黄麻与其他物种HD-ZIPⅠ和LEA14蛋白序列同源性及其盐胁迫表达分析
2022-06-06陈毓娜ManuelSebastianFiallos李云清祁建民徐建堂方平平林荔辉张立武陶爱芬
陈毓娜, Manuel Sebastian Fiallos, 李 静, 李云清, 祁建民, 徐建堂, 方平平,林荔辉, 张立武, 陶爱芬
(1.福建农林大学作物遗传育种与综合利用教育部重点实验室/福建省作物设计育种重点实验室;2.福建农林大学金山学院,福建 福州 350002)
黄麻(CorchorusL.),锦葵科黄麻属一年生草本韧皮纤维作物,是世界上最重要的长纤维作物之一,其产量和种植面积仅次于棉花[1].黄麻纤维由于具有吸湿、抗菌、吸音等优异性能而被运用在服饰、医疗、工业等产业中[2-3].近年来,菜用黄麻、重金属吸附专用黄麻等资源创新与产业化应用也取得了一定进展[4].盐胁迫是影响农作物生长的重要环境因素之一[5-6].目前,有关高等植物耐盐基因的研究主要集中在离子转运蛋白基因、渗透调节相关基因、信号传导相关基因、细胞抗氧化相关基因等方面[7].
同源异形域-亮氨酸拉链(homeodomain-leucine zipper, HD-ZIP)是一类植物特有的转录因子家族,在植物的非生物胁迫响应、病菌防御以及形态建成中发挥重要的调控作用[8-11]. HD-ZIP家族可分为4个亚家族,其中,HD-ZIPⅠ主要参与植物对非生物胁迫的响应过程[12].HD-ZIP家族基因的表达情况在许多植物如大豆、黄瓜、苜蓿、桑树等[13-16]上已被广泛研究.LEA蛋白(late embryogenisis abundant proteins),即胚胎发育后期丰富蛋白,是植物中广泛存在的一类小分子亲水性蛋白,最早从棉花胚胎中分离获得[17-18].LEA蛋白广泛参与植物非生物胁迫(尤其是失水胁迫)的应答过程,并在提高植物抗逆性方面发挥重要的功能[19].研究表明:超表达Os-LEA3-2能够提高水稻的耐旱性及耐盐性[20];大麦中的LEA-3组基因HVA1在水稻、玉米、燕麦、剪股颖草、桑树和菜豆中异源超表达,均不同程度地提高了转基因植物的抗旱性或耐盐性[21].根据表达模式和序列的不同可将植物中的LEA蛋白分成6个组,其中,在许多植物物种中发现的一类LEA蛋白是拟南芥的At1g01470.1,被归类为“LEA14蛋白”[22].研究表明,LEA14基因在植物不同组织中均有表达,其在拟南芥、甘薯、烟草中的过量表达提高了这些作物对盐的耐受性[23-25].
进化树在粮食作物、蔬菜和牧草等植物的基因分类和进化关系分析中得到了广泛应用[16,26].研究表明,从被子植物开始,HD-ZIP基因家族就稳定存在4个亚族,推测在形成4个亚族前,HD-ZIP分为两组,其中一组分化成Ⅰ和Ⅱ亚族,而另一组则分化成Ⅲ和Ⅳ亚族[27].系统发育分析发现,单子叶植物和双子叶植物分化后,在独自进化的过程中某些分支的成员扩张较快,单子叶植物和双子叶植物在分化之前,就已经建立了HD-ZIPⅣ亚家族的多样性[28].目前,学者们对多种作物如小麦、玉米、烟草等[29-31]的LEA基因家族进行了鉴定和系统进化分析.但有关黄麻HD-ZIPⅠ基因家族及LEA14的系统进化和表达情况尚未见报道.本研究对黄麻HD-ZIPⅠ和LEA14蛋白序列同源性及其在盐胁迫下的基因表达进行分析,以期为了解黄麻HD-ZIPⅠ基因家族和LEA14基因的进化关系以及黄麻的耐盐机制提供依据.
1 材料与方法
1.1 材料及盐胁迫处理
本试验以前期筛选的耐盐性有差异的2份长果种黄麻品种为材料,分别为较耐盐品种福农1号和对盐较敏感品种中黄麻1号,种子由福建省南方麻类种质资源共享平台提供.在育苗盘中播种,用购自Pindstrup公司的营养土作为培养基质;待黄麻幼苗长至三叶一心时,移栽到Hoagland营养液中进行培养,每3 d更换一次Hoagland营养液;幼苗长至五叶一心时,每个品种随机选取15株,用含200 mmol·L-1NaCl 的Hoagland营养液进行盐胁迫处理.分别在盐处理0、24、48和72 h取样(叶片和根系),经液氮冷冻后放于-80 ℃冰箱保存备用.
