牛角花齿蓟马成虫触角感器的扫描电镜观察
2022-06-03赵晓东崔晓宁胡桂馨刘艳君温雅洁
赵晓东,崔晓宁,胡桂馨,刘艳君,温雅洁
(甘肃农业大学草业学院,草业生态系统教育部重点实验室,甘肃省草业工程实验室,中-美草地畜牧业可持续发展研究中心,兰州 730070)
牛角花齿蓟马Odontothripsloti属缨翅目Thysanoptera蓟马科Thripidae齿蓟马属Odontothrips,是我国北方苜蓿田蓟马类优势害虫(吴永敷等, 1990; 张奔等, 2016)。该害虫主要行孤雌生殖,在苜蓿营养生长期偏好取食植株的心叶,导致苜蓿叶片缩小、光合效率降低、生长受阻,待苜蓿开花后则转移到花器中为害,严重影响苜蓿的产量与品质(张蓉等, 2003; 贺春贵等, 2007)。由于牛角花齿蓟马体小,取食方式隐蔽,繁殖力强,世代重叠严重,极易产生抗药性,加之苜蓿作为饲草产品的特殊功能,使用化学防治具有局限性(侯军和贺春贵, 2006; 花蕾等, 2007)。因此,发展和利用植物源诱剂或昆虫信息素的绿色监测和防控技术,对控制此虫危害,保障苜蓿产业健康发展具有重要意义。
触角是昆虫最主要的嗅觉和触觉器官,分布有丰富的触角感受器,不同类型的感器其规律分布、类型和数量在雌、雄虫之间往往具有差异(Schneider, 1964; Zacharuk, 1980)。昆虫通过触角各种感器的综合作用感知环境中化学、物理等刺激因子,从而调节其行为反应(那杰等, 2008; Mithoferetal., 2018)。如入侵害虫西花蓟马Frankliniellaoccidentalis可以利用其灵敏的嗅觉系统感受寄主植物释放的挥发物组分或信息素类化合物变化,指导其寄主定位、取食、栖息、选择配偶和生殖场所、防御和迁移等行为(Caoetal., 2014; Teulonetal., 2017; Caoetal., 2019)。因此,明确触角嗅觉感器的形态结构有利于探究蓟马嗅觉识别的生理和分子机制。
目前,已报道的蓟马类害虫触角感器方面的研究有西花蓟马(丁艳红等, 2010; 李维娜和冯纪年, 2013)、塔六点蓟马Scolothripstakahashii(徐洪涛等, 2011)、花蓟马Frankliniellaintonsa、禾花蓟马Frankliniellatenuicornis(李维娜和冯纪年, 2013)、茶棍蓟马Dendrothripsminowai(吕召云等, 2015)、茶黄蓟马Scirtothripsdorsalis(黄丽莉等, 2016; Zhuetal., 2017)、豆花蓟马Megalurothripsusitatus、棕榈蓟马Thripspalmi(Wangetal., 2019)、以及美洲棘蓟马Echinothripsamericanus(Huetal., 2020)。综合以上研究,发现蓟马触角感器主要有7种类型,即Böhm氏鬃毛(Böhm bristles, BB)、刺形感器(sensilla chaetica, SCh)、锥形感器(sensilla basiconca, SB)、钟形感器(sensilla campaniformia, SCa)、腔锥形感器(sensilla coeloconica, SCo)、腔形感器(sensilla cavity, SCav)和毛形感器(sensilla trichodea, ST),以及一类表皮衍生物——微毛(microtrichia, Mt)。目前,对牛角花齿蓟马的触角感器形态结构尚不清楚,其识别寄主挥发物或信息素的嗅觉感受机制有待探索。因此,本研究通过扫描电子显微镜观测牛角花齿蓟马的触角超微结构,以期对深入研究其与植物互作的化学生态机制提供基础。
1 材料与方法
1.1 供试昆虫
2020年7-8月,在甘肃农业大学校内牧草基地紫花苜蓿MedicagosativaL.试验田采集牛角花齿蓟马成虫。选取雌、雄虫各15头,在体视解剖镜下用刀片切取成虫头部,置于装有75%乙醇的1.5 mL离心管中,4℃保存备用。
1.2 样品制备与观察
样品固定:将蓟马头部先用去离子水、生理盐水和75%乙醇依次清洗两次,再用超声波清洗仪洗涤2次,每次15 s,结束后在样品管中加入2.5%戊二醛溶液,4℃固定10 h。
