基因编辑的“前世今生”
2022-06-01朱学明鲍坚东王教瑜林福呈
李 琳,朱学明,鲍坚东,王教瑜,林福呈,2,*
(1.浙江省农业科学院 植物保护与微生物研究所,省部共建农产品质量安全危害因子与风险防控国家重点实验室,浙江 杭州 310021; 2.浙江大学 农业与生物技术学院,水稻生物学国家重点实验室,浙江 杭州 310058)
“我们从哪里来?要到哪里去?”是关于生命本源的哲学追问,生命科学的发展让人们有了找到回答这一问题的可能。长期以来,人们从未停止对生命的探索。1865年,孟德尔提出的基因分离定律和基因自由组合定律,揭示了遗传最基本的规律。1909年,摩尔根发现遗传连锁定律,更加丰富了人们对遗传规律的认识。Mccarty等通过实验证实有活性的遗传物质是脱氧核糖核酸(deoxyribonucleic acid,DNA);1952年,Hershey等通过放射性同位素标记的T2噬菌体增殖试验,进一步验证了DNA是生物的遗传物质;1953年,沃森和克里克提出的DNA分子双螺旋结构模式,使人们对生命的认识进入到了分子水平。此时,人们已经充分认识到DNA对生物性状的决定性作用,DNA序列的改变,如碱基缺失、替换、插入等可能会引起表型的变化,或引发疾病。生物在自然情况下突变的概率极低,通过人为改造的方法可以提高突变率。因此,科学家提出是否可以人为改变或修饰DNA序列中特定位置的碱基以达到改良性状或治疗疾病的目的。基于上述认识,早期人们通过同源重组来提高突变率,经过不断的探索和努力,又发现了归巢核酸内切酶(meganuclease)和锌指核酸酶(zinc finger nucleases,ZFNs)等,这些技术能人为地改变基因,但仍然存在很多弊端和局限。直到CRISPR技术的出现使基因编辑技术进入了一个崭新的发展阶段。本文拟通过对基因编辑技术的发展历程、技术原理和应用进行综述,针对该技术目前存在的问题与面临的挑战进行展望,为人们科学认识并利用该技术奠定基础。
1 基因编辑技术的演进
1.1 同源重组的发现
20世纪70年代,科学家在研究细菌如何防御噬菌体过程中发现,限制性内切酶可以保护细菌免受噬菌体的侵害,这个发现具有里程碑式的意义,科学家发现可以对基因组进行编辑。20世纪80年代,Smithies等和Capecchi发现,可以通过同源重组将外源DNA整合到哺乳动物的基因组中;但这种方法有很大的局限性,不仅整合效率极低(整合效率取决于细胞的状态和类型),而且容易脱靶。
1.2 归巢核酸内切酶