1.2 方法
1.2.1 多序列比对与系统进化树构建 从NCBI网站(https://www.ncbi.nlm.nih.gov)中获得长果种黄麻蛋白质序列,拟南芥、大豆、棉花的LEA14蛋白质序列通过UniProt网站(https://www.uniprot.org/)获得,拟南芥的HD-ZIPⅠ蛋白序列从TAIR数据库 (http://www.arabidopsis.org/)下载,水稻、大豆的HD-ZIPⅠ转录因子家族序列从PlantTFDB网站(http://planttfdb.gao-lab.org/)下载.利用TBtools软件中的Blast进行序列对比,找到相关基因在黄麻中的同源序列;同时,通过SMART(http://smart.embl-heidelberg.de/)和在线Web CD-Search Tool获得蛋白质结构图,筛选与相关基因蛋白质结构域相同的黄麻蛋白质序列[32];最后采用Clustal W将黄麻基因序列与拟南芥、大豆、棉花等的蛋白质序列进行多序列比对,通过软件MEGA 7.0用邻近法(neighbor joining, NJ)构建系统进化树[33].采用TBtools的Simple Ka/Ks Calculator计算非同义替换(Ka)和同义替换(Ks)之间的比例(Ka/Ks值)及进行共线性分析.
1.2.2 DNA与RNA提取 利用TIANGEN植物基因组提取试剂盒(离心柱型)提取样品的DNA,并置于-20 ℃冰箱保存备用.采用北京华越洋生物科技有限公司的快速通用RNA提取试剂盒(0416-50 gk)提取样品的RNA,然后置于-80 ℃冰箱保存备用.通过1.2%(质量分数)的琼脂糖凝胶电泳和分光光度计(NanoDrop 2000, Thermo)对核酸质量进行检测.
1.2.3 cDNA第一条链的合成 按照TransScript反转录试剂盒(AQ301-01)的说明合成cDNA第一条链,并经1.2%(质量分数)琼脂糖凝胶电泳检测.
1.2.4 引物设计与验证 通过idta primers网站(https://www.idtdna.com/)在线设计引物,利用TBtools软件选择引物,片段大小为70~200 bp,引物长度为18~22 bp,引物所对应的模板序列复性温度(Tm)为59~65 ℃,引物GC含量为50%~60%.参照Hossain et al[34]的方法对引物的有效性进行验证.本试验所用引物的名称和序列见表1,其中,UBC2(the ubiquitin-conjugating enzyme like protein)为内参基因.
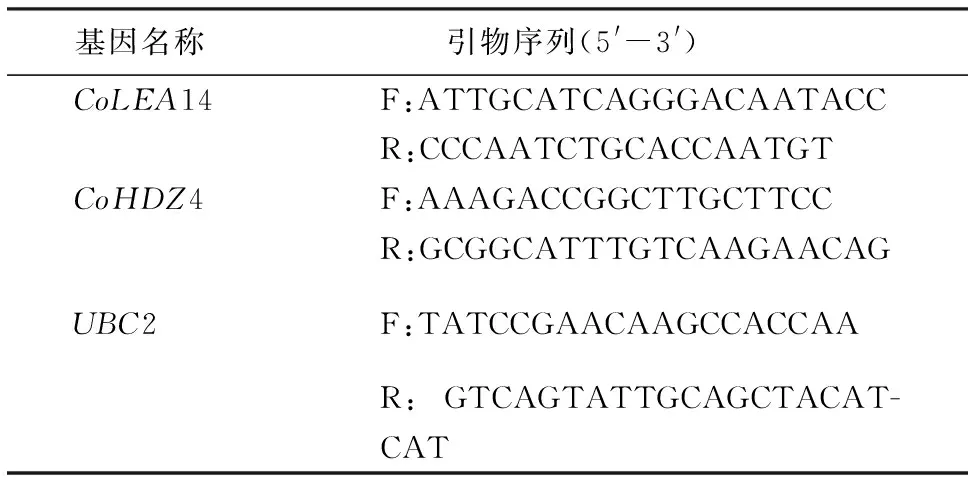
表1 本研究所用引物名称和序列Table 1 Names and sequences of primers used in the study
1.2.5 qRT-PCR反应 利用PerfectStartTMGreen qPCR SuperMix试剂盒(AQ602-24)进行qRT-PCR反应,采用2-△△Ct法计算各基因的相对表达量[35].
