葡萄碘泡虫、似葡萄碘泡虫和茄形碘泡虫的鉴别及系统发育
2022-05-27谭禄奇王苗苗杨承忠赵元莙
谭禄奇 周 杨 王苗苗 杨承忠, 赵元莙*
(1. 重庆师范大学生命科学学院, 重庆市动物生物学重点实验室, 重庆 401331; 2. 铜仁学院, 贵州省梵净山地区生物多样性保护与利用重点实验室, 铜仁 554300)
黏孢子虫是一类物种丰富、分布广泛的后生动物寄生虫, 主要寄生于鱼类, 迄今已报道的种类约有2600种[1]。早期对于黏孢子虫的分类主要依据成熟孢子的形态特征, 但因黏孢子虫个体微小、形态简单、可用于分类的形态学特征较少, 许多分类学问题无法得到解决, 形态相似种的鉴别尤其困难。近几十年来, 分子生物学的应用, 极大程度地促进了黏孢子虫分类学的发展, 解决了诸多经典分类学的遗留问题[2—6]。目前, 综合利用形态学、分子生物学和生态学等信息进行黏孢子虫分类学的研究已成为该领域学者们的共识。
葡萄碘泡虫Myxobolus acinosusNie & Li,1973、似葡萄碘泡虫Myxobolus pseudoacinosusGuo,et al., 2018和茄形碘泡虫Myxobolus toyamaiKudo, 1917均寄生于鲤科鱼类的鳃丝部, 且形态非常相似, 在病原鉴定中容易混淆。葡萄碘泡虫由倪达书和李连祥于1973年在采自湖北省黄石市花马湖的鲤Cyprinus carpio的鳃丝上检获并命名[7], 后来又有学者分别在采自湖北省孝感市的鲤、湖北省武昌市的镜鲤、浙江省灵江市的鲤及武陵山地区(湖南省吉首市、贵州省铜仁市)的鲫Carassius auratus auratus的鳃丝上获得该种[8]。似葡萄碘泡虫由Guo等[9]于2018年在采自湖北省武汉市的鲤的鳃丝上检获并命名。茄形碘泡虫由Kudo于1917年在鲤的鳃上检获并命名。据原始描述, 该种含有一个极囊及位于该极囊前端的一个长条状原生质团[10]。1933年, Kudo建立了单极虫属ThelohanellusKudo,1933, 并将茄形碘泡虫置于该属中[11]。然而, 多年来, 该物种孢子中的长条状原生质团是否为极囊一直备受争议, 因此茄形碘泡虫M.toyamai和茄形单极虫T.toyamai这两个物种名的有效性受到质疑[12—14]。2015年, Yokoyama等[15]发现上述长条状原生质团其实是一个小型极囊, 据此重新确认了物种名茄形碘泡虫M. toyamai的有效性。