固定后处理:将固定后的样品用75%乙醇洗涤2次,再加入磷酸缓冲液(0.2 M,pH 7.2)洗涤3次。洗涤后的样品中依次加入30%、50%、70%、90%和100%乙醇溶液,进行浓度梯度脱水,每次15 min。然后用无水乙醇稀释叔丁醇,分别配置成25%、50%、75%叔丁醇乙醇溶液,将样品进行逐级置换,每次15 min。最后将样品利用冷冻干燥机(VFD-21St-BUOH)干燥处理,将样品用双面导电胶粘到样品台上,置于日立MC-100型离子溅射仪内喷金,最后在扫描电子显微镜(Hitachi-S 3400N)下观察拍照。
1.3 图像处理及数据分析
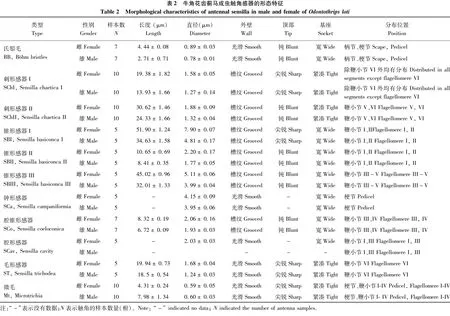
牛角花齿蓟马触角感器类型鉴定参考Schneider(1964)和Shields(2008)的方法。利用Photoshop CS 5.0软件对触角长度和宽度(中部),以及感器的长度和基部直径进行测量,用平均值±标准误(Mean±SE)表示。测量具体步骤:选择Photoshop CS 5.0“分析”选项中的“设置测定比例”,标定10 μm长度代表的像素值,以此为标准;将实际测到的触角或感器的长度或宽度的像素值,换算成实际测量长度值(μm)。所有数据采用SPSS 19.0软件分析,同一性别成虫的不同触角节间的长度或宽度显著性差异采用单因素方差分析(one way ANOVA, Tukey HSD, α=0.05),比较雌、雄虫之间的同一触角亚节的长度或宽度的显著性差异采用独立样本t检验(Student’s t test, α=0.05)。
2 结果与分析
2.1 触角形态结构
牛角花齿蓟马触角着生在头顶两复眼中间,雌、雄虫形态相似,呈丝状,共8节,从基部起依次为柄节Scape、梗节Pedicel、鞭节Flagellμm,雌虫触角314.22±7.21 μm明显长于雄虫260.58±5.69 μm(t=7.014,P<0.001),且每亚节间的长度存在显著差异(雌F=183.66,P<0.001;雄F=239.72,P<0.001)。柄节粗圆,表面较光滑,感器分布极少,雌虫比雄虫明显粗壮,长度雌虫为28.00±0.81 μm,雄虫为21.60±0.89 μm(t=5.326,P<0.001);宽度雌虫为37.00±1.19 μm,雄虫为29.82±1.17 μm(t=4.304,P=0.003)。梗节较柄节细长,感器分布也较柄节多,长度和宽度雌、雄虫之间均无显著差异。鞭节最为细长,每鞭小节呈梭形,具有丰富的感器类型和数量,分为6个小节(FI~FVI节),其中FI、FII和FII、FIII的节间明显缢缩,便于活动(图1-A, B)。雌虫的FII~FVI均较雄虫的粗长,就长度而言,其中FII(雌64.22±1.88 μm,雄53.02±1.37 μm,t=4.826,P<0.001)、FIV(雌63.36±1.71 μm,雄45.58±1.75 μm,t=7.270,P<0.001)节最长,其次是FI(雌50.74±1.50 μm,雄52.26±1.27 μm,t=0.787,P=0.454)、FIII(雌43.70±1.66 μm,雄32.66±0.99 μm,t=5.714,P<0.001)节,第FV(雌12.12±0.83 μm,雄8.48±0.37 μm,t=3.993,P=0.004)和FVI(雌17.16±1.12 μm,雄12.90±0.52 μm,t=3.441,P=0.009)节最短小(表1)。

图1 牛角花齿蓟马雌虫与雄虫触角形态Fig.1 Morphosis of the female and male Odontothrips loti adult antennae注:A,雌虫触角;B,雄虫触角;SC,柄节;PE,梗节;FL,鞭小节;FI-F6,鞭小节I-VI。Note: A, Female antennae; B, Male antennae; SC, Scape; PE, Pedicel; FL, Flagellμm; FI-FVI, Flagellomere I-VI.