2 结果与分析
2.1 黄麻HD-ZIPⅠ家族蛋白序列进化分析
基于拟南芥、水稻、大豆与黄麻4个物种的HD-ZIPⅠ家族基因编码蛋白质序列所构建的系统进化树见图1,根据Eva et al[36]定义的拟南芥中HD-ZIPⅠ亚家族的支系标记该进化树的支系分组.从图1可以看出,在黄麻中共鉴定获得10个HD-ZIPⅠ编码蛋白(以OMO或OMP开头),被分成7个亚组.其中,β1亚组包含2个黄麻蛋白,分别为OMO73039.1和OMP01294.1,前者与大豆有较近的进化关系,而后者处于一个较独立的分支.β2亚组中包含3个黄麻蛋白(OMO69727.1、OMO55126.1和OMO52137.1)、6个大豆蛋白和1个拟南芥蛋白,其中,OMO55126.1和OMO52137.1与大豆有较近的亲缘关系,而OMO69727.1与拟南芥和大豆均有同源性,但与大豆的进化关系更近.ε亚组中有1个黄麻蛋白(OMO83981.1)及2个大豆蛋白和2个拟南芥蛋白,OMO83981.1同样表现出与大豆更近的亲缘关系.γ亚组中有3个黄麻蛋白,分别为OMP08645.1、OMO97833.1和OMP02850.1,其中,前2个基因与大豆的亲缘关系较近,而第3个基因处于一个较独立的分支,其与大豆和拟南芥均有一定的同源性.在δ亚组的10个成员中,只有1个黄麻蛋白(OMO84308.1),位置相对较独立,但与拟南芥和大豆均有一定的进化关系,与水稻关系较远.α、Φ两个亚组均没有黄麻HD-ZIPⅠ蛋白,其中,α亚组包含拟南芥和水稻蛋白,Φ亚组只包含1个拟南芥蛋白(ATHB54).可见,黄麻HD-ZIPⅠ的蛋白序列与双子叶植物大豆和拟南芥的相似度高于与单子叶植物水稻的相似度[37].
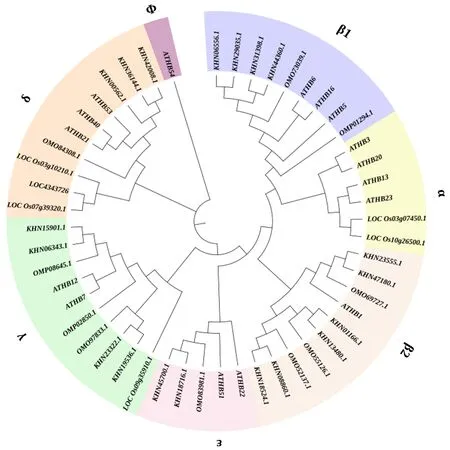
ATH代表拟南芥,KHN代表大豆,LOC代表水稻,OM代表黄麻.图1 拟南芥、水稻、大豆和黄麻HD-ZIPⅠ家族的系统进化树Fig.1 Phylogenetic tree of HD-ZIPⅠ family in Arabidopsis, rice, soybean and jute
为进一步明确黄麻与拟南芥和水稻HD-ZIPⅠ基因分化的情况,计算了Ka/Ks值(表2、3),并进行了共线性分析(图2).由表2和图2可知,黄麻HD-ZIPⅠ家族8个基因与拟南芥的8个基因有共线性关系,这些基因主要位于第2和第5染色体上,第1、第3和第4染色体上也有分布,其Ka/Ks值在0.11~0.27之间,它们之间的分化时间最长为1.214 6亿年,最短为0.473 0亿年.而黄麻HD-ZIPⅠ家族中仅有2个基因与水稻有共线性关系,位于第2和第4染色体上,分别对应水稻的3个基因,Ka/Ks值在0.07~0.26之间,它们之间的分化时间最长为1.478 8亿年,最短为0.673 0亿年(表3、图2).将与拟南芥和水稻均有同源性的2个基因(编号Co.02G0036900和Co.04G0025100)进行比较,发现它们与水稻在更早之前开始分化,而与拟南芥的分化时间较晚.如黄麻Co.02G0036900与水稻分化的时间为1.321 6亿年,显著长于其与拟南芥的分化时间(0.847 9亿年).以上结果表明,黄麻HD-ZIPⅠ家族的基因与双子叶植物拟南芥的进化关系比与单子叶植物水稻更近.