20世纪80年代末期,Rudin等和Rouet等发现,在靶标位置引入双链断裂的DNA(double-strand break,DSB)会显著提高目的基因的整合效率。研究人员最早通过归巢核酸内切酶(也称大范围核酸内切酶)在基因组中引入特定的双链断裂DNA。归巢核酸内切酶能够识别14~40 bp的DNA片段,识别之后绝大部分通过非同源末端连接修复(non-homologous end joining,NHEJ)(图1-A和图1-D)。人们在自然界中已经找到数百种归巢核酸内切酶,使用最广泛的主要有3种:Ⅰ-Ⅰ[Plessis等在啤酒酵母()的线粒体中发现]、Ⅰ-Ⅰ[Dürrenberger等在衣藻()的叶绿体中发现]和Ⅰ-Ⅰ[Silva等在超嗜热古菌()中发现]。然而,通过归巢核酸内切酶进行基因编辑存在很大的制约性:(1)尽管有数百种的归巢核酸内切酶,每个都有自己独特的识别序列,但要找到适合靶向特定基因序列的归巢核酸内切酶的概率极低;(2)由于DNA双链断裂主要通过非同源末端连接修复,该修复机制不能引入外源的DNA模板,并且会在断裂位点随机删除或者插入DNA片段。
1.3 锌指核酸酶
Klug等发现,锌指蛋白能够特异性识别3 bp的DNA序列,多个锌指蛋白可以组装成大复合物。Kim等发现,限制性核酸内切酶Ⅰ具有独特的DNA识别域和切割域,在靶标位点进行同源二聚化以切割目的DNA片段。研究人员将Ⅰ的DNA识别域去除,将其切割域与锌指模块融合,命名为锌指核酸酶。锌指核酸酶技术被称为第一代基因编辑技术,利用该技术进行基因编辑时,需要设计2个单独的锌指模块(分别结合DNA的2条链),针对DNA序列上相邻的2个位点(5-7 bp),通过Ⅰ同源二聚化导致DNA靶标位点的DNA双链断裂,DNA损伤后通过非同源末端连接修复或同源定向修复(homology directed repair,HDR),从而达到基因编辑的目的(图1-B和图1-D)。此外,还可以通过加工改造锌指模块,靶向不同的DNA序列进行特异性切割。然而该技术也有局限性:(1)可编辑的靶基因位点有限,有限的锌指蛋白类别只可识别有限的DNA序列;(2)ZFNs基因编辑的效率低,只有30%左右;(3)操作复杂,成本高,容易脱靶。科研人员一直在不断改进ZFNs技术,Miller等和Doyon等发现,可以通过电荷之间的相互排斥形成异二聚体的ZFN结构,从而阻止Ⅰ在非靶标位置进行二聚化并切割DNA序列。Guo等发现,通过蛋白质工程的方法可以提高Ⅰ的切割效率。2014年,Tebas等通过ZFNs编辑艾滋病毒(human immunodeficiency virus,HIV)感染者自体CD4 T细胞中的艾滋病毒入侵辅助受体CCR5来治疗艾滋病。
1.4 类转录激活因子效应物核酸酶
类转录激活因子(transcription activator-like effector,TALE)是植物病原菌黄单孢菌中的一种效应因子。2009年,Boch等和Moscou等发现,类转录激活因子蛋白可以识别DNA序列。2011年,第二代基因编辑技术——类转录激活因子效应物核酸酶TALENs诞生。与ZFNs一样,TALENs在形式和功能上也是模块化的,由Ⅰ切割结构域和TALE蛋白DNA结合结构域融合组成。TALENs识别模块一般由34个氨基酸组成,其中第12和13位的氨基酸称为可变双氨基酸残基(repeat variable diresidues,RVDs),RVDs特异性识别DNA上核苷酸的种类。