入侵植物少花蒺藜草异型种子萌发策略及其幼苗生长特性
2022-06-11曲婷周立业
曲婷,周立业
(内蒙古民族大学农学院,内蒙古 通辽 028000)
科尔沁沙地是我国面积最大、人口密度最高、交通干道最集中的沙地[1]。禾本科蒺藜属一年生草本植物少花蒺藜草(Cenchrus incertus),原产于北美洲热带沿海地区,20 世纪40年代传入我国,根据调查资料,少花蒺藜草从1983年在科尔沁沙地首次被记录以来,由点向面扩散,侵占了许多草场、农田,目前已成为科尔沁沙地危害最为严重的入侵杂草之一[2−3]。少花蒺藜草的入侵已严重影响了草地资源的正常利用及群落组成[4−5],使本来脆弱的生态系统进一步恶化,造成了不可逆转的破坏,给当地农牧民日常生活和畜牧业生产造成了极大损失[6−7]。
少花蒺藜草靠种子进行繁殖,在适宜环境下种子可随时萌发、扩散进而维持种群的建立和延续[8],种子表现为异型性(较大的为M 型种子,较小的为P 型种子)。种子异型性是植物在长期进化过程中应对干扰、胁迫等外界条件的一种主动适应[9],是许多旱区一年生植物产生的在形态结构和萌发特性等方面存在差异的现象[10],种子异型形态特征表现为种子大小、形状、颜色、种皮纹理和二级结构等存在差异[11−13]。而种子异型行为是生态行为,主要体现在种子传播能力、种子萌发与休眠特性、种子库持久能力及幼苗生长特性等方面[14−16],具有种子异型性的植物可在生长过程中降低单次萌发的风险,提高幼苗存活率,从而确保种群成功建立和顺利繁殖[17]。
前期研究发现,野外条件下少花蒺藜草通常以M 型种子萌发为主,P 型种子采取休眠策略;早期对少花蒺藜草M 型种子萌发所形成的植株进行去除,P 型种子会被激发补充种群数量,晚期去除,刺苞内的P 型种子保持休眠状态以待来年在适宜条件下萌发,P 型种子会根据其所处环境不同从而采取萌发或休眠的生存策略对少花蒺藜草成功入侵以及完成生活史起了关键作用[18]。本研究以少花蒺藜草刺苞及异型种子为对象,通过测定M 型和P 型种子的萌发特征及其形成幼苗的形态特征和生物量特征,分析少花蒺藜草异型种子的萌发策略以及在科尔沁沙地的适应和入侵机制。
1 材料与方法
1.1 研究区概况与供试材料
少花蒺藜草刺苞取于通辽市辽河镇四合屯(43°40′N,122°04′E),该地属于温带半干旱季风性气候,海拔187 m,年平均气温为5.6 ℃,平均日照3000 h 左右,≥10 ℃年积温3000~3200 ℃,无霜期147 d,年平均降水量为350~400 mm,降水大多集中在6−8月。少花蒺藜草刺苞及异型种子见图1。

图1 少花蒺藜草刺苞(A)和刺苞内异型种子(B)Fig.1 Bract(A)and seeds in the bract(B)of C. incertus
1.2 试验方法
1.2.1 模拟刺苞、两种异型种子及刺苞的萌发试验 试验所用刺苞取于2019年10月,为保证种子正常萌发,试验开始于2020年11月。将同一刺苞内的两种异型种子取出,用纱布进行包裹模拟刺苞果皮并用针线将纱布缝合,记为模拟刺苞处理(图2)。萌发试验采用水培法,将模拟刺苞、M 型种子、P 型种子及刺苞各30 粒置于培养皿中,以滤纸为萌发床,加入适量蒸馏水,培养皿加盖放置在(25±1)℃的培养箱里,经常变换培养皿在培养箱中的位置,确保所有种子所处的环境均一,每处理6 个重复。每日记录各处理发芽情况(种子萌发以胚根长度为种子或刺苞长度、胚芽露出种子或刺苞长度的1/2 为标准),并用直尺测定刺苞及两种异型种子的胚根、胚芽长,测定时间从种子发芽开始,并计算生长速率。
图2 少花蒺藜草模拟刺苞的发芽试验Fig. 2 Experiment of simulating the germination of bract ofC. incertus
1.2.2 刺苞及异型种子形成幼苗的形态及生物量特征的测定 将少花蒺藜草刺苞、M 型和P 型种子均匀一致地播种于相同规格的花盆(花盆上口直径20 cm,高12 cm)中,栽培基质为沙土和蛭石(体积比=1∶1),播种前浇透水,在每个花盆内种植50 粒刺苞(种子),刺苞覆土0.5 cm,异型种子覆土0.3 cm,两种异型种子各设置5 个重复,刺苞设置10 个重复(前5 个重复测定刺苞内M 型种子形成幼苗的形态及生物量特征,后5 个重复待刺苞内M 型种子出苗后,挖出幼苗,在盆中清洗干净,去除已萌发M 型幼苗的地上及地下部分,将留存有P 型种子的刺苞再次进行种植,将此处理记为刺苞内P 型种子萌发的幼苗),当以上4 种处理的幼苗株高达到10 cm 左右时测定生长指标。