表1 牛角花齿蓟马触角各节长度和宽度
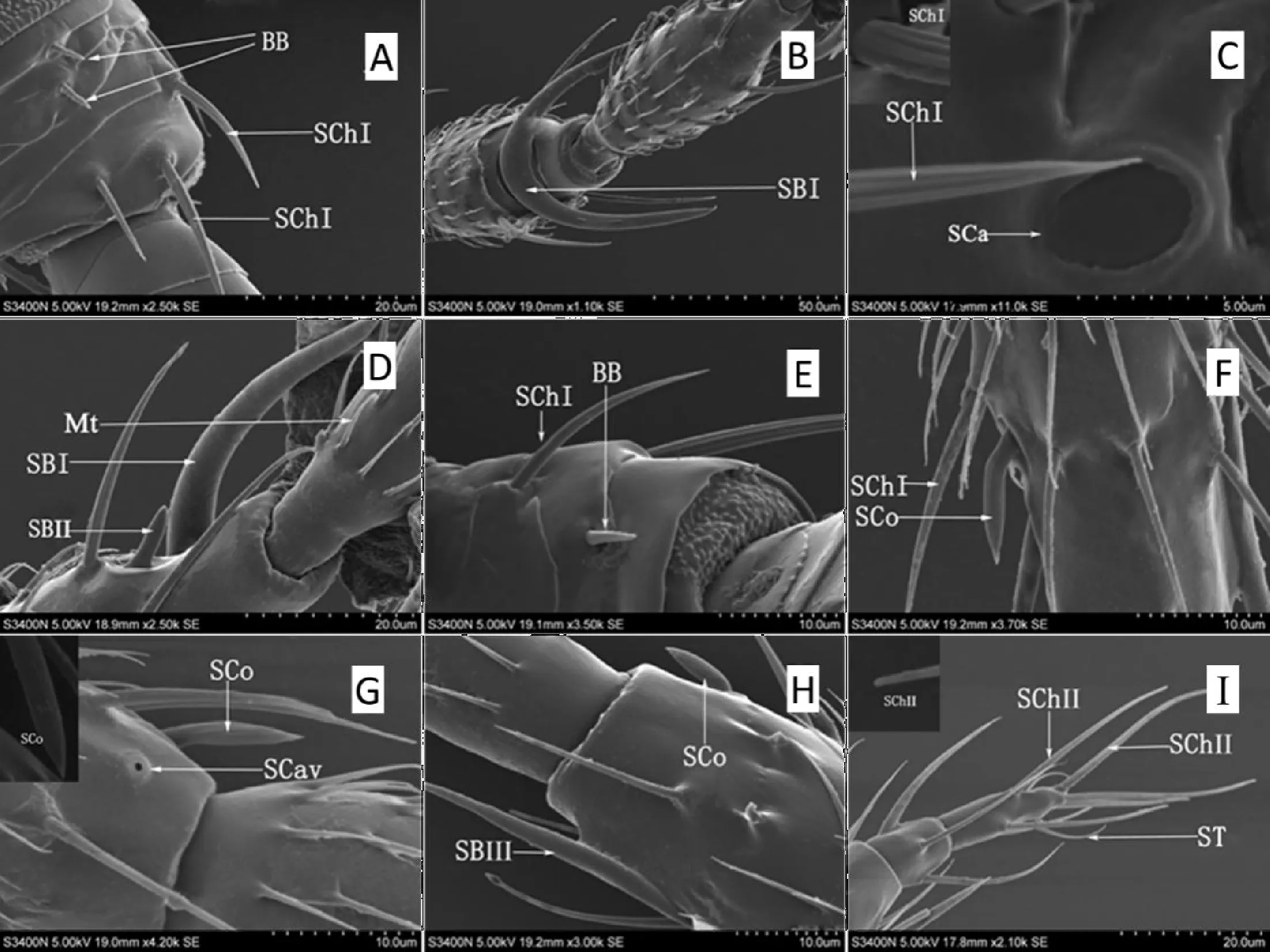
图2 牛角花齿蓟马雌虫触角感器的超微结构Fig.2 Ultrastructures of antennal sensilla in female Odontothrips loti adults注:A,柄节,Böhm氏鬃毛(BB),刺形感器I(SChI);B,第I鞭小节,锥形感器I(SBI);C,梗节,钟形感器(SCa),刺形感器I(SChI);D,第II鞭小节,微毛(Mt),锥形感器I(SBI),锥形感器II(SBII);E,柄节,Böhm氏鬃毛(BB),刺形感器I(SChI);F,第III鞭小节,腔锥形感器(SCo);G,第III鞭小节,腔形感器(SCav);H,第IV鞭小节,锥形感器III(SBIII);I,第VI鞭小节,刺形感器II(SChII),毛形感器(ST)。Note: A, scape, Böhm bristles (BB), sensilla chaetica I (SChI); B, flagellomere I, sensilla basiconca I (SBI); C, pedicel, sensilla campaniformia (SCa), sensilla chaetica I (SChI); D, flagellomere I, microtrichia (Mt), sensilla basiconca I (SBI), sensilla basiconca II (SBII); E, scape, Böhm bristles (BB), sensilla chaetica I (SChI); F, flagellomere III, sensilla coeloconica (SCo); G, flagellomere III, sensilla cavity (SCav); H, flagellomere IV, sensilla basiconca III (SBIII); I, flagellomere VI, sensilla chaetica II (SChII), sensilla trichodea (ST).