RVDs的识别密码为NI(Asn Ile)识别腺嘌呤(A),HD(His Asp)识别胞嘧啶(C),NN(AsnAsn)识别鸟嘌呤(G),NG(AsnGly)识别胸腺嘧啶(T)。TALENs的DNA结合域通常由1.5~33.5个重复的TALEN单体组成。如需进行基因编辑,如靶基因的敲除,将1对TALENs共转入细胞后,2个Ⅰ形成二聚体后在靶标位点剪切DNA,使DNA双链断裂,细胞通过非同源末端连接或同源定向修复DNA(图1-C和图1-D),在修复过程中插入或删除部分碱基,造成移码突变,导致下游一系列密码子改变,相当于敲除目的基因。在实际操作中,Mussolino等发现,1对TALFNs的识别位点要间隔17个碱基左右。与ZFNs相比,TALENs技术具有设计简单、特异性高的优势,是科研人员研究基因功能和潜在基因治疗的工具;但该技术仍存在模块组装过程繁琐、具有一定的细胞毒性等缺点。
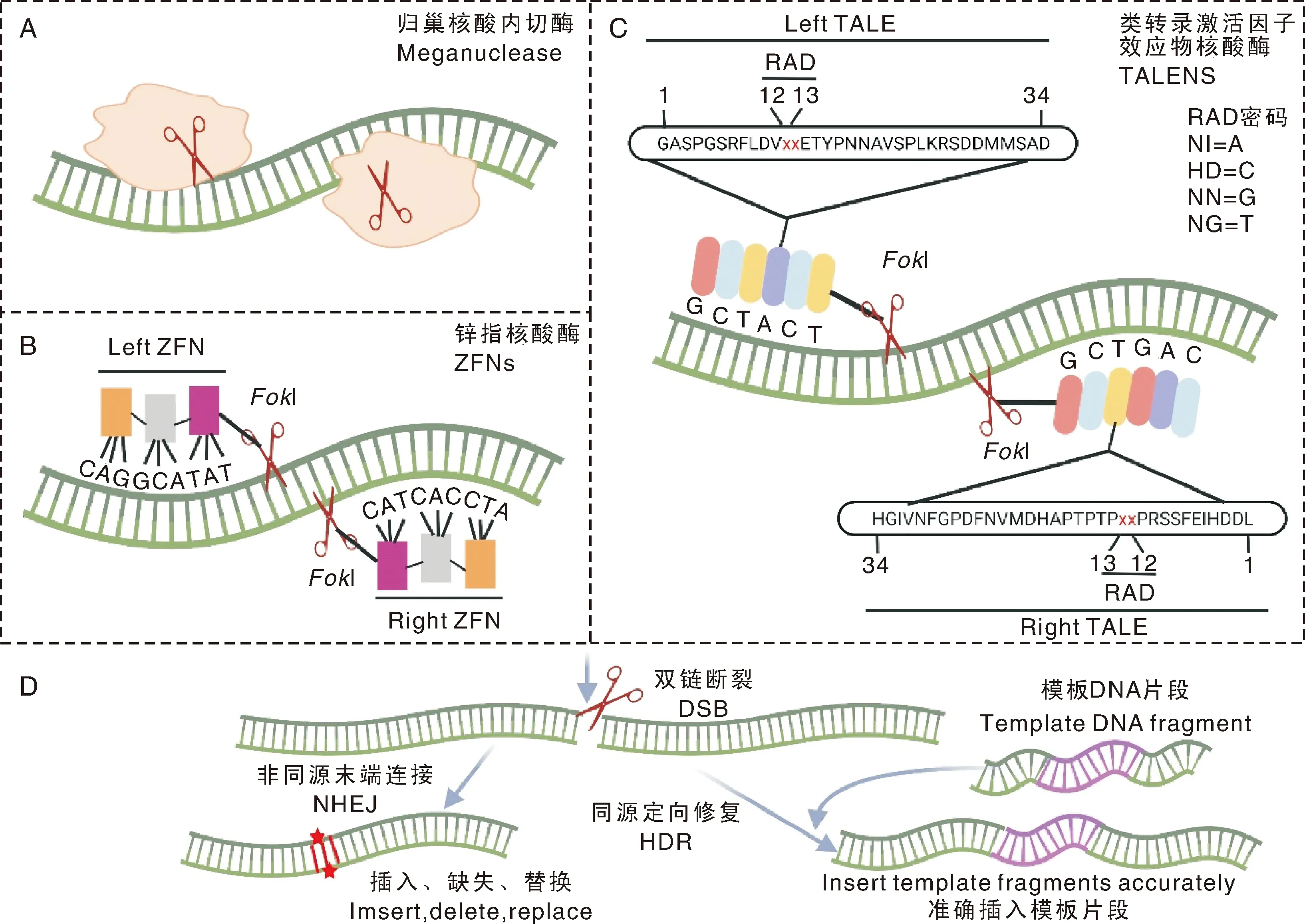
A,归巢核酸内切酶可识别长段DNA序列,没有明显的结合和切割结构域;B,锌指核酸酶,每个锌指核酸酶识别3个碱基。C,类转录激活因子效应物核酸酶,每个TALE识别单个碱基;D,以上3种酶都会导致DNA双链断裂,可以通过容易出错的非同源末端连接(NHEJ)或同源定向修复(HDR)来修复。NHEJ会导致目标位点的随机插入缺失和基因破坏,HDR可以在目标位点插入特定的DNA模板(单链或双链)以进行精确的基因编辑。
1.5 CRISPR-Cas系统
ZFNs和TALENs技术提高了基因敲除效率,但是针对基因组中的不同位点都需要重新设计新的蛋白质,操作繁琐,技术门槛高,限制了应用和推广。经过科学家的不懈努力, CRISPER技术应运而生,它的出现彻底弥补了以上各种基因编辑技术的缺陷,因其系统简单、精准、快速,极大降低了技术门槛,一出现就风靡整个生物界。CRISPR(clustered regularly interspaced shortpalindromic repeat,成簇的规律间隔的短回文重复序列)原本只是原核生物中一种比较特殊的DNA重复元件。1987年,Ishino等最早在大肠埃希菌中发现CRISPR,当时还没有将这些重复的序列命名。2002年,Jansen等将这些重复的序列正式被命名为CRISPR。CRISPR重复序列簇中间隔着非重复DNA序列,非重复DNA序列间隔区来自于病毒或者其他可移动的遗传元件,并且CRISPR与保守的Cas(CRISPR-associated)基因相邻。Barrangou等发现,在病毒攻击嗜热链球菌后,嗜热链球菌会将噬菌体基因组序列整合到新间隔区,这段新间隔区决定了Cas酶的靶向特异性,铸建了对噬菌体的防御系统。