茄形碘泡虫分布广泛, 在苏联、美国、越南北部、欧洲、日本和中国等均有发现[8,14—16]。本实验在我国境内再次获得了葡萄碘泡虫、似葡萄碘泡虫和茄形碘泡虫, 基于形态学和18S rRNA基因信息对三者进行鉴别和分子系统学研究, 以期更深入地了解这三个物种间的进化关系, 为鱼类病原的鉴定提供基础资料。
1 材料与方法
1.1 标本采集与物种鉴定
宿主鲤分别于2019年7月、2019年9月和2020年8月采自重庆市沙坪坝区和大足区及贵州省铜仁市碧江区。在被感染的宿主鳃丝部可以观察到乳白色的圆形和椭圆形孢囊, 参考赵元莙等[17]的方法进行后续样品的处理及基于形态学的物种鉴定。
1.2 主成分分析方法
基于成熟孢子的孢子长、孢子宽、大极囊长、大极囊宽、小极囊长和小极囊宽的测量值, 利用PAST3进行主成分分析(PCA), 以分析葡萄碘泡虫、似葡萄碘泡虫和茄形碘泡虫不同株系及不同物种间的形态差异。使用变量协变矩阵生成具有95.0%置信的散点图。
1.3 DNA提取与PCR扩增
将获得的葡萄碘泡虫孢囊戳破后立即挑取部分孢子镜检, 显微拍照, 剩下的孢子用无水乙醇浸泡在离心管中保存。从装有样品的离心管底部吸取10 μL含有黏孢子虫的液体, 经超纯水清洗2—3次以除去杂质, 再用Dneasy Tissue Kit (QIAGEN, Germany)试剂盒按照厂家提供的说明书进行基因组DNA的提取。提取得到的40 μL基因组DNA溶液置于-20 ℃冰箱保存备用。用于扩增18S rDNA基因的引物为18e(5′-CTGGTTGATCCTGC CAGT-3′)[18]和18R(5′-CTACGGAAACCTTGTT ACG-3′)[19]。25 μL的PCR反应体系: 超纯水6.5 μL,模板DNA 4 μL, Mix 12.5 μL, 10 μmol/L引物各1 μL。反应条件为: 95℃预变性5min; 95℃变性90s, 58℃退火30s, 72℃延伸2min, 35个循环; 最后72℃延伸5min。PCR产物于1.5%琼脂糖凝胶电泳分离, 用DNA凝胶回收试剂盒(Gel Extraction Kit)进行纯化回收, 将回收产物插入pMD18-T载体(TaKaRa, 日本), 导入大肠杆菌进行单克隆培养, 每株系送两个克隆子至英潍捷基(上海)生物公司测序, 得到每株系测序返回的两个克隆子18S rDNA序列相同。