图3 牛角花齿蓟马雄虫触角感器的超微结构Fig.3 Ultrastructures of antennal sensilla in male Odontothrips loti adults注:A,柄节,Böhm氏鬃毛(BB);B,梗节,Böhm氏鬃毛(BB);C,梗节,刺形感器I(SChI),钟形感器(SCa);D,第II鞭小节,锥形感器I(SBI),刺形感器I(SChI),锥形感器II(SBII);E,第III鞭小节,微毛(Mt),刺形感器I(SChI);F,第II鞭小节,锥形感器II(SBII);G,第IV鞭小节,腔锥形感器(SCo),锥形感器III(SBIII);H,第V鞭小节,锥形感器III(SBIII);I,第VI鞭小节,毛形感器(ST),刺形感器II(SChII)。Note: A, scape, Böhm bristles (BB); B, pedicel, Böhm bristles (BB); C, pedicel, sensilla chaetica I (SChI), sensilla campaniformia (SCa); D, flagellomere II, sensilla basiconca I (SBI), sensilla chaetica I (SChI), sensilla basiconca II (SBII); E, flagellomere III, microtrichia (Mt), sensilla chaetica I (SChI); F, flagellomere II, sensilla basiconca II (SBII); G, flagellomere IV, sensilla coeloconica (SCo), sensilla basiconca III (SBIII); H, flagellomere V, sensilla basiconca III (SBIII); I, flagellomere VI, sensilla trichodea (ST), sensilla chaetica II (SChII).