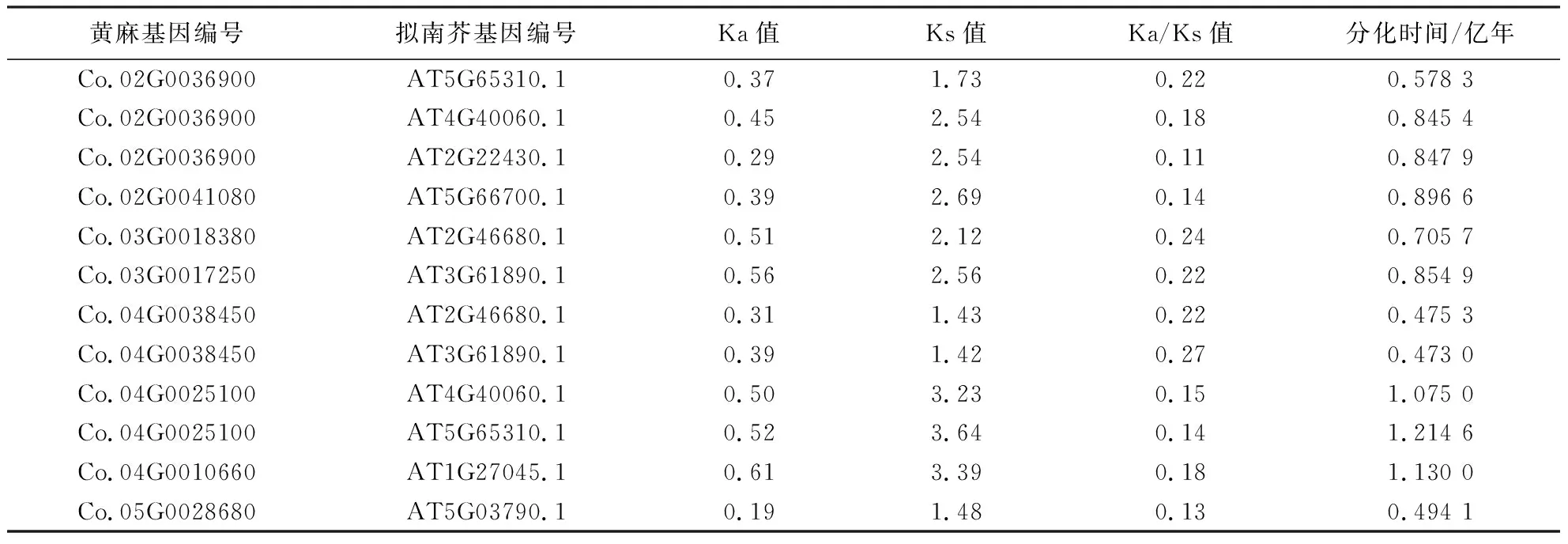
表2 黄麻与拟南芥HD-ZIPⅠ基因家族的Ka/Ks值和分化时间Table 2 Ka/Ks ratio and divergence time of HD-ZIPⅠ gene family between jute and Arabidopsis
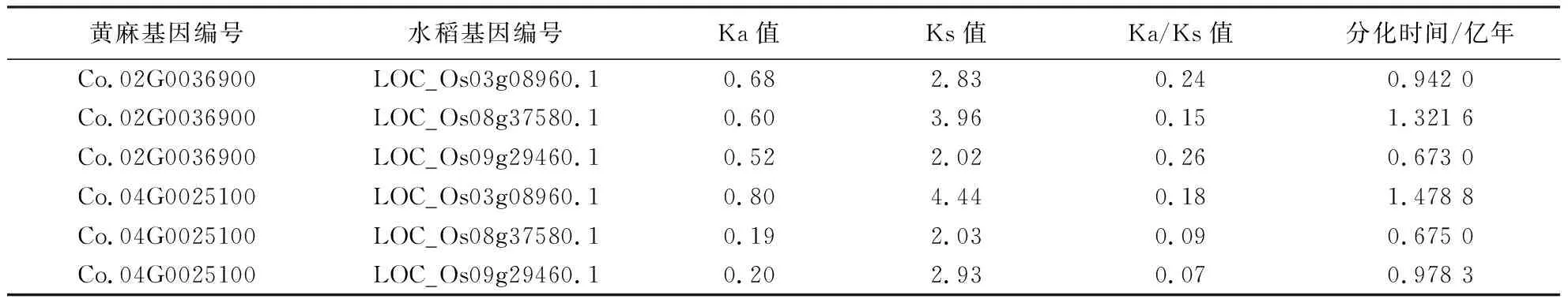
表3 黄麻与水稻HD-ZIPⅠ基因家族的Ka/Ks值和分化时间Table 3 Ka/Ks ratio and divergence time of HD-ZIPⅠ gene family between jute and rice
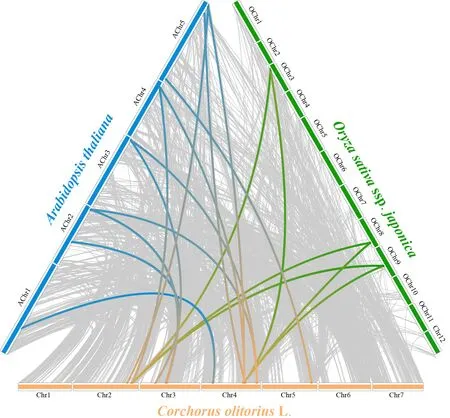
图2 黄麻与水稻和拟南芥HD-ZIPⅠ基因家族的共线性分析Fig.2 Synteny analysis of HD-ZIPⅠ genes in jute, rice and Arabidopsis
2.2 黄麻LEA14蛋白序列进化分析