1.4 序列分析和系统发育
序列相似度的计算用在线序列双重比对工具(http://www.ebi.ac.uk/Tools/psa)完成。所选序列两两之间的遗传距离利用MEGA 6.0[20]选择K2P模型计算完成。变异位点分析借助Bioedit和MEGA 6.0完成。将本实验获得的4条葡萄碘泡虫18S rDNA序列、1条似葡萄碘泡虫18S rDNA序列和2条茄形碘泡虫18S rDNA序列分别在GenBank数据库中通过BLAST进行序列同源比对。根据比对结果, 选取与这7条序列同源性较高的67条序列, 加上本实验得到的7条序列, 另选取鲑两极虫Myxidium truttaeAF201374、金色楚克拉虫Zschokkella auratisKC84942作为外群构建系统发育树。利用MrBayes 3.1.2软件[21]构建BI树, 序列最佳进化模型为GTR+I+G, 共执行10000000代, 每200代抽样1次,在舍弃25%的老化样本后, 根据剩余样本构建一致树。利用在线软件CIPRES Science Gateway V. 3.1(htlp://www.phylo.org/sub_sections/portal)构建ML树, 选用模式为RAxML-HPC2 XSEDE(8.2.12)。然后用FigTree v 1.3.1和Photoshop CS3完成系统树的绘制。
1.5 18S rRNA二级结构预测
二级结构模型从欧洲核糖体RNA数据库(http://bioinformatics.Psb.ugent.Be/webtools/rRNA/s ecmodel/index.html)[22]获得。利用MEGA 6.0对所选18S rRNA基因进行比对以获得可变序列区域, 基于18S rRNA二级结构对进化趋势进行比较研究。用软件RNA Structure 5.3采用自由能最小化模型[22,23]对二级结构进行预测, 所有参数均设置为默认值。获得的二级结构与已知的真核生物18S rRNA二级结构模型进行比较, 并使用RNAViz 2.0手动调整[24,25]。
2 结果
2.1 葡萄碘泡虫、似葡萄碘泡虫和茄形碘泡虫的形态学描述