2.2 触角感器类型
牛角花齿蓟马雌、雄虫触角各节着生感器类型和数量存在差异,柄节感器数量最少且类型单一,其次是梗节,鞭节感器数量分布最多,类型多样(图1)。牛角花齿蓟马雌、雄虫触角表面共观察到7种感器,分别是Böhm氏鬃毛、刺形感器、锥形感器、钟形感器、腔锥形感器、腔形感器和毛形感器,以及微毛(图2,图3)。雌虫触角具有以上所述的7种类型感器,而雄虫触角没有发现腔形感器。
2.2.1Böhm氏鬃毛
Böhm氏鬃毛存在于雌、雄虫触角柄节基部、以及梗节与柄节的节间,基部表面稍凹陷,毛体短小,端部圆钝,表面光滑、无孔,与触角表面角度呈近90°夹角。雌虫氏鬃毛长4.44±0.08 μm,直径0.89±0.03 μm;雄虫长2.71±0.71 μm,直径0.78±0.01 μm(图2-A;图3-A,B;表2)。
2.2.2刺形感器
刺形感器形如带刺植物表皮坚刺,基部较端部粗,生于表皮略突起的基节窝中,在雌、雄虫触角柄节、梗节和鞭节均有分布,根据形态特点分为两个亚型SChI和SChII。
SChI基部着生于丘状基座中,端部尖细,体壁具有明显的棱形纵脊。零散分布,主要分布在第I~V鞭小节,其次是梗节,柄节数量最少,且鞭节SChI毛体最粗大,柄节最小。雌虫较雄虫稍长,雌虫长度为19.38±1.82 μm,直径1.58±0.05 μm;雄虫长度为13.93±1.66 μm,直径1.27±0.14 μm(图2-A,C,E,F;图3-C,D,E;表2)。
SChII仅分布在雌、雄虫的第V、VI鞭小节,形体较SChI更粗大,从基部到端部逐渐变细,端部钝圆,尾部略弯曲。雌虫感器也较雄虫稍长,雌虫长度为30.62±1.46 μm,直径1.88±0.09 μm;雄虫长24.33±1.66 μm,直径1.32±0.04 μm(图2-I;图3-I;表2)。
2.2.3锥形感器
锥形感器体中空,表面多孔、布有纵纹。根据形态特征及分布部位分为3种亚型,即SBI、SBII和SBIII。
SBI感器形似角形或叉状,是感器中最粗大的一类,分布在雌、雄虫第I、II鞭小节端部的节间位置,每根触角2根感器。感器基部粗壮,着生在凹陷的基节窝中,向臂端部逐渐变细。雌虫单臂长51.9±1.24 μm,直径7.9±0.07 μm;雄虫单臂长34.63±1.58 μm,直径4.81±0.17 μm(图2-B,D;图3-D;表2)。
SBII主要分布在第I、II鞭小节侧面,基部底座较宽大,中出感器形似手指,体壁被纵纹,端部圆钝,雌虫长10.65±0.69 μm,直径2.2±0.17 μm;雄虫长8.41±0.35 μm,直径1.77±0.05 μm(图2-D;图3-D,F)。
SBIII分布在第III~V鞭小节侧面,是最长的锥形感器,毛体基部较粗,从触角表面直接长出,体壁布有纵纹,端部钝圆。雌虫长45.02±0.96 μm,直径5.11±0.06 μm;雄虫长32.01±1.33 μm,直径3.99±0.04 μm(图2-H;图3-G,H;表2)。
2.2.4钟形感器
钟形感器仅在触角梗节端部背面分布,触角表皮隆起形成不规则环状结构,环体厚实,内部表皮光滑略凹陷。雌虫感器内直径4.15±0.09 μm,雄虫3.95±0.06 μm(图2-C;图3-C;表2)。
2.2.5腔锥形感器
腔锥形感器分布在雌、雄虫第III、IV鞭小节侧面,数量少,形如萌发的嫩芽。基部着生在较大基节窝中,基部略细,表面光滑,从基部1/3处开始逐渐变粗,锥体具纵纹,直至距端部1/5处槽纹消失,端部收敛呈锥状。雌虫长8.32±0.19 μm,基部直径2.06±0.16 μm;雄虫长6.72±0.09 μm,基部直径1.93±0.03 μm(图2-F,G,H;图3-G;表2)。
