Sirtuin1 attenuates acute liver failure by reducing reactive oxygen species via hypoxia inducible factor 1 α
2022-06-11PanCaoQianChenChunXiaShiLuWenWangZuoJiongGong
Pan Cao,Qian Chen,Chun-Xia Shi,Lu-Wen Wang, Zuo-Jiong Gong
Abstract
Key Words: Acute liver failure; Deacetylation; Hypoxia inducible factor 1 α; Reactive oxygen species;Sirtuin1
INTRODUCTION
Acute liver failure (ALF) refers to a large number of necrosis of liver cells or severe liver damage caused by various reasons[1 ]. ALF is often accompanied by coagulation dysfunction and progressive multiple organ failure due to liver metabolism disorders and decreased immune function[2 ]. The occurrence and development of ALF is closely related to a series of inflammatory reactions, such as the release of inflammatory cytokines and the production of reactive oxygen species (ROS)[3 ].
Hypoxia-inducible factor (HIF)-1 consists of an oxygen-regulated subunit HIF-1 α and a constitutive expression subunit HIF-1 β. The activity and stability of the alpha subunit of HIF are regulated by its post-translational modifications such as acetylation[4 ]. Under hypoxic conditions, HIF-1 α acts as a primary transcription factor to regulate hypoxia-related anti-inflammatory responses[5 ]. HIF-1 α is a key factor that regulates oxygen homeostasis and redox and promotes effective adaptation to hypoxia[6 ].During the development of liver diseases such as liver cancer, hypoxia is a common finding. Hypoxia promotes the stabilization of HIF-1 α. HIF signal in innate immune cells and liver cancer cells is beneficial to the recruitment and maintenance of primordial tumorigenic immune cells and promotes immune evasion[7 ].
The monitoring of HIF-1 α activity by members of the Sirtuin (Sirt) family has been a topic of interest in recent years[8 -10 ]. According to reports, HIF-1 α has been confirmed to be related to Sirt1 , Sirt2 , and Sirt3 in the Sirt family, and the stability of HIF-1 α is related to the ROS level regulated by Sirt3 and the oxygen level regulated by Sirt6 [11 -14 ]. Sirt2 causes protein hydroxylation and ubiquitination by increasing the binding of HIF-1 α to propylamine hydroxylase[8 ]. However, the regulation mechanism of Sirt1 on HIF-1 α activity has always been a controversial topic.
Sirt1 in the sirtuin family is a nicotinamide adenine dinucleotide-dependent protein lysine deacetylase with diverse physiological functions such as anti-inflammation, neuronal signaling, DNA repair, and stress response. Sirt1 has been shown to be an important target for the treatment of various diseases[15 ,16 ], and its activation will lead to a powerful antioxidant defense system and therapeutic effects in liver ischemia reperfusion[17 ]. Studies have shown that Sirt1 regulates HIF-1 α through the formation of physical complexes between proteins, and Sirt1 may have a negative regulatory effect on HIF-1 α[18 ]. Sirt1 has also been reported to regulate HIF-1 α actively by stabilizing the protein[19 ].Whether Sirt1 is used as a negative regulator or a positive regulator of HIF-1 α or depends on the experimental conditions or experimental models remains to be further studied.
In this study, we examined the regulation of Sirt1 on HIF-1 α activity in ALF and explored its possible molecular mechanisms.
MATERIALS AND METHODS
Mice
Male C57 BL/6 J wild-type mice aged 5 -6 wk were purchased from Wuhan Biomedical Research Institute of Wuhan University. All mice were raised in the specific pathogen free animal facility of Renmin Hospital of Wuhan University with conditions of light-controlled, room temperature 25 °C, and humidity 55 ± 5 %, and they were free to eat and drink. All animal operations were approved by the Animal Care and Use Committee of Renmin Hospital of Wuhan University, China (Approval No.WDRY2021 -K016 ).
Animal models
The mice were randomly divided into six groups with 6 mice in each group: Saline control group;Hypoxia group; Lipopolysaccharide (LPS) group; Hypoxia + LPS group; Resveratrol group; and LPS +Hypoxia + Resveratrol group. Hypoxia group and Hypoxia + LPS group were cultured in COY Vinyl Anaerobic Chambers (COY, Grass Lake, MI, United States). To avoid pulmonary and cerebral edema caused by a rapid drop in oxygenation, the fraction of inspired oxygen (FiO2) (1 %/d) was gradually decreased from 21 % normoxia (room-air oxygen) to 8 % oxygen (severe hypoxia) over the course of 2 wk, followed by continual exposure to 8 % oxygen for an additional 2 wk. On the 14thd after being exposed to 8 % oxygen, Resveratrol (10 mg/kg; Sigma-Aldrich, St. Louis, MI, United States)[20 ] was given intragastrically in Resveratrol group and LPS + Hypoxia + Resveratrol group while LPS (100 μg/kg; Sigma-Aldrich) was administrated by intraperitoneal injection combined with D-Gal (400 mg/kg) in LPS group and LPS + Hypoxia + Resveratrol group[21 ]. Twenty-four hours after LPS administration, animals were quickly euthanized with inhaled CO2, followed by the collection of blood samples and liver tissues[21 ].