本实验共获得葡萄碘泡虫4个株系, 均寄生于鲤的鳃丝部, 其中株系1(S1)采自重庆沙坪坝区, 另3株系采自贵州铜仁碧江区(S2、S3和S4; 表 1)。4株系孢囊均呈乳白色, 圆形, 直径为0.1—0.3 mm(图 1A)。成熟孢子壳面观呈长葡萄形, 前端稍窄略弯, 后端圆钝(图 2A和2B), 孢子长(10.8±0.4) μm(9.3—11.9 μm;n=120), 孢子宽(6.2±0.4) μm (5.0—7.1 μm;n=120)。两极囊位于孢子前端, 形态和大小差异较大, 大极囊呈梨形, 长(4.6±0.5) μm (3.1—5.4 μm;n=120), 宽(2.6±0.3) μm (1.8—3.2 μm;n=120), 极丝缠绕5—6圈; 小极囊呈球棒形, 长(2.3±0.3) μm(1.7—2.9 μm;n=120), 宽(0.9±0.1) μm (0.7—1.3 μm;n=120), 极丝缠绕2—3圈。缝面观呈透镜形, 缝脊较粗(图 2C和2D)。4株系形态与《中国动物志》[8]报道株系(S5)及原始报道株系(S6)一致(表 1)。主成分分析显示, 本实验获得的葡萄碘泡虫4株系间形态学量度无显著差异(图 3)。

图1 葡萄碘泡虫孢囊(A)和茄形碘泡虫孢囊(B)Fig. 1 Cysts of M. acinosus (A) and M. toyamai (B)

表1 葡萄碘泡虫各株系形态比较Tab. 1 The morphological comparison among different strains of Myxobolus acinosus
本实验获得似葡萄碘泡虫1个株系, 采自重庆大足区, 寄生于鲤的鳃丝部, 孢囊呈乳白色, 椭圆形。似葡萄碘泡虫成熟孢子壳面观呈茄形, 前端狭窄且弯曲, 后端圆钝, 孢子长(15.9±0.5) μm (15.1.8—17.0 μm;n=30), 宽(5.5±0.5) μm (5.0—6.3 μm;n=30)。两极囊位于孢子前端, 形态和大小差异较大, 大极囊呈梨形, 长(7.7±0.5) μm (6.7—8.7μm;n=30), 宽(3.5±0.3) μm (3.0—4.1 μm;n=30), 极丝缠绕7—8圈, 小极囊呈球棒形, 长(3.2±0.2) μm (2.5—3.5 μm;n=30), 宽(0.9±0.1) μm (0.7—1.0 μm;n=30),极丝缠绕2—3圈(图 2E和F)。未观察到缝面观。本实验所得似葡萄碘泡虫株系(S1)形态与原始报道的宽型株系(S3)一致(表 2)。