1.3 测定指标与方法
1.3.1 生长指标的测定 1)发芽率=最终发芽种子数/供试种子总数×100%,24 小时内无新增发芽种子视为种子发芽终止[19]。
2)发芽势:按照发芽过程中日发芽种子数达到最高峰(本试验为发芽后第3 天)发芽的种子数占供测样品种子数的百分率计算[19]。
3)发芽指数(germination index,GI):GI=∑(Gt/Dt),式中:Dt 为萌发天数,Gt 为Dt 相对应的每天发芽种子数[20]。
4)活力指数(vigor index,VI):VI=S×GI,式中:S 为平均胚根长,GI 为发芽指数[20]。
5)胚根(胚芽)生长速率=胚根(胚芽)的长度/生长所需时间。
1.3.2 幼苗叶面积的测定 当少花蒺藜草株高达到10 cm 时,选择健康且长势一致的植株,用YMJ−CH 型智能叶面积扫描仪(杭州市余杭区)测量第2、3 片长势接近的功能叶叶面积、叶片长、叶片宽,每个花盆内随机选取10 株植物,进行3 次重复,取平均值。
1.3.3 幼苗生物量的测定 当株高达到10 cm 时,浇水后连根挖出幼苗,用自来水洗净泥土后用滤纸吸干植株表面水珠,选取30 株生长一致的植株,将每个植株的根、茎、叶分别分离,将冲洗干净的根、茎和叶装入牛皮纸袋,在105 ℃下杀青15 min,75 ℃烘干至恒重,用电子天平称量各部分干重。


1.4 数据处理
采用Microsoft Office Excel 2007 进行作图,用WPS 进行表格的制作,用SPSS 23.0 数据处理系统进行统计及差异显著性检验,显著度水平为0.05。
2 结果与分析
2.1 少花蒺藜草刺苞、模拟刺苞及异型种子的发芽特性
2.1.1 发芽率和发芽势 刺苞(M 型种子)、模拟刺苞内M 型种子及独立条件下M 型种子的发芽率分别为93.3%、90.0%和90.0%,三者之间差异不显著(P>0.05),表明刺苞对M 型种子的萌发无显著影响;刺苞内P 型种子(刺苞内同时发2 芽)的发芽率为40.6%,模拟刺苞内P 型种子的发芽率为41.7%,独立条件下P 型种子的发芽率为85.6%,刺苞内P 型种子与模拟刺苞内P 型种子的发芽率无显著差异(P>0.05),独立条件下P 型种子的发芽率分别比刺苞及模拟刺苞内P 型种子发芽率显著增加110.8%和105.3%,模拟刺苞内P 型种子保持较低的发芽率(图3),表明抑制P 型种子萌发的主要因素不是刺苞果皮而是M 型种子,M 型种子对P 型种子在刺苞内及模拟刺苞内的抑制率分别为52.5%和51.4%(图4)。单独M 型种子的发芽势为70.0%;单独P 型种子的发芽势为72.2%;刺苞内M 型种子的发芽势为68.9%,三者之间无显著差异(P>0.05);模拟刺苞内M 型种子的发芽势为48.3%,显著低于独立的M 型及刺苞(M 型种子)的31.0%和29.9%;模拟刺苞内P 型种子的发芽势为8.3%,独立条件下P 型种子的发芽势为68.3%,模拟刺苞内P 型种子的发芽势显著低于独立的P 型种子87.8%(图3)。

图3 少花蒺藜草刺苞及种子的发芽率及发芽势Fig.3 Germination rate and germination energy of bract and seed of C.incertus不同小写字母表示不同类型种子之间差异显著(P<0.05)。图中字母A 为模拟M 型、B 为模拟P 型、C 为M 型、D 为P 型、E 为刺苞、F 为刺苞内P 型处理。下同。Different lowercase letters indicated significant difference among different types at 0.05 level. The letters A is simulated M type,B is simulated P type,C is M type,D is P type,E is the bur,F is P type in the bur.The same below.