Cell culture
Human embryonic liver cell line L02 was purchased from China Center for Type Culture Collection(Wuhan, China). N-acetyl-L-cysteine (NAC) (Beyotime, Shanghai, China) (5 mmol/L)[22 ], nicotinamide(NAM) (Beyotime) (5 mmol/L)[23 ], GW6471 (Sigma-Aldrich) (3 μM)[24 ] or Compound C (Sigma-Aldrich) (10 μM)[25 ], which were dissolved in dimethyl sulfoxide (Sigma-Aldrich), were used to pretreat L02 cells for 1 h, followed by LPS (5 μg/mL)[26 ] treatment. Hypoxic conditions (1 % O2 ) were obtained using humidified variable aerobic workstation InVivo2400 (Ruskinn, Pencoed, United Kingdom)[27 ]. For transient transfection, cells were transfected with 2 μg plasmid of pECE-flag-Sirt1 (Addgene, Cambridge, MA, United States) and pECE empty vector (Addgene).
Biochemical analyses
Blood samples were collected after mice were anesthetized. The level of malondialdehyde (Cat. No.GM1134 ), superoxide dismutase (Cat. No. GM1133 ), and glutathione peroxidase (Cat. No. GM1135 )were determined with commercial kits (Servicebio, Wuhan, China), respectively, according to the manufacturer's instructions.
Histopathological examination
The liver tissues were sliced completely and stained with hematoxylin-eosin. The pathological changes of liver tissue were observed and evaluated by light microscope (Olympus, Tokyo, Japan). The degree of liver damage in the ALF models were assessed by the liver histology score.
Immunofluorescent staining
Liver tissue sections were intact, and L02 cell suspensions were fixed on glass slides. Sections were fixed with 4 % paraformaldehyde for 30 min, and 50 -100 μL membrane rupture working solution and 3 %hydrogen peroxide solution were added in sequence according to the manufacturer's instructions.Primary antibody against acetyl-lysine or HIF-1 α (Santa Cruz Biotechnologies, Dallas, TX, United States)diluted 1 :100 with 5 % bovine serum albumin was added on the slides and tissue sections, and the slides were incubated overnight at 4 °C in a wet box. Then, slides were incubated with secondary antibody(1 :50 dilution, Beyotime), and they were imaged using a fluorescent microscope (Olympus).
Immunoprecipitation
Approximately 1 mg of total protein was incubated with anti-Sirt1 antibody (Servicebio) or anti-HIF-1 α antibody (Servicebio) overnight at 4 °C followed by precipitation with 20 μl of protein A/G-Plus-Agarose (Servicebio) for 4 h at 4 °C. The precipitated complex was immunoblotted with anti-Sirt1 , anti-HIF-1 α, or anti-acetyl-lysine.
Detection of ROS production
L02 cell suspensions were fixed on glass slides. Cell culture fluid (2 mL) was added and the culture was continued for about 6 h. Dihydroethidium (1 mL) (Cat. No. GDP1018 ), which was dissolved in dimethyl sulfoxide at a ratio of 1 :1000 , was added to each well, and the samples were incubated in the dark. An appropriate amount of DAPI solution was added to the wells and stained. Then, a drop of antifluorescence quenching medium was added into the hole; the slides were imaged under a fluorescent microscope.
Western blotting
Proteins were extracted from cells and tissues as directed by the radioimmunoprecipitation assay kit(Sigma-Aldrich). An appropriate amount of concentrated sodium dodecyl sulfate polyacrylamide gel electrophoresis protein loading buffer was added to the collected protein samples, and then 5 -10 μL of the sample was loaded in the sodium dodecyl sulfate polyacrylamide gel electrophoresis gel sample holes. Low voltage constant pressure electrophoresis for the upper gel and high voltage constant voltage electrophoresis were applied, when bromophenol blue entered the lower gel. After electrophoresis, the proteins were transferred to polyvinylidene fluoride membranes. The following primary antibodies were used: Sirt1 (Cat. No. 9475 , Cell Signaling Technology, Danvers, MA, United States), peroxisome proliferator-activated receptor alpha (PPARα, Cat. No. 23398 R, Bioss, Woburn, MA, United States), HIF-1 α (Cat. No. 20398 R, Bioss), AMP-activated protein kinase (AMPK, Cat. No. 32047 , Abcam, Cambridge,United Kingdom), p-AMPK (Cat. No. 131357 , Abcam), Bnip3 (Cat. No. 109414 , Abcam), and glyceraldehyde-3 -phosphate dehydrogenase (Cat. No. 8245 , Abcam). Image Lab statistical software (Bio-Rad,Hercules, CA, United States) was used to evaluate band intensities on Western blots.