PAMs(protospacer-adjacent motifs,原始间隔区相邻基序)的发现为CRISPR的发展奠定了坚实的基础。Deveau等发现,CRISPR的间隔序列与PAMs高度相似,并且PAMs对CRISPR系统的运行极其重要。随后Garneau等发现,Cas9是嗜热链球菌中唯一具有DNA催化活性的蛋白。Jinek等和Gasiunas等发现,嗜热链球菌的CRISPR基因座能够重建对大肠埃希菌的干扰,此外Cas9酶可以重新编程以靶向细菌中目的DNA序列,该发现是CRISPR作为一种生物技术工具的标志。Cas9发挥作用时需要2种短RNA,即crRNA(CRISPR RNA)和tracrRNA(trans-activating crRNA)。成熟的crRNA和反式激活的crRNA的发现简化了CRISPR的操作。crRNA由作为引导序列的部分和与tracrRNA碱基配对的另一部分组成。crRNA、tracrRNA和Cas9形成Cas9-RNA复合物在靶标位置通过DNA双链断裂切割DNA。sgRNA(single guide RNA)的发现进一步简化了该技术,Jinek等发现,CRISPR-Cas9可以被sgRNA引导,sgRNA由tracrRNA和crRNA融合形成(图2)。至此以后,研究人员只需设计一个简单的sgRNA,就可以轻松引导CRISPR-Cas9识别基因组中的准确位置并进行基因编辑,CRISPR-Cas9技术被视为第三代基因编辑技术。
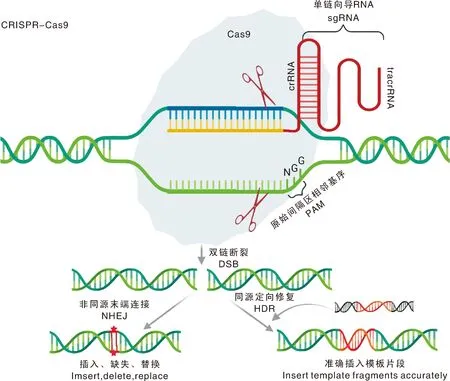
Cas9核酸酶靶向切割与单链向导RNA (sgRNA)互补的DNA序列,该序列位于原始间隔区相邻基序(PAM)的上游。
Koonin等从广义上将CRISPR分为2大类,第一类包括在古细菌中发现的Ⅰ型和Ⅲ型CRISPR系统;第二类包括Ⅱ、Ⅳ、Ⅴ和Ⅵ型CRISPR系统。目前使用最广泛的是来自酿脓链球菌()的Ⅱ型CRISPR-Cas9系统,因为其NGGPAM序列很简单。不过研究人员仍然在积极探索不同的CRISPR系统,主要侧重于3个方面:(1)减少Cas9核酸酶的分子量;(2)提高其保真性;(3)扩大Cas9的靶向范围。研究人员在现有的基础上,发明了第二代CRISPR基因编辑技术。第二代技术可以精准地将单个碱基转变为想要的碱基(将碱基直接变成T、A或者G),并且不会引起DNA的双链断裂。目前主要有胞嘧啶剪辑编辑器(cytidine base editing,CBE)和腺嘌呤碱基编辑器(adenine base editing,ABE)这两类DNA碱基编辑器。CBE是将C-G碱基对转换为T-A碱基对,ABE将A-T碱基对转换为G-C碱基对。这些新颖的单碱基编辑方法扩大了基因组靶向的范围。dCas9蛋白是Cas9蛋白的突变体,Chen等利用dCas9调节基因表达。dCas9技术的进步大大提高了活细胞成像的效率和活细胞染色质成像的基因组靶向范围。