本实验获得茄形碘泡虫2个株系(S1和S2), 均采自贵州铜仁碧江区, 寄生于鲤的鳃丝部, 孢囊呈乳白色, 圆形, 直径为0.2—0.5 mm(图 1B), 成熟孢子壳面观呈茄形, 前端狭窄稍弯, 后端圆钝, 孢子长(15.3±0.6) μm (14.3—16.6 μm;n=60), 宽(6.2±0.4) μm(5.0—7.0 μm;n=60)。两极囊位于孢子前端, 形态和大小差异较大, 大极囊呈梨形, 长(5.6±0.4) μm(4.6—6.4 μm;n=60), 宽(2.9±0.2) μm (2.4—3.4 μm;n=60), 极丝缠绕6—8圈。小极囊呈球棒形, 长(3.0±0.3) μm (2.2—3.7 μm;n=60), 宽(0.9±0.1) μm(0.6—1.2 μm;n=60), 极丝缠绕2—3圈(图 2G和2H)。孢子缝面观似梨形, 前端狭窄弯曲, 后端圆钝(图 2I和2J)。本研究所得茄形碘泡虫形态与原始报道株系一致(表 2)。主成分分析显示, 本实验获得的茄形碘泡虫2株系间形态量度无显著差异(图 3)。
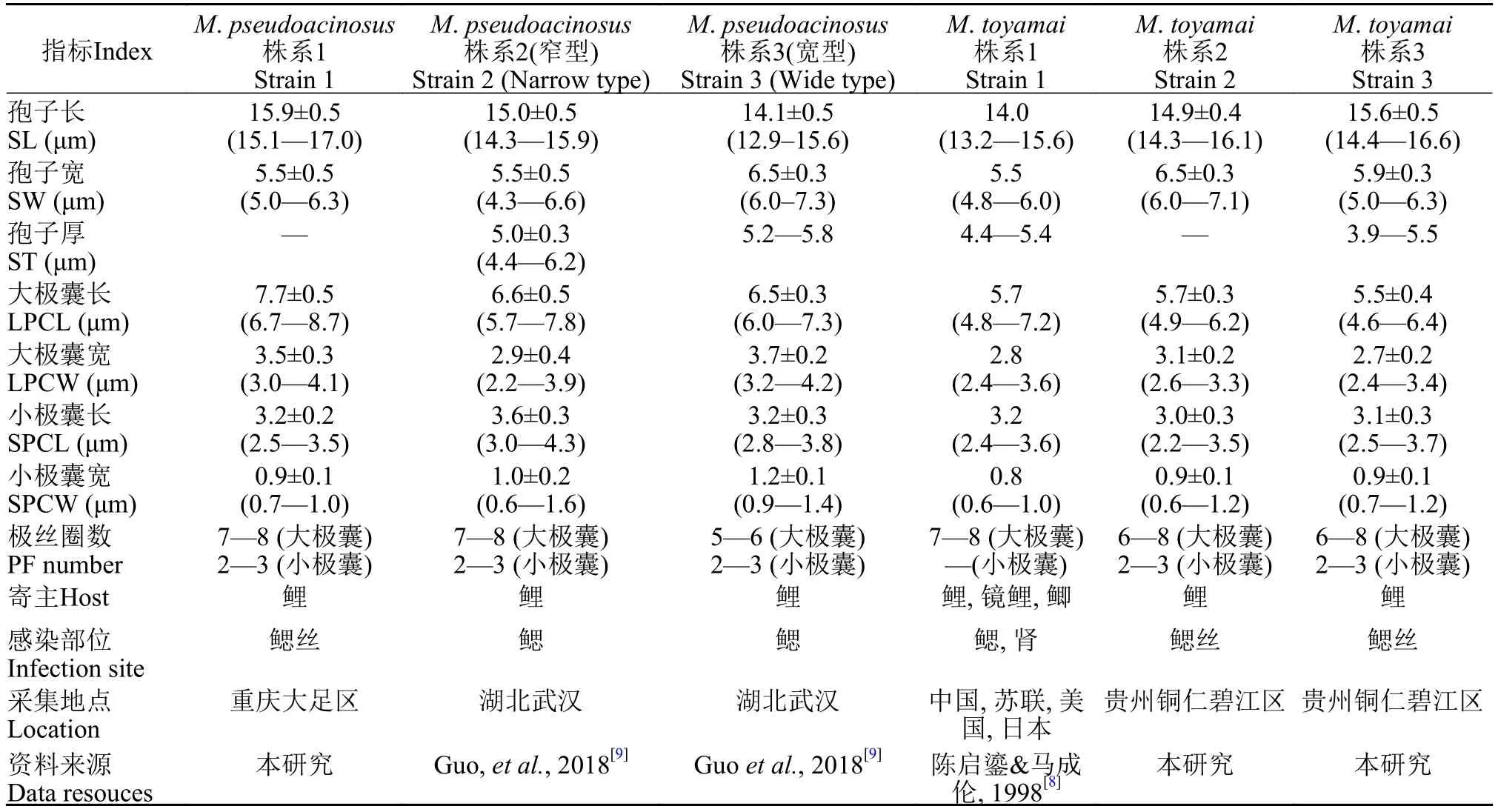
表2 似葡萄碘泡虫和茄形碘泡虫株系形态比较Tab. 2 The morphological comparison among different strains of Myxobolus pseudoacinosus and Myxobolus toyamai

图2 葡萄碘泡虫、似葡萄碘泡虫和茄形碘泡虫成熟孢子形态图Fig. 2 Mature spores of M. acinosus, M. pseudoacinosus and M. toyamai
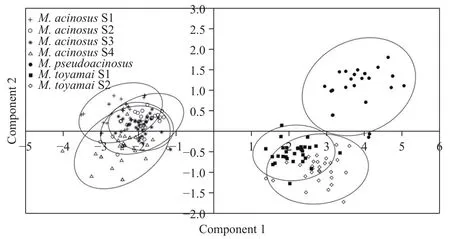
图3 基于葡萄碘泡虫、似葡萄碘泡虫和茄形碘泡虫的形态学数据的主成分分析图Fig. 3 Principal component analysis based on morphological data of M. acinosus, M. pseudoacinosus and M. toyamai
2.2 葡萄碘泡虫、似葡萄碘泡虫和茄形碘泡虫18S rRNA基因分析