Statistical analyses
Statistical analysis was performed using GraphPad Prism software version 8 .0 (San Diego, CA, United States). The Y axis was labeled as fold of control mean. Data were expressed as the means ± standard deviations. Differences among multiple groups were evaluated using conventional Student’sttest or analysis of variance. Statistical significance was considered atP< 0 .05 .
RESULTS
Hypoxia aggravated ALF and increased the expression and acetylation of HIF-1 α
The liver structure of each group was shown by histopathological examination. Compared with the control group, large-scale hepatocyte necrosis in the LPS and Hypoxia groups and the number of infiltrating inflammatory cells were significantly increased, while the inflammatory response was significantly more severe in the LPS + Hypoxia group (Figure 1 A). Next, we tested the expression of some key proteins in ALF. As shown in Figure 1 B, compared with the control group, the expression of Sirt1 in the LPS group was significantly reduced, and hypoxia aggravated this effect. The expression of Bcl-2 adenovirus E1 B-interacting protein 3 (Bnip3 ) in the LPS + hypoxia group was significantly increased, as was carbonic anhydrase 9 (CA9 ), both of which are regulated by HIF-1 α, suggesting that hypoxia significantly increased the activity of HIF-1 α in the LPS group. Of note, the expression of HIF-1 α in the LPS + Hypoxia group was significantly increased in the form of acetylation. LPS significantly increased HIF-1 α acetylation induced by hypoxia (Figure 1 C).
Hypoxia reduced the expression of Sirt1 , causing the activation and acetylation of HIF-1 α
To detect changes in the expression of Sirt1 in L02 cells during hypoxia, we measured the expression levels of Sirt1 , HIF-1 α, and Bnip3 using Western blotting. Compared to the control group, hypoxia reduced Sirt1 expression and upregulated HIF-1 α and Bnip3 expression in a time-dependent manner(Figure 2 A-D). Through immunofluorescence experiments, we found that as the duration of hypoxia increased, the expression of HIF-1 α increased significantly in the form of acetylation (Figure 2 E). We then analyzed the interaction between Sirt1 and HIF-1 α. After hypoxia induced endogenous HIF-1 α,Sirt1 -HIF-1 α binding was observed (Figure 2 F). We next examined whether Sirt1 deacetylates HIF-1 α.Immunoblotting with anti-acetyl-lysine in HIF-1 α immunoprecipitates was used to detect the lysine acetylation level of HIF-1 α. As shown in Figure 2 G, Sirt1 overexpression significantly decreased HIF-1 α acetylation, suggesting that Sirt1 regulated lysyl acetylation of HIF-1 α. These results suggested that hypoxia-induced enhancement of HIF-1 α activity and lysine acetylation were related to the downregulation of Sirt1 .
The inhibition of Sirt1 induced activation of HIF-1 α and subsequently increased the production of ROS induced by hypoxia
Next, we explored the possible molecular mechanisms of the interaction between Sirt1 and HIF-1 α. As shown in the Figure 3 A, LPS increased the expression of HIF-1 α, and the expression of Sirt1 was further reduced after HIF-1 α was increased by hypoxia in L02 cells. At the same time, the use of a specific Sirt1 inhibitor NAM to inhibit Sirt1 further aggravated this effect. Sirt1 appear to interact with HIF-1 α in L02 cells. Studies have found that excessive production of ROS is considered harmful and related to hypoxia[28 ]. Oxidative stress has been shown to promote inflammation during ALF[29 ]. How oxidative stress is involved in inflammation during ALF remains unclear. Therefore, we examined the antioxidant effect of Sirt1 during hypoxia. DHE staining showed that the level of ROS stimulated by LPS was significantly increased by hypoxia, and this effect was enhanced when NAM was used to inhibit the Sirt1 signaling pathway (Figure 3 B and C). Next, we found that the expression of Sirt1 was increased and HIF-1 α was opposite when NAC was used, which is an effective ROS scavenger (Figure 3 D). At the same time, LPSinduced levels of ROS were significantly reversed by NAC (Figure 3 E and F).