2.2.6腔形感器
腔形感器仅分布在雌虫第I、III鞭小节腹面,基部呈丘状,中间表皮光滑,凹陷形成空腔,周围伴生刺形感器。感器底座直径2.03±0.03 μm,空腔直径0.75±0.02 μm(图2-G;表2)。
2.2.7毛形感器
毛形感器仅分布在第VI鞭小节上,毛体较长,表面光滑,末端较细。雌虫长19.94±0.73 μm,直径1.68±0.04 μm;雄虫长18.5±0.54 μm,直径1.24±0.03 μm(图2-I;图3-I;表2)。
2.2.8微毛
微毛是触角表皮的衍生物,毛体光滑,基部无基窝,毛体从触角表面长出,呈45°夹角,端部尖锐,略弯曲。主要分布在第I~IV鞭小节上,梗节也有少量分布,尤其是第I鞭小节分布最密集,6圈微毛环绕而生。雌虫长4.31±0.24 μm,直径0.59±0.05 μm;雄虫长7.98±1.34 μm,直径0.60±0.03 μm(图2-D;图3-E;表2)。
3 结论与讨论
通过扫描电镜观察,发现牛角花齿蓟马雌、雄成虫触角形态相似,但雌虫触角比雄虫略长,除梗节无明显差异外,其它各亚节雌虫均较雄虫粗长。雌、雄虫触角共观察鉴定到7种类型感器,即Böhm氏鬃毛、刺形感器、锥形感器、钟形感器、腔锥形感器、腔形感器、毛形感器,以及一类表皮衍生物——微毛。其中刺形感器和锥形感器数量较多,其它几种感器数量较少,表明牛角花齿蓟感器类型多样。
Böhm氏鬃毛体小,在昆虫触角上数量较少,主要是单个或成对分布在柄节基部和梗节与柄节相接的节间,在鞘翅目(徐伟等, 2019)、鳞翅目(狄贵秋等, 2020)、双翅目(袁珂等, 2020)和缨翅目(Zhuetal., 2017)等昆虫中,表现出分布的保守性。丁艳红等(2010)将西花蓟马的Böhm氏鬃毛定义成角锥形感器,但多数学者根据Schneider(1964)的方法将其划分成独立的感器类型之一,如美洲棘蓟马(Huetal., 2020)、茶黄蓟马(Zhuetal., 2017)和茶棍蓟马(吕召云等, 2015)。研究表明Böhm氏鬃毛可通过本体感受机械刺激,减缓触角下沉的重力作用,调节触角的运动状态和位置(Schneider, 1964; Krishnanetal., 2012)。牛角花齿蓟马触角Böhm氏鬃毛的形态和分布特点与以上昆虫描述类似,可能具有感受机械刺激的功能。
牛角花齿蓟马触角刺形感器有SchI和SChII两种类型,SchI稍短,略弯曲,末端尖锐,除在柄节和梗节零星分布外,主要分布于鞭节I~V侧面。SChII较SchI长,直立,末端圆钝,仅分布于第VI鞭小节。这两种类型感器与西花蓟马、花蓟马、禾花蓟马、美洲棘蓟马、茶黄蓟马和茶棍蓟马的刺形感器形态特征和分布特点基本相同。另外,除丁艳红等(2010)将西花蓟马的SchI和SChII分别定义成剑状刺形感器和长锥形感器外,以上6种蓟马均鉴定为刺形感器SchI和SChII。Slifer和Sekhon(1974)曾将刺形感器定义为“触毛”(tactile hair),推测其具有感受机械刺激的作用。透射电镜发现造瘿蓟马Kladothripsintermedius(deFaccietal., 2011)和茶黄蓟马(Huetal., 2020)的刺形感器SchI末端均无端孔,内部也无神经元树突,但SChII末端均有一端孔,并且内部含有神经元树突结构。同样,Isidoro等(1998)发现油菜蓝跳甲Psylliodeschrysocephala长刺形感器内有5~6个神经元,其中一个神经元树突与端孔相连,电生理试验也证实化学感受细胞对油菜挥发物反应敏感。这表明鞭节侧面的刺形感器SchI,特别是柄节和梗节上的SchI可能与感知机械刺激有关,而鞭节末端的较长SChII能更易感受环境刺激,具有感知机械和化学刺激的双重作用。虽然本次试验未对牛角花齿蓟马触角感器进行透射电镜解剖观察,但该蓟马刺形感器SchI和SChII也很可能分别具有感知外界机械刺激和感受化学刺激的功能。