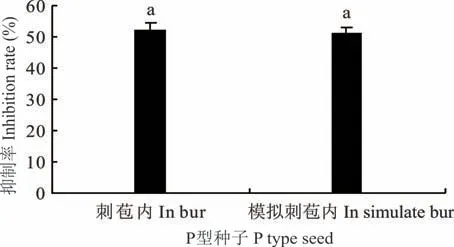
图4 M 型种子对P 型种子的抑制率Fig.4 Inhibition rate of M seed to P seed
2.1.2 发芽指数和活力指数 刺苞(M 型种子)、模拟刺苞内M 型种子及独立条件下M 型种子的发芽指数分别为6.47、5.23 和7.58,独立条件下M 型种子的发芽指数分别显著高于刺苞(M 型种子)及模拟刺苞内M 型种子17.2%和44.9%(P<0.05);模拟刺苞内P 型种子的发芽指数为0.75,独立条件下P 型种子的发芽指数为6.68,模拟刺苞内P 型种子的发芽指数显著低于独立条件下P 型种子88.8%(P<0.05)(图5)。
刺苞(M 型种子)、模拟刺苞内M 型种子及独立条件下M 型种子的活力指数分别为71.01、14.98 和87.50,刺苞及模拟刺苞内M 型的活力指数分别显著低于独立条件下M 型种子18.8%和82.9%(P<0.05);模拟刺苞内P型种子的活力指数为1.75,独立条件下P 型种子活力指数为59.83,模拟刺苞内P 型种子的发芽指数显著低于独立条件下P 型种子97.1%(P<0.05)(图5)。

图5 少花蒺藜草刺苞、种子的发芽指数及活力指数Fig.5 Germination index and vigor index of bract and seed of C. incertus
2.1.3 胚根及胚芽的生长速率 由图6 可知,少花蒺藜草刺苞及两异型种子胚根的初始萌发时间为24 h,在前3 d 胚根的生长速率表现为刺苞>P 型种子>M 型种子,3 d 后生长速率为M 型种子>P 型种子>刺苞。胚芽的初始萌发时间为24 h,在前3 d 的生长速率表现为M 型种子=刺苞>P 型种子,最终的生长速率为P 型种子>M 型种子>刺苞。图7 为模拟刺苞内M、P 型种子发芽图片,大部分只有M 型种子萌发。
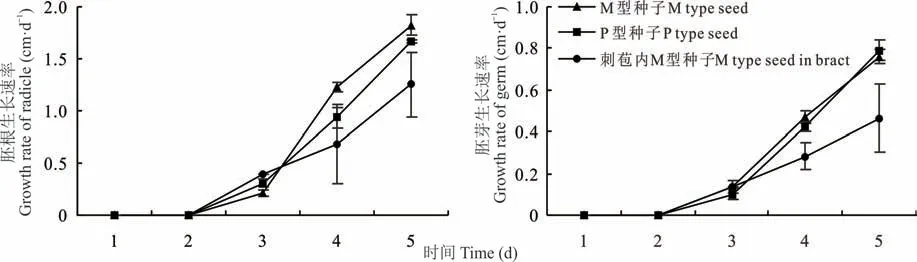
图6 少花蒺藜草刺苞、种子胚根及胚芽的生长速率Fig.6 Growth rate of spiny bract,seed radicle and germ of C.incertus
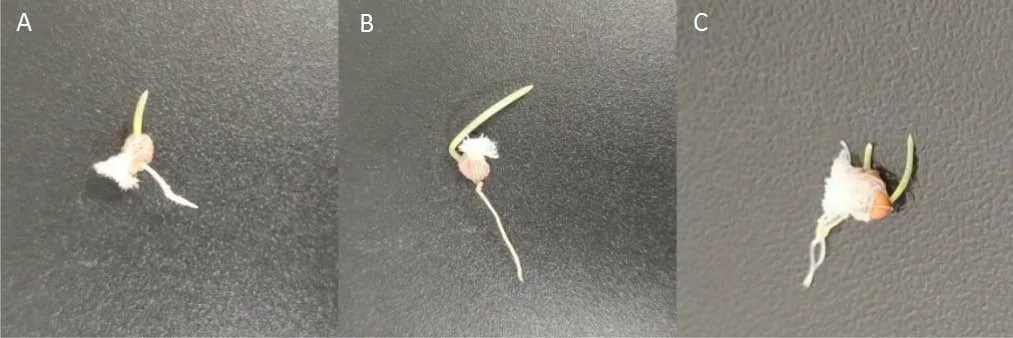