Figure 1 Hypoxia aggravated acute liver failure and increased the expression of hypoxia inducible factor-1 α and its acetylation. A: The representative images of hematoxylin and eosin staining of liver in each group; B: Western blotting was performed to measure the levels of Sirtuin1 (Sirt1 ), Bcl-2 adenovirus E1 B-interacting protein 3 (Bnip3 ) and carbonic anhydrase 9 (CA9 ) in liver tissues; C: The representative images of immunofluorescence staining for Acetyl-lysine and hypoxia inducible factor (HIF)-1 α. Data shown are means ± standard deviation of three separate experiments. aP < 0 .05 vs Control group; bP < 0 .05 vs Lipopolysaccharide (LPS)-treated group; one-way analysis of variance combined with Bonferroni's post hoc test; the error bars indicate the standard deviations.GAPDH: Glyceraldehyde-3 -phosphate dehydrogenase.

Figure 2 Hypoxia decreased Sirtuin1 expression leading to the acetylation and activation of hypoxia inducible factor-1 α. A-D: Western blotting was performed to measure the levels of Sirtuin1 (Sirt1 ), Bcl-2 adenovirus E1 B-interacting protein 3 (Bnip3 ), and hypoxia inducible factor (HIF)-1 α in L02 cells;E: The representative images of immunofluorescence staining for Acetyl-lysine and HIF-1 α; F and G: Equal amounts of protein were subjected to immunoprecipitation with Sirt1 antibody or HIF-1 α antibody followed by immunoblotting with antibody against Sirt1 , HIF-1 α, or acetyl-lysine and effect of Sirt1 overexpression (O/E) was shown. Data shown are means ± standard deviations (SDs) of three separate experiments. aP < 0 .05 vs Control group; one-way analysis of variance combined with Bonferroni's post hoc test; the error bars indicate the SDs.
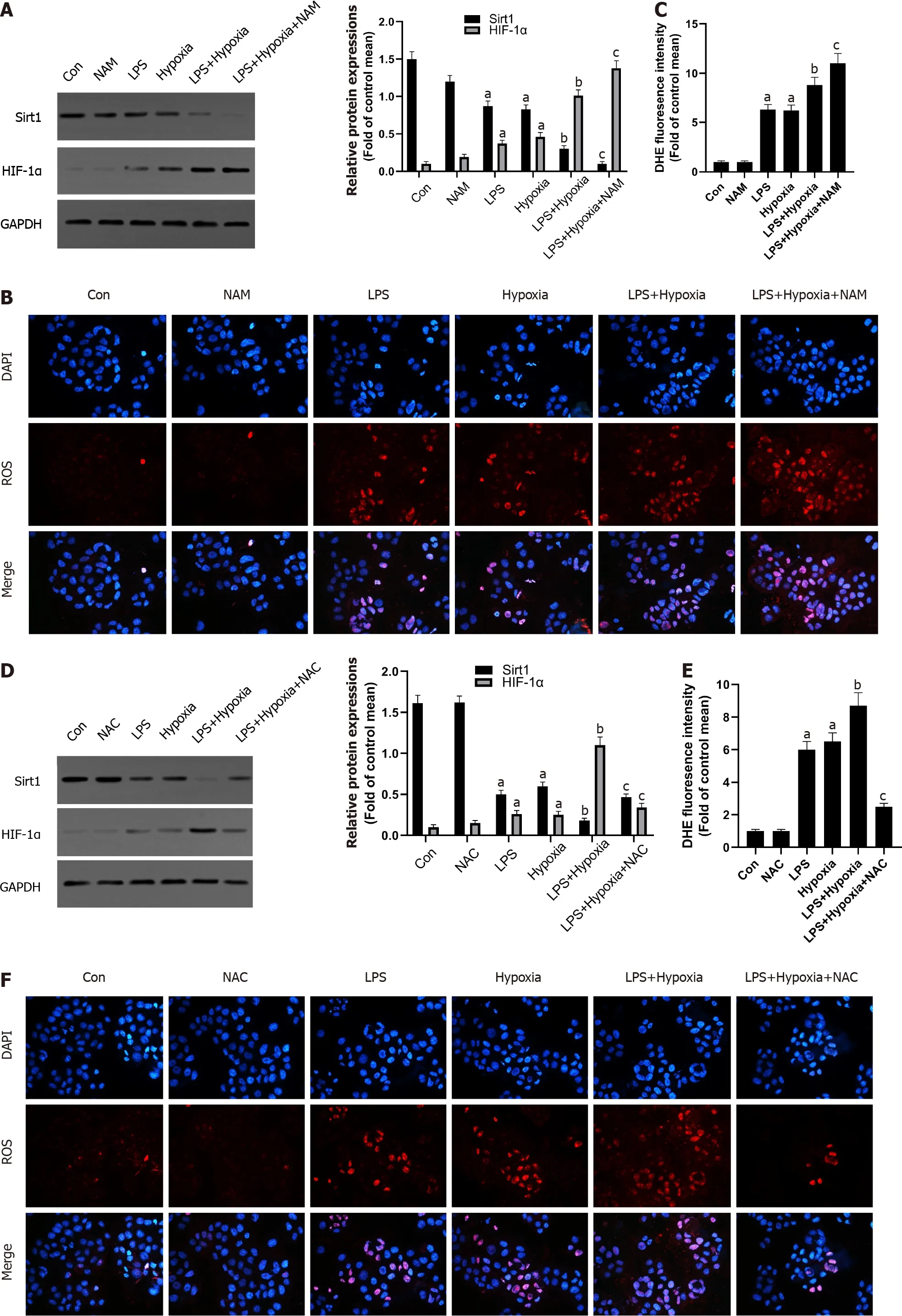
Figure 3 The inhibition of Sirtuin1 induced activation of hypoxia inducible factor-1 α and subsequently increased hypoxia-induced reactive oxygen species production. A: Western blotting was performed to measure the levels of Sirtuin1 (Sirt1 ) and hypoxia inducible factor (HIF)-1 α in L02 cells; B: Reactive oxygen species (ROS) productions were detected by dihydroethidium (DHE) staining. Representative images of the DHE staining in different groups; C: ROS productions were evaluated by quantification of mean fluorescence intensity in DHE staining; D: Western blotting was performed to measure the levels of Sirt1 and HIF-1 α in L02 cells; E and F: ROS productions were detected by DHE staining. Data shown are means ± standard deviations (SDs) of three separate experiments. aP < 0 .05 vs Control group; bP < 0 .05 vs Lipopolysaccharide (LPS)-treated group; cP < 0 .05 vs LPS + Hypoxia-treated group; one-way analysis of variance with Bonferroni's post hoc test; the error bars indicate the SDs.
The inhibition of Sirt1 /PPARα signaling pathway increased hypoxia-induced ROS production in vitro