前期分析发现,黄麻LEA蛋白家族中仅LEA14基因的蛋白序列与其他物种有同源性,因此,利用拟南芥、棉花、大豆和黄麻4个物种的LEA14蛋白序列构建了系统进化树(图3).从黄麻中仅鉴定获得1个与拟南芥、大豆和棉花LEA14蛋白具有同源性的蛋白(OMP00313.1),其与棉花(XP016708350)属于同一个分支,相似度为86%,表明黄麻的LEA14基因与棉花的进化关系最近.同时,OMP00313.1和 XP016708350所构成的小分支,与3个大豆蛋白处在同一个较大的分支内,表明黄麻的LEA14基因与大豆也有较近的进化关系.
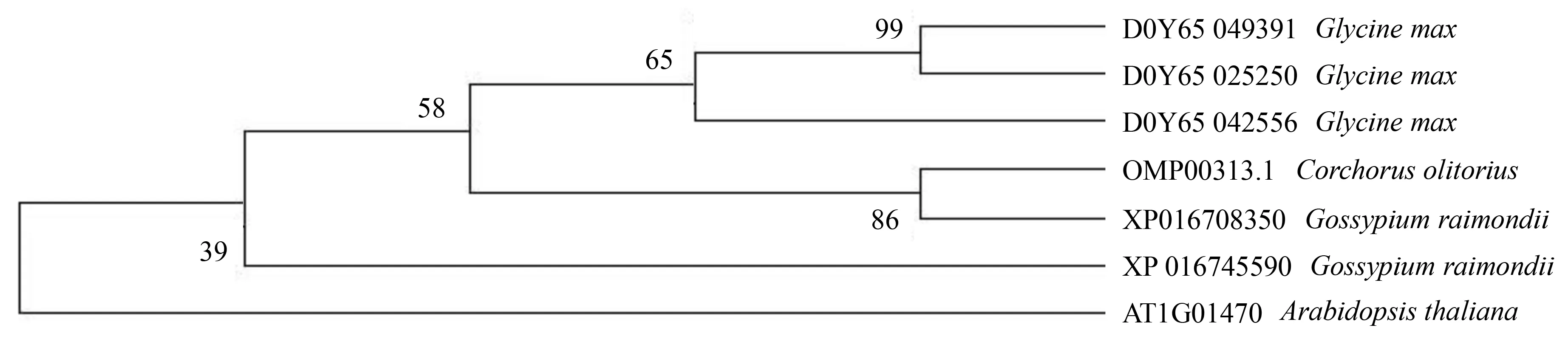
图3 拟南芥、大豆、棉花和黄麻LEA14蛋白的系统进化树Fig.3 Phylogenetic tree of LEA14 proteins in Arabidopsis, soybean, cotton and jute
2.3 黄麻总RNA质量检测与引物验证
图4表明,本研究所提取的黄麻 RNA完整性较好,无DNA残留,无明显拖尾降解或受污染的现象.用吸光光度计检测所提取RNA的质量,发现D260 nm/D280 nm的值在1.9~2.2之间,D260 nm/D230 nm的值大于2.0,说明RNA纯度较高.可见,本研究所提取的RNA质量较好,可以用于后续的试验.