图7 模拟刺苞内只有M 型种子发芽(A,B)和M 型、P 型种子同时发芽(C)Fig.7 Only M type seeds germinated in the bract(A,B)and M type and P type seeds germinated simultaneously(C)
2.2 少花蒺藜草刺苞及异型种子幼苗的形态及生物量特征
从形态特征分析,刺苞萌发幼苗的叶面积分别显著高于独立的P 型种子及刺苞内P 型种子萌发的幼苗51.9%和84.3%,刺苞萌发的幼苗与M 型种子萌发的幼苗叶面积无显著差异;刺苞内P 型种子萌发幼苗的比叶面积分别显著高于M 型种子、P 型种子及刺苞萌发幼苗87.2%、68.3%和392.2%,M 型、P 型种子萌发幼苗的比叶面积无显著差异。
从生物量特征分析,刺苞萌发幼苗的地上生物量显著高于其他三者,M 型种子萌发幼苗的地上生物量低于刺苞76.7%,P 型种子萌发幼苗的地上生物量低于刺苞的82.7%,刺苞内P 型种子萌发幼苗的地上生物量低于刺苞的86.5%,M 型、P 型种子萌发幼苗的地上生物量无显著差异;刺苞萌发幼苗的地下生物量显著高于其他三者,M型种子萌发幼苗的地下生物量显著低于刺苞79.1%,P 型种子萌发幼苗的地下生物量显著低于刺苞68.5%,刺苞内P 型种子萌发幼苗的地下生物量低于刺苞97.1%,M 型与P 型种子的地下生物量无显著差异;P 型种子萌发幼苗的根冠比显著高于其他种子及刺苞萌发幼苗的根冠比,M 型种子、刺苞以及刺苞内P 型种子萌发幼苗的根冠比分别低于P 型种子50.8%、45.5%、88.1%,M 型种子与刺苞的根冠比无显著差异。
从生物量分配特征角度分析,M 型种子、刺苞以及刺苞内P 型种子萌发幼苗的根生物量比分别显著低于P 型种子萌发幼苗29.8%、26.3%和52.6%,M 型种子与刺苞无显著差异(表1)。结果表明,刺苞及种子所萌发的幼苗会根据获得的营养物质及对自身能力的支配产生不同的生长分配策略。
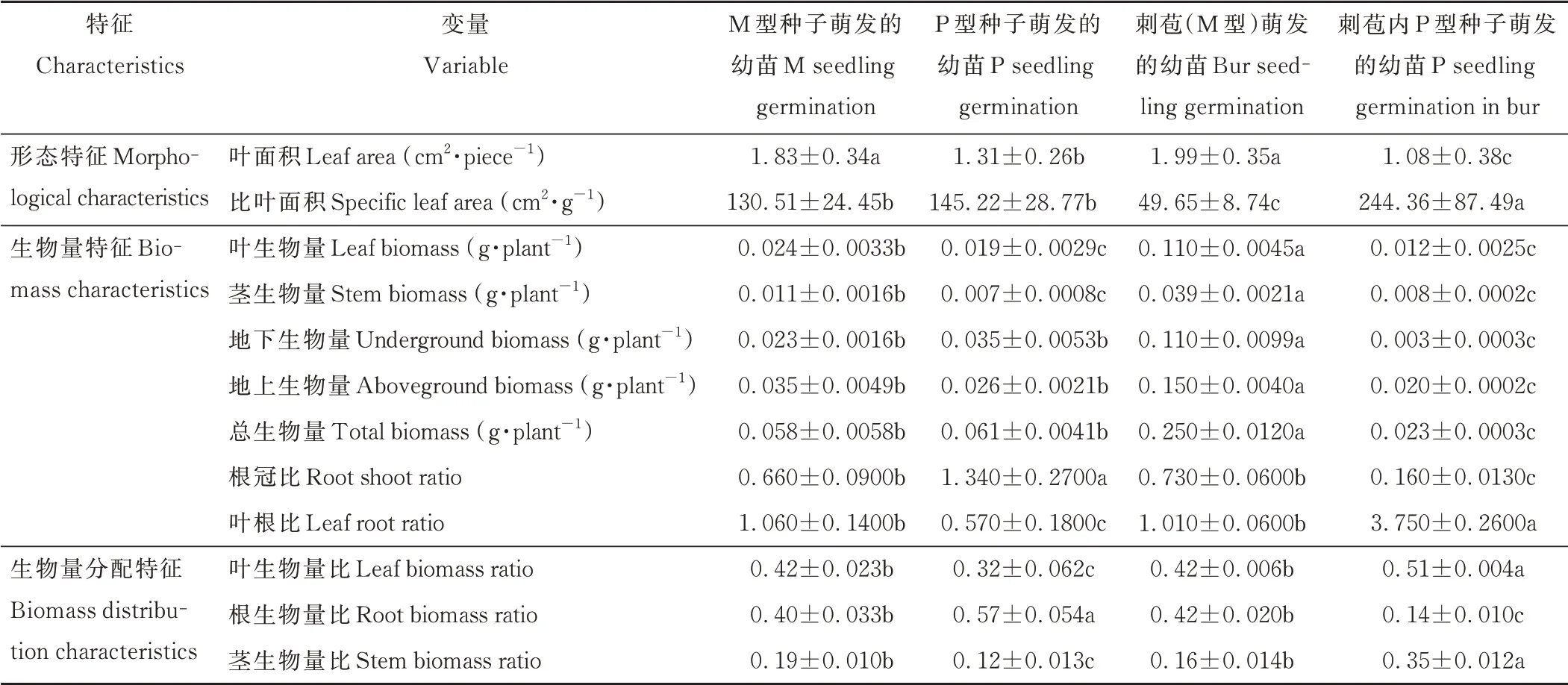