Some studies have shown that liver PPARα expression is lower in patients with hepatitis C and advanced nonalcoholic fatty liver disease, perhaps due to the inhibitory effect of multiple cytokines[30 ].This also shows that increasing PPARα may help reduce liver inflammation. In our study, as shown in Figure 4 A, in the L02 cells stimulated by LPS, PPARα expression was decreased and aggravated after hypoxia intervention, and its effect was further aggravated when NAM was used to inhibit the Sirt1 signaling pathway, suggesting that hypoxia-induced PPARα inhibition was closely related to Sirt1 . In addition, Sirt1 expression was further reduced by the PPARα inhibitor GW6471 , while HIF-1 α was opposite (Figure 4 B) and the levels of ROS were also improved (Figure 4 C), suggesting that the inhibition of Sirt1 /PPARα signaling pathway might increase hypoxia-induced ROS production in L02 cells.
The inhibition of Sirt1 /AMPK signaling pathway increased hypoxia-induced ROS production in vitro
AMPK acts as a regulator of cellular energy metabolism and redox homeostasis. More and more evidence shows that AMPK plays a protective role by regulating the redox system[31 ]. Next, we further studied whether Sirt1 can regulate AMPK and its role in cell hypoxia in L02 cells. As shown in Figure 5 A, the phosphorylation level of AMPK in L02 cells induced by LPS after hypoxia treatment was significantly reduced, while NAM pretreatment aggravated this effect, indicating AMPK could be modulated by hypoxiaviaSirt1 . In addition, AMPK inhibitor Compound C further reduced the expression of Sirt1 , the expression of HIF-1 α was further increased (Figure 5 B), and the levels of ROS were also improved (Figure 5 C). Therefore, these results suggested that Sirt1 /AMPK signaling pathway might be involved in modulating ROS in LPS-stimulated L02 cells during hypoxia.
The activation of Sirt1 induced the inactivation and deacetylation of HIF-1 α and subsequently rescued the progressive aggravation of ALF induced by hypoxia in vivo
Finally, to determine further whether Sirt1 attenuated the progressive aggravation of ALF induced by hypoxia through the Sirt1 /AMPK or the Sirt1 /PPARα pathway, LPS-stimulated mice were exposed to hypoxia with or without resveratrol treatment, which is a Sirt1 activator. Compared with the LPS group,activation of Sirt1 by resveratrol alleviated the more sever liver tissue damage in the LPS + Hypoxia group (Figure 6 A and B). LPS + Hypoxia group mice showed lower activity of superoxide dismutase and glutathione peroxidase, while malondialdehyde levels were increased, indicating that hypoxia led to decreased antioxidant activity. However, resveratrol treatment could significantly improve the activity (Figure 6 C). As shown in Figure 6 D, resveratrol dramatically alleviated hypoxia-induced reduction levels of PPARα protein and the phosphorylation of AMPK in LPS-stimulated mice,suggesting that Sirt1 was a key regulator on the activation of PPARα and the phosphorylation of AMPK during hypoxia in ALF. Finally, we demonstrated with animals whether Sirt1 has a regulatory effect on hypoxia-induced HIF-1 α lysine acetylation and HIF-1 α activity. As shown in Figure 6 E, with the intervention of resveratrol, the expression of HIF-1 α and the level of acetylation decreased significantly.These findings indicate that the activation of Sirt1 induced HIF-1 α inactivation and deacetylation,thereby alleviating the progressive aggravation of ALF induced by hypoxia.