左侧1~4为中黄麻1号叶片RNA,5~8为其根系RNA;右侧1~4为福农1号叶片RNA,5~8为其根系RNA.图4 黄麻总RNA电泳图Fig.4 Electropherogram of total RNA from jute
用HDZ4和LEA14及内参基因UBC2的引物进行常规PCR扩增,根据测序结果进行对比,基于验证成功的引物选择qRT-PCR的引物,以黄麻反转录cDNA第一链为模板进行PCR扩增,获得的产物经凝胶电泳检测,结果如图5所示.所得条带单一且清晰,没有非特异性扩增条带,表明引物特异性强,片段大小与预期相一致,符合后续的qRT-PCR试验要求.
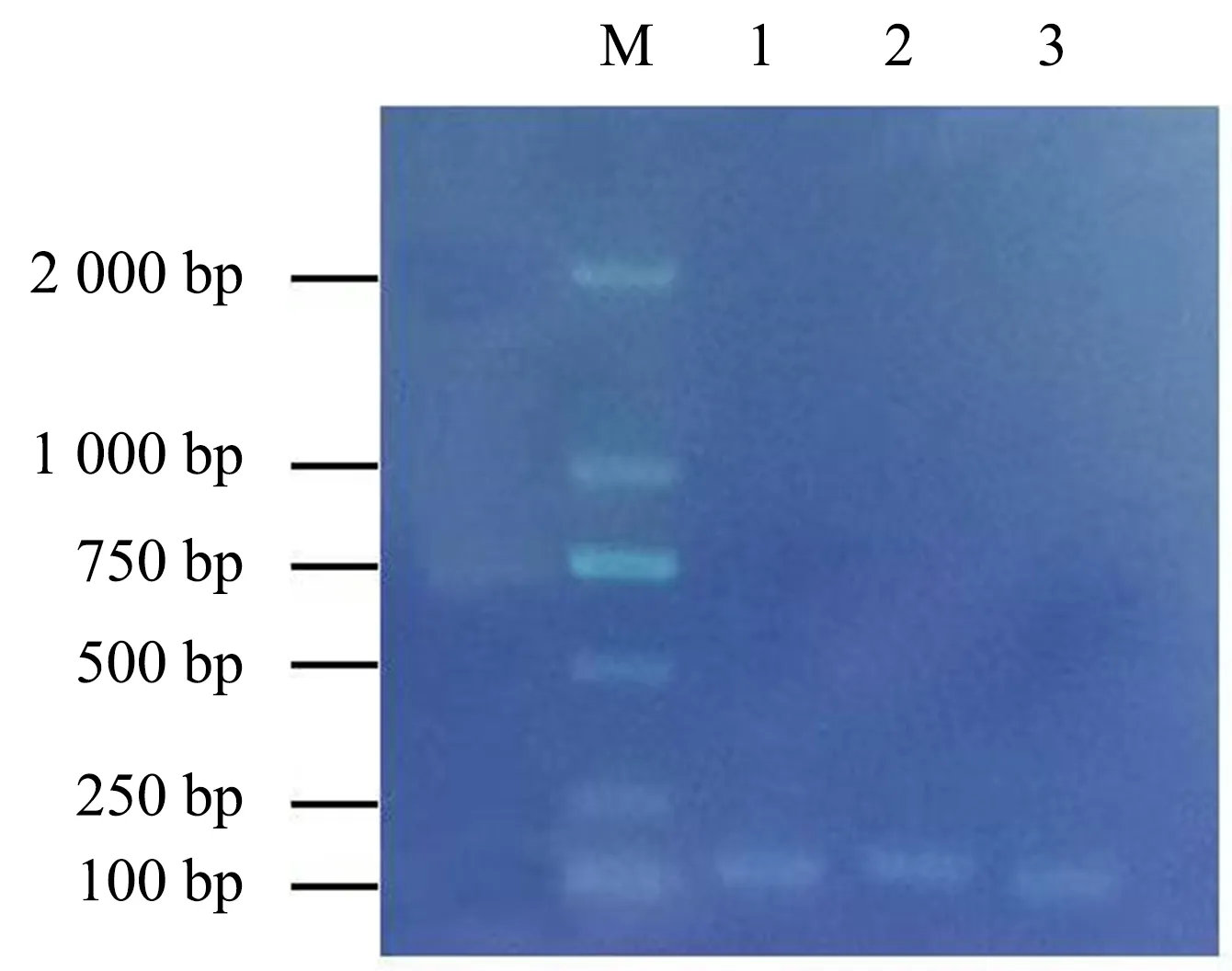
M为Marker,1~3分别为UBC2、HDZ4和LEA14基因.图5 qRT-PCR引物电泳结果Fig.5 Electropherogram of qRT-PCR primers
2.4 盐胁迫下黄麻LEA14基因的表达分析
由图6A可知,随着盐胁迫时间的推移,LEA14在中黄麻1号叶片中的表达量呈现先下调再上调后又下调的规律,盐胁迫48 h时LEA14基因表达量最高;LEA14在中黄麻1号根部的表达规律与叶片中相同,盐胁迫48 h时表达量最高,但72 h时表达量几乎为0.此外,盐胁迫24 h时中黄麻1号叶片与根部LEA14的表达量接近,而盐胁迫48和72 h时LEA14在叶片中的表达量明显高于根部.
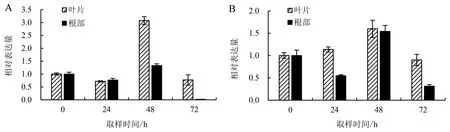
图6 LEA14在中黄麻1号(A)和福农1号(B)叶片和根部的表达量Fig.6 Expressions of LEA14 in leaves and roots of Zhonghuangma No.1 (A) and Funong No.1 (B)
LEA14在福农1号叶片中的表达量呈现先上调再下调的趋势,盐胁迫48 h时达到最高值;而LEA14在福农1号根部的表达规律为先下调再上调后又下调,胁迫48 h时表达量达到最高水平.此外,盐胁迫24和72 h时LEA14在叶片中的表达量为根部的2倍左右,而48 h时在叶片中的表达量略高于根部(图6B).