表1 少花蒺藜草刺苞、异型种子萌发幼苗的形态特征及生物量特征Table 1 Morphological and biomass characteristics of seedlings germinated with bract and heteromorphic seed of C.incertus
3 讨论
3.1 少花蒺藜草异型种子的萌发策略
任何植物在特定环境中的持续繁衍和生存都依赖其自身的功能特征,在植物的众多特性中,种子是植物物种延续、扩大种群生态位空间的生命载体[21−22],种子萌发是植物生长阶段极为重要的时期,发芽率、发芽势等是检验种子生活力、反映种子发芽能力和发芽时间的重要指标[23]。本研究结果中M 型种子在刺苞内外均具有较高的萌发率,表明刺苞果皮对M 型种子的萌发无显著影响;模拟刺苞试验表明,在无刺苞果皮的情况下,P 型种子仍具有较低萌发率,显示刺苞果皮对P 型种子的萌发无显著影响,结合野外调查,少花蒺藜草在自然条件下受到外界环境影响时,P 型种子会在季节周期内对环境进行感应(被激发),选择当年萌发生长完成生命周期或是选择次年在适宜环境条件下萌发生长拓展种群数量(采取机会主义策略),可见,刺苞果皮对两种异型种子的萌发均无显著影响,可见刺苞果皮的作用一是对种子进行保护,避免家畜啃食,二是由于刺苞果皮具有倒刺更易于通过人和牲畜携带进行传播扩散,从而扩大其在种群中的生态位[24]。P 型种子在独立条件下比非独立条件下萌发率高,说明M型种子对P 型种子的萌发产生了一定的抑制效应,前期试验表明,M 型种子的浸提液降低了P 型种子的萌发率,说明M 型种子可能存在某种化感物质对P 型种子的萌发产生了一定程度的影响,有关M 型种子内的化感物质将在后续的试验中进行研究;本研究结果中M 型种子在两种培养条件下均具有较好的发芽能力,马赫等[10]研究表明野榆钱菠菜(Atriplex aucheri)3 种异型种子的萌发物候不同步,大种子提前萌发2 d,较小种子提前萌发7 d,且大种子的出苗率显著高于另外两小种子的出苗率;王梦茹等[25]对钠猪毛菜(Salsola nitraria)的研究表明,异型种子中较大的种子具有更顽强的生命力和适应能力,较大种子具有更强地对光、水、肥资源的竞争和利用能力以及对资源缺乏环境的忍耐和适应能力[26],这也与魏梦浩等[27]对盐角草(Salicornia europaea)的研究结果一致,盐角草3 种类型的种子在大小和质量上有着显著差异,一般大粒种子有较大的胚,萌发和生长一般较快;小粒种子存在休眠,可形成种子库,确保物种延续。