DISCUSSION
Recently, more and more studies have confirmed the effect of Sirt1 in liver disease. Sirt1 has been confirmed to have a protective effect in a variety of disease models, including liver fibrosis[32 ], druginduced liver injury[33 ], non-alcoholic fatty liver disease[34 ], and fatty liver[35 ]. As well known, HIF-1 α is a transcription factor that can promote the adaptive response of cells to hypoxia. Some reports have mentioned the connection between Sirt1 and HIF protein, but there are still many controversies about the results. According to reports, in hypoxic Hep3 B or HEK293 cells, Sirt1 targeted HIF-2 α and increased the transcriptional activity of HIF-2 α but not HIF-1 α[36 ]. On the contrary, another group of studies showed that Sirt1 interacted with HIF-1 α, causing HIF-1 α deacetylation to promote its activity in Hep3 B and Huh7 cells[19 ]. Therefore, the regulation of Sirt1 on the activity of HIF-1 α and its expression seems to be cell-type-specific, which is currently unclear. It has not been reported that the beneficial effect of Sirt1 activation is related to its HIF-1 α deacetylation against ALF.
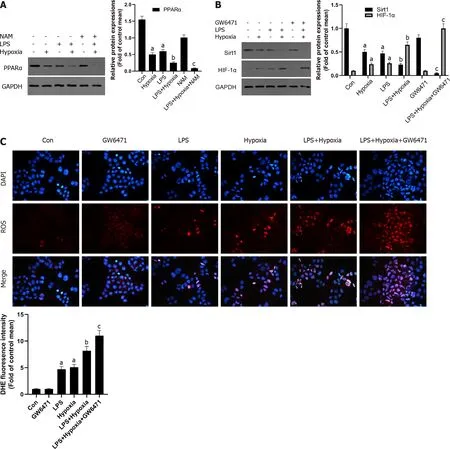
Figure 4 The inhibition of Sirtuin1 /peroxisome proliferator-activated receptor alpha signaling pathway increased hypoxia-induced reactive oxygen species production. A: Western blotting was performed to measure the levels of peroxisome proliferator-activated receptor alpha (PPARα) in L02 cells; B: The levels of Sirtuin1 (Sirt1 ) and hypoxia inducible factor (HIF)-1 α in L02 cells; C: Reactive oxygen species productions were detected by dihydroethidium (DHE) staining and evaluated by quantification of mean fluorescence intensity in DHE staining. Data shown are means ± standard deviations (SDs)of three separate experiments. aP < 0 .05 vs Control group; bP < 0 .05 vs Lipopolysaccharide (LPS)-treated group; cP < 0 .05 vs LPS + Hypoxia-treated group; one-way analysis of variance with Bonferroni's post hoc test; the error bars indicate the SDs.
In our research, we found that the activity of HIF-1 α increased after acetylation and promoted hepatocyte apoptosis in ALF models and hypoxia modelsin vitro. In addition, we demonstrated that the expression of Sirt1 in L02 cells decreased in a time-dependent manner due to hypoxia, which was closely related to the activation and acetylation of HIF-1 α. During hypoxia, with the decrease of the level of nicotinamide adenine dinucleotide, the activity of Sirt1 decreased and HIF-1 α transcription activity further increased[18 ,19 ]. Therefore, the insufficient expression of Sirt1 in the liver or the acetylation of HIF-1 α might be the key mediators of ALF.

Figure 5 The inhibition of Sirtuin1 /AMP-activated protein kinase signaling pathway increased hypoxia-induced reactive oxygen species production. A: Western blotting was performed to measure the levels of AMP-activated protein kinase (AMPK) and p-AMPK in L02 cells; B: The levels of Sirtuin1 (Sirt1 ) and hypoxia inducible factor (HIF)-1 α in L02 cells; C: Reactive oxygen species (ROS) productions were detected by dihydroethidium (DHE) staining and evaluated by quantification of mean fluorescence intensity in DHE staining. Data shown are means ± standard deviations (SDs) of three separate experiments. aP <0 .05 vs Control group; bP < 0 .05 vs Lipopolysaccharide (LPS)-treated group; cP < 0 .05 vs LPS + Hypoxia-treated group; one-way analysis of variance combined with Bonferroni's post hoc test; the error bars indicate the SDs.