获得葡萄碘泡虫S1—S4株系18S rDNA序列长度分别为1986 nt(GenBank登录号: MW821466)、1899 nt(GenBank登录号: MW821467)、1896 nt(GenBank登录号: MW821468)和1903 nt(Gen-Bank登录号: MW821469)。4株系间序列相似度为100%, 遗传距离为0.000—0.001, 变异位点有0—1个。4株系18S rDNA序列与GenBank数据库中的葡萄碘泡虫(KX810022、KX810021)的相似度最高(99.9%—100%)。6条序列之间相似度为99.9%—100%,遗传距离为0.000—0.002, 变异位点有0—2个(图 4)。
获得似葡萄碘泡虫18S rDNA序列1条, 序列长度1897 nt(GenBank登录号: MW821470), 与Gen-Bank数据库中的似葡萄碘泡虫(KX586684和KX810019)相似度最高(99.8%), 三条序列的相似度为99.6%—99.8%, 遗传距离为0.002—0.007, 变异位点有4—8个(图 4)。
获得茄形碘泡虫S1和S2株系18S rDNA序列长度分别为1641 nt(GenBank登录号: MW821471)和1758 nt(GenBank登录号: MW821472), 两株系序列相似度为100%, 遗传距离为0.000, 变异位点0个。两株系与GenBank数据库中的茄形碘泡虫(LC010116、C010115、FJ710802和HQ338729)相似度最高(99.5%—100%), 6条序列之间相似度为99.5%—100%,遗传距离为0.000—0.003, 变异位点有0—3个(图 4)。
葡萄碘泡虫与似葡萄碘泡虫之间的18S rDNA相似度为98.4—98.8%, 遗传距离为0.013—0.020,变异位点有23—26个; 葡萄碘泡虫与茄形碘泡虫18S rDNA序列相似度为96.1—97.2%, 遗传距离为0.038—0.042, 变异位点有55—58个; 似葡萄碘泡虫和茄形碘泡虫18S rDNA序列相似度为96.4—97.6%,遗传距离为0.033—0.040, 变异位点有46—63个(图 4)。
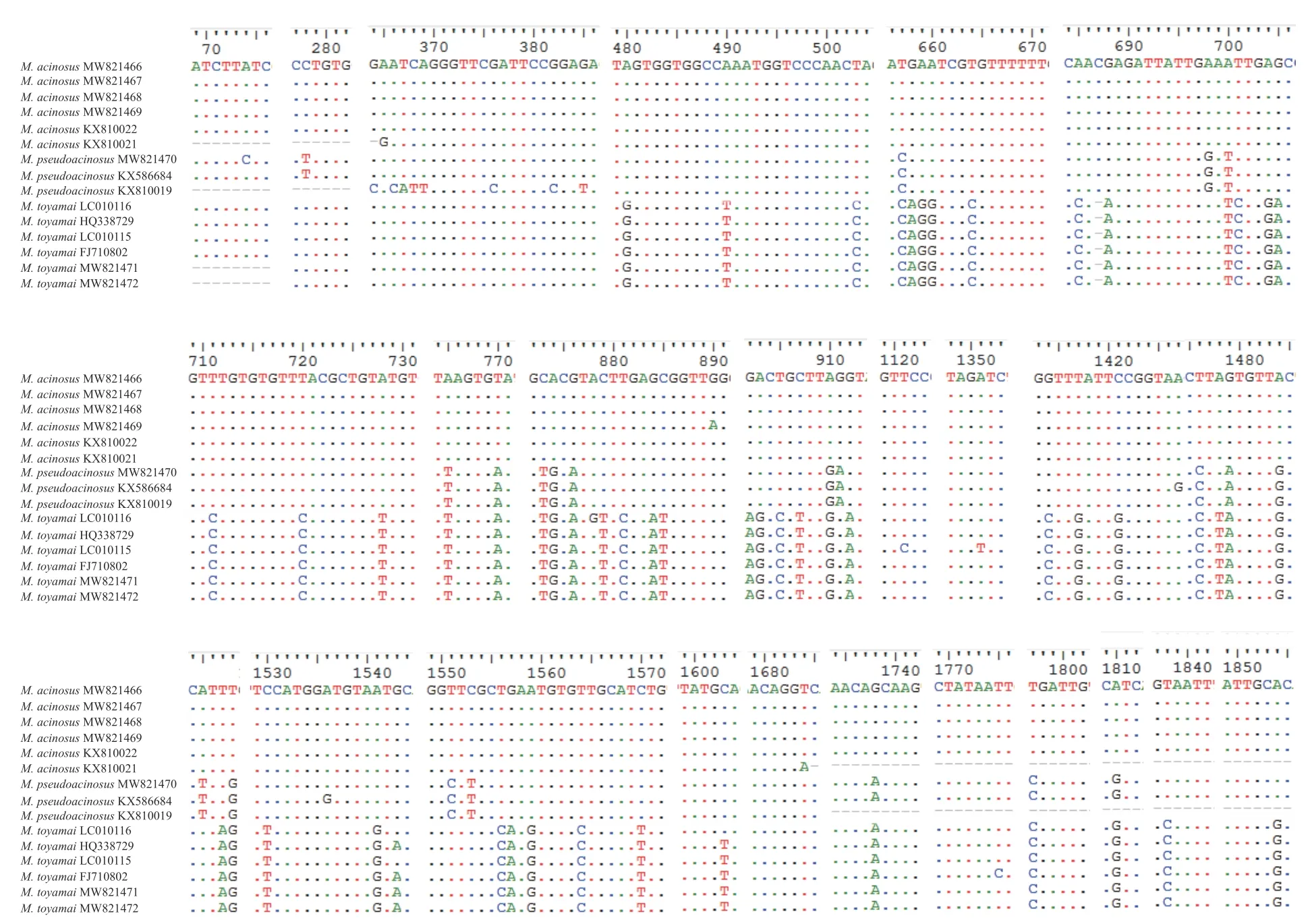