比较LEA14在两个黄麻品种中的表达量(图6)发现,盐胁迫24 h时,福农1号叶片中LEA14的表达量明显高于中黄麻1号,而此时二者根部LEA14的表达量差别不大.盐胁迫48 h时,中黄麻1号叶片中LEA14的表达量显著高于福农1号,而福农1号根部LEA14的表达量较中黄麻1号高.盐胁迫72 h时,福农1号叶片和根部LEA14的表达量均高于中黄麻1号.整体来看,在盐胁迫条件下,LEA14在福农1号叶片和根部的表达量高于中黄麻1号(除48 h叶片外).
2.5 盐胁迫下黄麻HDZ4基因的表达分析
由图7可以看出:随着盐胁迫时间的推移,HDZ4在中黄麻1号叶片和根部的表达量变化不明显,且同一个时间点,HDZ4在根部的表达量稍高于叶片,差异不显著;HDZ4在福农1号叶片和根部的表达量几乎没有差异,且在不同时间点的表达量差异也不明显.
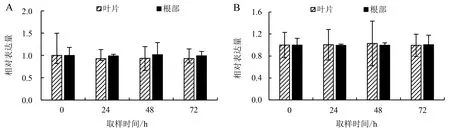
图7 HDZ4在中黄麻1号(A)和福农1号(B)叶片和根部的表达量Fig.7 Expressions of HDZ4 in leaves and roots of Zhonghuangma No.1 (A) and Funong No.1 (B)
3 讨论
3.1 HD-ZIPⅠ亚家族及LEA14蛋白的进化关系
系统发育树不仅可以分析不同物种亲缘关系的远近,而且可以直观表现不同物种的发生关系[38-39],该方法在LEA基因家族的分类和进化关系研究中得到了广泛应用[40].本研究发现,黄麻的1个LEA14基因编码蛋白与棉花的亲缘关系比与其他作物的关系更近.黄麻与棉花同属锦葵科,具有较好的共线性.与棉花相比,黄麻的遗传研究相对落后,而利用棉花遗传信息加强对黄麻的遗传研究,可为黄麻和棉属的比较基因组学和遗传研究提供依据,对于发现黄麻新基因、创造新种质、培育新品种等均具有重要意义.