Next, we carefully evaluated Sirt1 's regulatory effect on HIF-1 α activity in ALF and explored its possible molecular mechanisms. ROS are by-products of normal metabolism in living cells, but excessive ROS accumulation can damage organelles, leading to increased oxidative stress[37 ,38 ]. ALF produces excessive amounts of ROS due to insufficient detoxification of toxic substances in the liver[39 ]. Sirt1 has been reported to play an important role in anti-inflammatory and antioxidant processes[40 ]. Here, we demonstrated that HIF-1 α was over-activated in hypoxia due to increased level of ROS in the absence of Sirt1 , and the effect was inhibited by the antioxidant NAC, indicating that ROS was involved in this activation.
In particular, PPARα is reported to be a potent inhibitor of NF-κB signaling pathway and inflammation[41 ]. The positive effect of Sirt1 on the inflammatory pathway may be related to PPARα[42 ], and the interference of PPAR transcriptional activity may disturb estrogen/androgen receptor expression and impair steroidogenesis and ROS metabolism[43 ]. In addition, PPARα contributes to the protection of redox homeostasis[44 ]. Previous studies have confirmed that Sirt1 can regulate AMPK, which is an important energy sensor[45 ]. AMPK acts as a regulator of cellular energy metabolism and redox homeostasis. More and more evidence shows that AMPK plays a cardiovascular protective role by regulating the redox system[46 ]. In diabetes, the activation of AMPK increases the expression of mitochondrial antioxidant enzymes and leads to a decrease in the production of mitochondrial ROS in endothelial cell[47 ].


Figure 6 The activation of Sirtuin1 induced the deacetylation and inactivation of hypoxia inducible factor-1 α, and subsequently rescued the progressive aggravation of acute liver failure induced by hypoxia. A: Mice were pretreated with resveratrol or exposed to hypoxia and then stimulated with lipopolysaccharide (LPS). The representative images of hematoxylin and eosin staining of liver in each group; B: The liver histological score of liver in each group; C: The levels of malondialdehyde (MDA), superoxide dismutase (SOD), and glutathione peroxidase (GSH-Px) of mice in each group; D: Western blotting was performed to measure the levels of Sirtuin1 (Sirt1 ), hypoxia inducible factor (HIF)-1 α, peroxisome proliferator-activated receptor alpha (PPARα) and p-AMPactivated protein kinase (AMPK) in liver tissues and the protein expression were quantified; E: The representative images of immunofluorescence staining for Acetyllysine and HIF-1 α. Data shown are means ± standard deviations (SDs) of three separate experiments. aP < 0 .05 vs Control group; bP < 0 .05 vs LPS-treated group; cP< 0 .05 vs LPS + Hypoxia-treated group; one-way analysis of variance combined with Bonferroni's post hoc test; the error bars indicate the SDs.
Our experiments revealed that the inhibition of PPARα and the phosphorylation of AMPK induced by hypoxia were closely related to Sirt1 , and the inhibition of Sirt1 /PPARα or Sirt1 /AMPK signaling pathway might increase hypoxia-induced ROS production in L02 cells. In order to determine whether the activation of Sirt1 induced inactivation of HIF-1 α, the progressive aggravation of ALF induced by hypoxiain vivowas rescued in mice treated with resveratrol. As expected, the activation of Sirt1 significantly alleviated the degree of liver damage in ALF and enhanced antioxidant activity.Resveratrol dramatically alleviated the hypoxia-induced reduction level of PPARα protein and the phosphorylation of AMPK in LPS-stimulated mice. In addition, the activation of Sirt1 induced the deacetylation of HIF-1 α compared to LPS-stimulated mice exposed to hypoxia; the expression of HIF-1 α and the level of acetylation decreased significantly.
One limitation of our study is that we did not use HIF-1 α overexpressing micein vivoto test whether the increase of Sirt1 activity can rescue ALF. We need to conduct further experiments to solve this problem.
CONCLUSION
In summary, we have demonstrated that Sirt1 reduced oxidative stress in ALF by regulating the activity and acetylation of HIF-1 α, achieved by normalizing the Sirt1 /PPARα and Sirt1 /AMPK pathway. Our research showed that the deacetylation and inactivation of HIF-1 α induced by the activation of Sirt1 might have therapeutic benefits in reducing liver damage during ALF.
ARTICLE HIGHLIGHTS

Research motivation
The results of this study might provide a basis for the application of Sirt1 in the treatment of ALF and further understanding of the mechanism of Sirt1 and HIF-1 α in the process of ALF.
Research objectives
This study detected the changes in the expression of Sirt1 and HIF-1 α in liver tissues and hepatocytes under hypoxia during the ALF process as well as the differences in the expression levels of key enzymes. In addition, this study further explored the relationship and mechanism of Sirt1 signaling pathway and HIF-1 α expression.