3.2 少花蒺藜草刺苞及种子形成幼苗的生长特性
在不同的物种种群内,种子的大小变异主要发生在植物的个体内及个体间,种子大小和幼苗更新对策是植物生活史策略的重要组成部分[28]。从种子到幼苗阶段是植物种群生活史中亏损最严重的时期之一,而幼苗更新对策又是植物生活史对策中最重要的阶段,因为幼苗更新是植物种群扩大、扩散和延续的一个主要方面[29−31]。植物在幼苗阶段是通过将更多的资源分配到功能组织来更好地获取资源[32−33]。为了构建和维持植物正常的生长,幼苗主要将资源向茎投入,也可通过调节生物量的分配来更好地适应环境[34−35]。本研究中,刺苞形成的幼苗(刺苞中M 型种子萌发的幼苗)与独立条件下M 型种子形成的幼苗在形态特征及生物量特征方面均具有较高的优势,原因是种子大小与幼苗生物量呈显著正相关[36],潘慧超等[37]研究表明,相对较大的种子形成的幼苗生长快,获得的资源更多,具有更强的竞争能力,对植被的更新贡献更大;本研究中4 种处理条件下形成幼苗的生物量特征之间差异显著,表明少花蒺藜草异型种子及刺苞会根据自身所处环境对吸收到的营养物质进行分配从而调整自身的存活策略,这一结果与许多学者对植物受到资源限制时通常会调节植物地上部和地下部生物量资源以适应环境变化的研究结果一致[38−40]。根系是生长最重要的地下营养器官,植物为适应环境,根系表现出明显的可塑性,从而导致植物在根系生长、分布结构和地上部生物量分配上产生差异[41−42]。本研究中,少花蒺藜草两种异型种子根系生长速率的差异不显著,表明两种异型种子根系均具有较高的生长能力,当M 型种子形成的植株无法完成种群扩展的使命时,P 型种子可以迅速生根发芽以维持或补充种群数量。
4 结论
刺苞吸足水分后依旧采取保守萌发策略(大多数只发1 芽);M 型种子在适宜条件下迅速萌发,快速完成生活史得以维持种群数量,P 型种子在同样拥有相对较高萌发活力的同时作为后备力量补充种群数量,这种相互感应与配合的萌发及生长策略是少花蒺藜草在科尔沁沙地迅速扩散蔓延的重要原因之一。少花蒺藜草M、P 型种子补偿性萌发策略是其强入侵性的生物学基础,有效控制刺苞形成或对已形成刺苞进行灭活技术的研发是遏制少花蒺藜草种群蔓延的关键。