锥形感器的表面多孔、具槽纹的结构是薄壁嗅觉感器的典型特征,通常被认为可以感受寄主释放的挥发物(Lopesetal., 2002)。牛角花齿蓟马的SBI感器锥体粗大,在第I、II鞭小节各分布一个,这类感器在蓟马触角上的分布数量和位置较保守(涂洪涛等, 2011; 李维娜和冯纪年, 2013; Zhuetal., 2017; Wangetal., 2019)。透射电镜发现茶黄蓟马的3种锥形感器SBI、SBII和SBIII的锥腔内均含有数量较多的神经元树突,这是感器执行嗅觉功能的重要依据(Zhuetal., 2017)。因此,推测分布于牛角花齿蓟马雌、雄虫触角鞭节的锥形感器也可能参与到感知植物挥发物的嗅觉过程中。
钟形感器在不同昆虫触角中的分布位置和功能存在差异,通常认为着生于梗节上的属于本体机械类感受器,可感知环境温度变化(Mclver, 1975; Skordosetal., 2002),而分布在鞭节上的是味觉感受器(Must, 2006; Ahmedetal., 2013)。缨翅目昆虫钟形感器Sca的分布较保守,在茶棍蓟马(吕召云等, 2015)、花蓟马(李维娜和冯纪年, 2013)和茶黄蓟马(Zhuetal., 2017)中,此类感器均着生在梗节上,鞭节上未发现。因此,分布在牛角花齿蓟马触角梗节上的钟形感器很可能具有感知应力机械变化和环境温度变化的双重功能。
许多昆虫的腔锥形感器表面具纵纹,有微孔,内部有双层膜结构的典型特征(Altner and Loftus, 1985; Hunger and Steinbrecht, 1998; Zhuetal., 2017),这类感器在缨翅目昆虫触角上的分布位置是一致的,均在第III、IV鞭节上(deFaccietal., 2011; Zhuetal., 2017; Huetal., 2020)。透射电镜发现茶黄蓟马的腔锥形感器内部有多个不分枝的神经元树突,每个树突周围淋巴液中含有较多嗅觉受体蛋白,类似的结构在其它昆虫触角中也被发现(MacKayetal., 2014; Zhuetal., 2017),而且这类感器可以感知寄主植物释放的挥发物组分(Pophofetal., 2005),所以牛角花齿蓟马的腔锥形感器也可能执行识别寄主挥发物的功能。
牛角花齿蓟马雌虫触角发现腔形感器,但雄虫触角未观察到,这与茶棍蓟马的此类感器在雌虫触角特异性分布类似(吕召云等, 2015)。牛角花齿蓟马腔形感器分布于触角第I、III鞭小节,周围伴生刺形感器,与锥形感器SBIII和腔锥形感器相近,这符合温湿度感器和嗅觉感器共分布的特点(Alter and Prillinger, 1980),推测此类感器的作用与感受环境温湿度变化相关。
毛形感器在牛角花齿蓟马触角上分布数量极少,仅分布在VI鞭小节。茶黄蓟马触角上的此类感器表面光滑、无孔,推测是一类机械性刺激感受器(Zhuetal., 2017)。
微毛在牛角花齿蓟马触角上分布数量较多,环状分布于第I~IV鞭节上。透射电镜发现微毛内部并无神经元树突结构,表明其不参与嗅觉过程(Zhuetal., 2017)。
本研究利用扫描电镜对苜蓿重要害虫——牛角花齿蓟马雌、雄虫触角感器进行了超微形态观察,比较了触角结构、感器类型以及分布特点在雌、雄种间的差异,并分析了不同感器可能具有的功能。研究结果为进一步研究此虫的化学生态学提供基础资料,但本研究仅明确了蓟马触角扫描电镜的形态特征,在后续工作中需结合透射电镜、单细胞记录仪等技术深入探究触角感器的功能。