Research methods
Western blotting was used to detect the expression levels of Sirt1 and HIF-1 α related proteins in mouse liver tissues, and immunofluorescence staining was used to observe the acetylation level of HIF-1 α.Detection of HIF-1 α and reactive oxygen species (ROS) levels and the correlation analysis between Sirt1 and HIF-1 α were performed. Finally, Sirt1 was activated to observe the influence of the Sirt1 signaling pathway and HIF-1 α on ALF, and changes in the expression levels of related markers were detected.
Research results
The expression of Sirt1 decreased and the level of HIF-1 α acetylation increased in hypoxic mice, and the levels of carbonic anhydrase 9 and Bcl-2 -adenovirus E1 B interacting protein 3 increased significantly,which was regulated by HIF-1 α, indicating an increase of HIF-1 α activity. Under hypoxia, the downregulation of Sirt1 activated and acetylated HIF-1 α in L02 cells. The inhibition of Sirt1 significantly aggravated this effect and the massive production of ROS. The regulation of ROS was partly through peroxisome proliferator-activated receptor alpha or AMP-activated protein kinase (AMPK). The activation of Sirt1 effectively relieved ALF aggravated by hypoxia, the production of ROS, and cell apoptosis. It also induced the deacetylation of HIF-1 α and inhibited the activity of HIF-1 α.
Research conclusions
The inhibition of peroxisome proliferator-activated receptor alpha and the phosphorylation of AMPK induced by hypoxia were closely related to Sirt1 , and the inhibition of Sirt1 /PPARα or Sirt1 /AMPK signaling pathway might increase hypoxia-induced ROS production. The activation of Sirt1 reduced oxidative stress in ALF by regulating the activity and acetylation of HIF-1 α.
Research perspectives
The results of this study showed that the deacetylation and inactivation of HIF-1 α induced by the activation of Sirt1 might have therapeutic benefits in reducing liver damage during ALF. This study preliminarily clarified the role of Sirt1 and HIF-1 α in ALF, so as to deepen the understanding of the mechanism of ALF, and provided guidance for the selection of ALF treatment targets. The results of this study indicate that Sirt1 activator may have a certain prospective application as a therapeutic drug for ALF.
ACKNOWLEDGEMENTS
The authors would like to thank the central laboratory at Renmin Hospital of Wuhan University(Wuhan, Hubei Province, China) for their support of our study.
FOOTNOTES
Author contributions:Cao P and Gong ZJ designed the study; Cao P, Chen Q and Shi CX performed most of the experiments and wrote the article; Wang LW analyzed the data; All authors approved the final version of the article.
Supported byNational Natural Science Foundation of China, No. 82070609 .
Institutional review board statement:This study was approved by the Institutional Review Board of Renmin Hospital of Wuhan University.
Institutional animal care and use committee statement:All animal operations were reviewed and approved by the Institutional Animal Care and Use Committee of Renmin Hospital of Wuhan University (Approval No. WDRY2021 -K016 ), and were conducted in accordance with the established International Guiding Principles for Animal Research.
Conflict-of-interest statement:The authors declare no conflict of interest.
Data sharing statement:No additional data are available.
ARRIVE guidelines statement:The authors have read the ARRIVE Guidelines, and the manuscript was prepared and revised according to the ARRIVE Guidelines.
Open-Access:This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution NonCommercial (CC BYNC 4 .0 ) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is noncommercial. See: https://creativecommons.org/Licenses/by-nc/4 .0 /
Country/Territory of origin:China
ORCID number:Pan Cao 0000 -0001 -6216 -6901 ; Qian Chen 0000 -0002 -2411 -2838 ; Chun-Xia Shi 0000 -0001 -8977 -2784 ; Lu-Wen Wang 0000 -0002 -0654 -1196 ; Zuo-Jiong Gong 0000 -0002 -4676 -5856 .
Corresponding Author's Membership in Professional Societies:Renmin Hospital of Wuhan University, Renmin Hospital of Wuhan University.
S-Editor:Wu YXJ
L-Editor:Filipodia
P-Editor:Yuan YY
杂志排行

World Journal of Gastroenterology的其它文章
- Current status and future of targeted peptide receptor radionuclide positron emission tomography imaging and therapy of gastroenteropancreatic-neuroendocrine tumors
- Biliary metal stents should be placed near the hilar duct in distal malignant biliary stricture patients
- Clinical outcomes of endoscopic papillectomy of ampullary adenoma: A multi-center study
- New insights in diagnosis and treatment of gastroenteropancreatic neuroendocrine neoplasms
- Gut homeostasis, injury, and healing: New therapeutic targets
- Peroxisome proliferator-activated receptor-alpha activation and dipeptidyl peptidase-4 inhibition target dysbiosis to treat fatty liver in obese mice