除了Cas9蛋白,Cas12、Cas13、Cas14蛋白也陆续被发现。Teng等发现,Cas12擅长识别双链DNA,在识别DNA序列中的单碱基时,发现有匹配不好的碱基,会继续寻找直至识别到正确的序列,然后结合形成封闭的R-环。Cox等发现,Cas13擅长识别单链RNA,即靶向RNA进行基因编辑。Cas13专注靶向RNA的功能补充了靶向DNA的CRISPR-Cas9系统。目前已经发现Cas13a、Cas13b、Cas13c和Cas13d这4种亚型。各种亚型中,Cas13d具有蛋白体积最小、载体构建方便、易于向细胞内输送等明显优势。Harrington等发现,Cas14擅长识别单链DNA,剪切时不依赖PAM序列,它比Cas9蛋白小,是迄今为止发现的最小的功能性CRISPR系统。
2 基因编辑的应用
2.1 基因编辑在农业生产上的应用
(2)将航拍影响拼接为正射影响,该影像为geotiff格式,包含rgba三个通道;将LiDAR数据提取为DEM,并以单通道的geotiff格式保存。
2.1.1 提高产量品质
提高农作物的产量是育种最重要的目的之一。在水稻育种方面,Wang等利用CRISPR技术获得了株高降低且有增产潜力的水稻材料。Lv等利用CRISPR技术敲除水稻中5基因后,水稻籽粒显著增多,产量显著提高。Miao等研究表明,通过靶向水稻中一个调节粒型和株高的基因396,使之突变,突变体植株的籽粒变大、穗长增加,且在低氮条件下表现出高产稳产。在小麦育种方面,Wang等利用CRISPR技术敲除小麦籽粒重基因2后,突变株的籽粒大小和千粒重明显提高。为保证水稻的蒸煮品质,Zeng等、Xu等和Huang等利用基因编辑技术,通过靶向调控基因,培育出口感优良的水稻品种。香稻品种的米饭有独特香味和口感,其香味和口感受2基因的调节,Ashokkumar等和Tang等利用CRISPR技术编辑该基因后增强了米饭的香味。
2.1.2 增强抗性
植物病虫害等生物胁迫对粮食安全造成了严重的威胁。Wang等利用基因编辑技术敲除调控稻瘟病菌的抗性基因922后,培育出抗稻瘟病的纯合突变株系。Oliva等通过编辑基因启动子区域培育出对百叶枯病具有广谱抗性的水稻品种。Wang等通过TALENs和CRISPR-Cas9技术修饰六倍体小麦中MLO蛋白的多个等位基因后,小麦对白粉病的抗性增强。
干旱、高温、寒冷、重金属污染等非生物胁迫对作物生长发育和产量有严重影响,通过基因编辑技术增强农作物对非生物胁迫的耐受性是亟待解决的问题。镉是一种对人体有害的重金属,镉超标的大米是饮食上镉摄入的主要来源。Clemens等、Ishikawa等和Tang等通过基因编辑镉转运相关蛋白,筛选出耐镉的水稻品种。Shi等通过CRISPR技术编辑玉米中8基因,增强玉米对干旱的抗性。Zhang等通过改良的基因编辑技术敲除水稻相关调控基因166,增强水稻的抗旱性。
2.1.3 基因编辑作物的监管现状
2.2 基因编辑在医学上的应用
2.2.1 药物靶标基因的筛选