图4 葡萄碘泡虫、似葡萄碘泡虫和茄形碘泡虫18S rDNA的变异位点Fig. 4 The variable sites of 18S rDNA among M. acinosus, M. pseudoacinosus and M. toyamai
本文对葡萄碘泡虫、似葡萄碘泡虫和茄形碘泡虫的18S rRNA二级结构中的V1、V2、V3、V4、V5、V6、V7、V8 和V9等9个可变区进行了分析, 结果显示V4区的E23-2构型可将葡萄碘泡虫与似葡萄碘泡虫和茄形碘泡虫有效地区分开: 与似葡萄碘泡虫和茄形碘泡虫相比较, 葡萄碘泡虫E23-2的发卡环更大(8碱基vs.11碱基), 且在发卡环旁边少一个单碱基(A碱基)侧凸(图 5)。同时, V7区的H43构型可以将茄形碘泡虫分别与葡萄碘泡虫和似葡萄碘泡虫有效区分: 与葡萄碘泡虫和似葡萄碘泡虫相比较, 茄形碘泡虫H43的内部环更大(9碱基vs.10碱基), 而发夹环更小(7碱基vs.6碱基)。
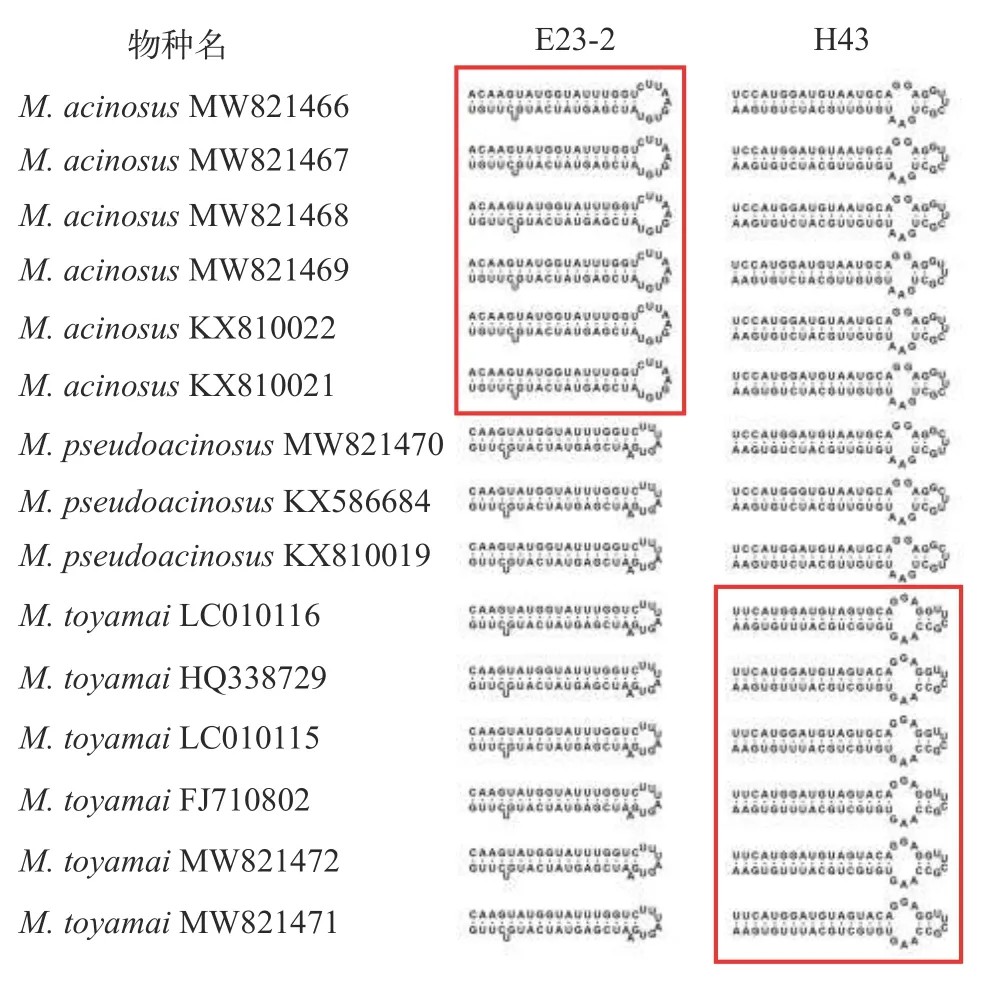
图5 葡萄碘泡虫、似葡萄碘泡虫和茄形碘泡虫的18S rRNA二级结构的E23-2 和 H43构型Fig. 5 The model of E23-2 and H43 of secondary structure of 18S rRNA among M. acinosus, M. pseudoacinosus and M. toyamai
2.3 系统发育树
基于18S rDNA构建的BI和ML树呈现相似的拓扑结构(图 6)。系统发育树首先分为两大进化支(Clade A和Clade B, 图 6), 葡萄碘泡虫、似葡萄碘泡虫和茄形碘泡虫为A进化支中分化较晚的一支。虽然葡萄碘泡虫和茄形碘泡虫有些种内株系间的系统关系在ML树和BI树中不一致, 但两系统树均支持葡萄碘泡虫各株系聚为一支, 似葡萄碘泡虫各株系聚为一支, 茄形碘泡虫各株系聚为一支。在亚支Ⅰ中(Subclade I, 图 6), 似葡萄碘泡虫位于基部位置, 葡萄碘泡虫与长孢碘泡虫形成姐妹群关系, 茄形碘泡虫与拟茄形碘泡虫(GenBank登录号: LC228237)形成姐妹群关系。

图6 基于18S rDNA序列构建的ML/BI树Fig. 6 Phylogenetic trees generated by ML/BI methods based on 18S rDNA sequences
