定时段取食行为中能量的累积而导致的木纤维断裂。为此,本文将从产生缓慢啃食AE信号到能量突变的这段时间记为幼虫的取食周期,统计发现取食周期绝大部分集中在(10,21)s区间,分别有1次达到最低的4 s和最高的24 s。
图6的爬行行为能量图中,结合原始信号,AE波形(图4)密集,但是幅值趋于稳定,能量图中出现密集的能量峰。同样地,将从幼虫爬行行为能量突变的这段时间记为幼虫的爬行周期,统计发现爬行周期绝大部分集中在(2,7)s区间,分别有1次达到最高的10 s。
对比天牛幼虫不同行为,天牛幼虫取食周期和爬行周期有显著差异,幼虫平均取食周期为14 s,平均爬行周期是5 s,因此,可以用取食周期和爬行周期来辨识幼虫的取食和爬行行为。综上所述,取食行为信号的平均能量小于爬行行为信号的平均能量,爬行行为信号能量变化频繁,但是爬行行为信号能量的最大值却低于取食行为信号能量的最大值;平均取食周期大于平均爬行周期。
3 结论与讨论
本研究是在新鲜木材中利用人工钻孔的方式模拟麻点豹天牛幼虫的蛀道,采集该幼虫取食和爬行行为的AE信号,处理试验数据,分析麻点豹天牛幼虫取食和爬行的AE信号特征。得出如下结论:可通过频谱分析得到的主频率辨别麻点豹天牛幼虫取食和爬行的行为特征;也可以通过能量变化的取食周期和爬行周期来辨别麻点豹天牛幼虫取食和爬行的行为特征;其余参数差异显著,不能有效辨别幼虫的行为学特征。麻点豹天牛幼虫取食信号的频谱峰值明显,其主频率在50 kHz附近;爬行信号的频谱图有多个峰,主频率在15 kHz附近。两种行为下信号的主频率稳定,基本不变。天牛幼虫取食行为过程是能量的积蓄。取食行为能量最大值为40.09×10-6v·s;爬行行为是幼虫一次爬行所蕴含的能量,爬行行为的能量变化较为频繁,该行为的能量最大值为33.79×10-6v·s。但是取食行为的平均能量小于爬行行为的平均能量。爬行行为的幅值比取食行为的幅值低。幼虫平均取食周期为14 s,平均爬行周期是5 s。幼虫平均取食周期大于平均爬行周期。
目前,研究人员对钻蛀性昆虫的声行为研究有限;此类昆虫的声信号普遍存在着多样性和复杂性,不同的昆虫有不同的发音机制,研究昆虫的声信号特征,可将昆虫分类并排除声音叠加的干扰,为麻点豹天牛幼虫的综合治理提供理论依据。国内对麻点豹天牛幼虫的研究很少,主要是对麻点豹天牛的营养成分[15],防治[16-18]和木材检疫方面的研究,利用声发射无损检测技术的研究仅仅是监测了麻点豹天牛幼虫在1 d中活跃的时段,为我们杀虫提供必要的条件,但并未研究麻点豹天牛的生物学行为的信号特征,而本试验是对麻点豹天牛幼虫特定行为下的AE信号研究,可进一步研究麻点豹天牛幼虫的生活习性和规律。该研究也是对麻点豹天牛幼虫更深层次的了解。
本研究在实验室条件下,只对杨树新鲜木段中采集到的麻点豹天牛幼虫的声发射信号的波形和能量进行分析,研究了天牛幼虫取食和爬行行为AE信号的特征。天牛幼虫的生物学行为有诸多不确定性,是否在取食行为或者爬行行为中伴有其他生物学行为的发生,这是在以后研究中要解决的问题,也为以后研究天牛幼虫的生物学行为提供方法,同时为直接在活立木中检测蛀干害虫提供参考。