基于全基因组的CRISPR技术敲除筛选可用于功能基因组学研究,通过该技术能够检测细胞耐药性的基因组位点,明确细胞如何诱导宿主免疫反应,阐明某些病毒如何诱导细胞死亡。利用基因编辑技术发现的功能性非元件为研究人类基因组的结构和进化、药物靶标的筛选提供了一种新的手段。
2.2.2 疾病建模和生产器官供体
基因编辑技术的成熟加速了转基因动物的出现,如转基因斑马鱼、转基因小鼠、转基因大鼠、转基因猴子和转基因猪,这些转基因生物加速了人类疾病的建模,为发现新的治疗方法奠定基础。
CRISPR技术还可以解决外来移植器官的免疫排斥反应。猪被认为是人外来器官的首选动物。2020年12月,美国食品药品管理局批准了敲除-半乳糖的转基因猪GalSafe上市,它既可以为半乳糖过敏者提供安全肉类食物,又可以提取肝素等生产药物。2021年9月25日,美国研究人员将一头转基因猪的肾移植入脑死亡的志愿者体内,移植后肾脏工作了54 h,未见身体出现排斥反应,这一“变革时刻”的医学进步可能为成千上万需要器官移植的患者带来希望;该项研究的论文还没有经过同行评议正式发表,并且此项研究只是获得初期的成功,移植后的1个月乃至10 a后会发生什么还是未知数。
2.2.3 基因治疗
基因编辑可以精准改造人类基因,在治疗遗传性疾病方面有巨大潜力。目前来说,比较成功的案例是Tebas等利用ZFNs技术破坏HIV共受体CCR5,已经在临床Ⅰ期试验成功。Tabebordbar等和Nelson等利用基因编辑技术恢复了肌营养不良蛋白基因的表达,拯救了杜氏肌营养不良症模型小鼠的肌肉功能。在模型小鼠中,Yin等利用基因编辑技术治疗人类遗传性酪氨酸血症。最近,Stadtmauer等研究表明,CRISPR编辑细胞在单次注射后可以长时间保持抗肿瘤功能。2021年6月26日,首个人体基因编辑临床实验数据刊登在新英格兰医学杂志上,Gillmore等利用CRISPR-Cas9剔除编码甲状腺素运载蛋白淀粉样变性的基因,这将开启一针治疗人类遗传病的时代。CRISPR技术可以靶向人类病毒如HIV-1、疱疹病毒、乙型肝炎病毒等。Hu等利用CRISPR特异性识别宿主细胞,对病毒相关基因进行精准编辑,从而抑制病毒的表达和复制。
2.2.4 人类胚胎基因编辑
人类胚胎的基因编辑工作一直是舆论关注和争论的焦点。2015年4月,Liang等利用CRISPR-Cas9技术同源修复了人类胚胎中引起地中海贫血基因的突变。2016年,Kang等利用CRISPR编辑受精卵中的5,结果由于脱靶产生了嵌合式的受精卵。
目前关于人类胚胎基因编辑存在较多问题和争论:(1)安全性问题,目前人们对胚胎编辑的安全性无法做出有效评估,CRISPR技术存在脱靶效应的报道屡见不鲜。Haapaniemi等研究认为,CRISPR编辑可能增加患癌症的风险。(2)权利问题,任何人是否有权利决定采取胚胎基因编辑技术对患者进行治疗;此外,如何平衡个人与整体利益也亟待解决。(3)公平问题,如果基因编辑技术已经很成功,谁可以享受该技术?