前人也利用进化树分析的方法,确定了不同作物HD-ZIP蛋白的分类和进化关系,发现HD-ZIPⅠ为成员最多的亚家族.钟宣伯[16]揭示了拟南芥、水稻、野生大豆和大豆 HD-ZIPⅠ蛋白的进化关系,推测 HD-ZIPⅠ亚组的分化起始于单、双子叶植物分化之前,并在单、双子叶植物分化后持续分化.本研究鉴定到10个与其他作物基因家族具有同源性的黄麻HD-ZIPⅠ蛋白,其与大豆、拟南芥(双子叶)的序列相似度高于与水稻(单子叶)的相似度,且与拟南芥有更多的共线性基因,并且与拟南芥的分化时间远远短于与水稻的分化时间,表明HD-ZIPⅠ家族基因在单、双子叶中存在功能分化[37].该结果与钟宣伯[16]的研究结果类似.
3.2 盐胁迫下HDZ基因的表达
HD-ZIP转录因子可通过激活抗氧化系统、调节渗透稳态、维持Na+/K+的稳态以及调控下游胁迫响应基因参与植物对盐胁迫的应答.研究表明:HD-ZIP基因可以提高抗氧化酶的活性和一些可溶性有机物质的积累[41];玉米HD-ZIP转录因子HD-ZIPⅠ亚家族参与脱落酸(abscisic acid, ABA)介导的盐胁迫响应途径,在高浓度盐胁迫下,转基因植株存活率远高于野生型[42];罗少[11]发现,在所鉴定的33个桑树HD-ZIP基因中,6个基因在高盐处理下被显著诱导,推测这6个基因与桑树的抗盐能力有关.本研究中,HDZ4基因在不同品种以及盐胁迫的不同时间的表达量差异均不明显.这可能是由于取样间隔时间(24 h)太长,推测黄麻HDZ4基因可能在盐胁迫后短时间(12 h)内表达上调或下调,之后表达趋于稳定,对此有待进一步验证.
3.3 盐胁迫下LEA基因的表达
LEA蛋白是一种亲水性蛋白,主要应对与脱水相关的非生物逆境胁迫,包括耐盐、抗旱、抗冻等[43].LEA基因在不同作物中有不同的表达趋势,且过量表达该基因能增强作物对逆境胁迫的耐性.例如:将大麦LEA基因HVA1转化至水稻中,转基因植株抵御盐胁迫的能力较野生型明显增强[44];拟南芥Atlg01470.1基因在干旱、盐、强光3种胁迫下的表达量比非胁迫条件下高5倍[22];Jia et al[23]从拟南芥中鉴定获得一种LEA蛋白AtLEA14,其过量表达的转基因拟南芥对盐的耐受性增强;甘薯在盐胁迫条件下,叶片和根系中IbLEA14的表达量从12 h后开始上调,24 h后下调[24];天山雪莲的SiLEA14基因,在低温和盐胁迫24 h时表达量达到高峰,并且该基因过量表达能提高烟草的耐盐能力[25].本研究中:LEA14在盐胁迫处理的黄麻中也基本呈现先上调后下调的规律,且盐胁迫48 h时是该基因被诱导表达的关键时间点;同时,LEA14在不同黄麻品种叶片中的表达量均高于根部,表明该基因在不同植物组织中的表达水平存在差异;另外,LEA14在较耐盐品种福农1号根部和叶片中的表达量基本高于在较敏感品种中黄麻1号中的表达量,这表明LEA14的上调表达与植物对盐胁迫的耐性有关.后续可通过该基因的过量表达研究其提高黄麻耐盐性的作用机理,为高产、优质、耐盐黄麻新品种的选育提供理论依据.
4 结论
本研究在黄麻中鉴定到10个与其他作物具有同源性的HD-ZIPⅠ蛋白,发现其与双子叶植物蛋白序列的相似度高于与单子叶植物的相似度;同时,鉴定到1个与其他作物同源性较高的黄麻LEA14蛋白OMP00313.1,其与棉花的同源性最高,进化关系最近.另外,在盐胁迫处理下,HDZ4在不同黄麻品种、不同组织及不同时间点的表达量没有明显差异;而LEA14在2个黄麻品种叶片中的表达量高于在根部的表达量,并且在较耐盐品种福农1号中的表达量总体上高于在较敏感品种中黄麻1号中的表达量;同时,盐胁迫48 h时是LEA14基因表达的关键时间点.