2.3 基因编辑在工业生产上的应用
优质的微生物菌种对提高发酵工业产品的品质与产量有重要意义。基因编辑技术可以对菌种进行定向遗传改良。放线菌是工业生产次生代谢物的最重要来源之一。Tong等通过基因编辑,使放线菌中某些基因失活,从而提高代谢物的产量,提高其经济价值。目前,仅在极少数工业微生物中建立了基因编辑系统,如酿酒酵母、稻瘟病菌、产甘油假丝酵母等。因此,利用基因编辑技术开发和改造兼容性的核酸酶,并建立多种微生物成熟的应用体系,对创造工业微生物菌株有重要意义。
3 基因编辑技术面临的挑战
基因编辑技术在疾病治疗、动植物品种改良、抗病育种、医药生产、工业生产等领域取得了不同程度的进展,但是基因编辑技术本身及其应用也呈现出了复杂性和不确定性的特征,这给传统伦理学与相关伦理准则的制定带来了巨大的冲击与挑战。
3.1 基因编辑在工农业应用上的局限
基因编辑在技术上的局限性:(1)CRISPR技术中PAM序列的限制。在实际应用时,PAM序列只识别NGG,对于特定靶位点的选择存在一定限制。(2)转化系统的限制。植物中主要通过农杆菌介导、基因枪法、原生质体转化3种方式,这3种方法都有局限性。(3)脱靶现象。脱靶现象是没有找到要编辑的位置,而去随机切割替换的非目的位点,从而造成意想不到的严重后果;科研人员不断改进CRISPR技术,减少了脱靶几率,但是脱靶现象仍然存在,无法根除。(4)广泛应用问题。基因编辑作物是否能够市场化还存在较大的争议。人类可食用植物经过了选择性培育,选择方向非常明确,第一是提高营养成分含量,第二是降低原来的毒性,很多野生植物已经充分驯化,成为了主要食物。目前基因编辑的食物还未经历长时间的自然选择和试验,是否存在潜在的风险短时间难以判断,这加剧了大众对于基因编辑食品的不认可、排斥和担忧。
3.2 基因编辑在医学上的弊端
基因编辑在医学上的研究很多,但仍然是初步阶段,目前存在四大问题:(1)有效性。基因编辑成功的效率比较低,有效性不高。(2)递送的方式。CRISPR/Cas9在细胞内发挥基因编辑作用的形式分别是Cas9蛋白和sgRNA,而Cas9进入细胞的形式有3种:DNA、RNA或蛋白,sgRNA进入细胞的形式有2种:DNA或RNA,由于缺乏理想的递送系统,限制了CRISPR/Cas9技术在临床上的应用。(3)脱靶问题。sgRNA可能结合到与靶基因相似的序列上,从而在非靶标基因处切割DNA,从而造成脱靶。Cas9也可能会识别非标准的PAM,导致不同程度的脱靶效应。Cas9蛋白的持续表达也是引起脱靶现象的重要原因。如果在编辑人类细胞过程中造成脱靶现象,对于个体的影响将是致命的。(4)伦理安全问题。基因编辑技术发展到现在还不足10 a,人类从基因的角度研究生命也才60多年,我们还没有完全了解每个基因的功能,而且人是一个复杂的有机体,对人类基因功能的研究需要上升到时间、空间、环境等,甚至四维、五维的认知高度。人类的大多数疾病是由多种因素决定的,不仅仅是通过编辑一个基因就能达到治病的效果,被编辑的基因也可能潜在影响身体的其他性能。
4 展望
基因编辑技术经过近十年长足的发展,形成四大技术,极大地推动其在农业、医学、工业等领域的应用。基因编辑技术也面对巨大的风险和挑战,基因编辑能否高效靶向治疗疾病还需要长时间的研究和临床试验,技术发展背后的伦理、公平正义、文明等问题不断激增。基因编辑存在3个伦理上的困惑:(1)如何认定基因编辑有利于人类的繁衍和健康?(2)如何判别伦理上允许和不允许的界限?即什么样的基因编辑可以被允许或不被允许?(3)如果被允许的基因编辑所想要达到的目的可以通过诸如环境、后天的教育等达到同样的效果,那么允许这些基因编辑行为是否还值得提倡和发展?
要充分客观评价基因编辑技术并解决其带来的负面问题,将技术不断完善改进的同时也要加强立法监督管理。应该摸着伦理底线的石头过河,在生命伦理的框架中稳步健全地对生命进行无尽的探索,只有对个人和社会都有益时基因编辑技术才能大放异彩。在追求基因编辑技术进步的同时,也要大力发展遗传转化技术,提高转化效率。如花粉管导入技术,其能够与常规育种技术相结合,操作简单,经济适用,是一种改变作物遗传性、培育优良品种的好方法,利用花粉管导入技术将玉米的基因导入水稻中产生玉米稻,玉米稻是远缘杂交的结果,打破了生殖隔离,提高了抗性,实现了品种的高产。将基因编辑技术与花粉管导入技术相结合,或许可以达到良好的基因编辑效果。要保证多种优良技术齐头并进的发展模式,为人类和环境的健康发展做出贡献。