3 讨论
葡萄碘泡虫、似葡萄碘泡虫和茄形碘泡虫有着共同的宿主(鲤)和寄生部位(鳃), 成熟孢子均前端较尖后端圆钝, 均具一大一小极囊, 在鉴定中容易混淆, 但三者形态仍有细微差别: 葡萄碘泡虫孢子为长葡萄形, 可与孢子呈茄形的似葡萄碘泡虫和茄形碘泡虫区分; 与茄形碘泡虫相比, 似葡萄碘泡虫孢子更细长且其大极囊在孢子腔内的占比更大(图 2)。另外, 主成分分析显示, 三者形态量度存在显著差异(图 3)。虽然学者们普遍认为物种鉴定的遗传学阈值并不存在, 但基于18S rDNA, 有学者认为大多数黏孢子虫的种内相似度范围应为99.0%—100%[26], 冉佼等[27]认为种内遗传距离范围大多数集中在0.000—0.007, Zhao等[28]认为种内变异位点应少于10个。葡萄碘泡虫与似葡萄碘泡虫的18S rDNA相似度为98.4—98.8%, 遗传距离为0.013—0.020, 变异位点有23—26个, 超出上述种内范围, 两者应为种间水平。类似的, 葡萄碘泡虫与茄形碘泡虫及似葡萄碘泡虫与茄形碘泡虫也应属于种间水平。另外, 18S rRNA二级结构V4区的E23-2构型可将葡萄碘泡虫分别与似葡萄碘泡虫和茄形碘泡虫区分, 而V7区的H43构型可将该虫与葡萄碘泡虫和似葡萄碘泡虫区分(图 5)。同时, 通过18S rDNA序列比对发现, 葡萄碘泡虫含有15个关键变异位点可将该虫与似葡萄碘泡虫和茄形碘泡虫区分; 似葡萄碘泡虫含有5个关键变异位点可将该虫与葡萄碘泡虫和茄形碘泡虫区分; 茄形碘泡虫含有33个关键变异位点可将该虫与葡萄碘泡虫和似葡萄碘泡虫区分(图 4)。综上表明三者无论在形态上还是遗传上均具有独立物种的特征。
系统发育分析显示, 黏孢子虫成熟孢子的形态与系统发育关系密切, 如A进化支(Clade A)中大部分物种孢子形态为梨形, 前端稍尖, 后端圆钝; 进化支B(Clade B)中大多数物种孢子形态为圆形或椭圆形, 前端和后端都较圆钝(图 6)。但仍有一些黏孢子虫的形态并不完全符合该规律, 如A支中圣安德烈碘泡虫Myxobolus szentendrensisCech,et al.,2015, 鳃侧碘泡虫Myxobolus branchilateralisMolnár,et al., 2012, 肥满碘泡虫Myxobolus obesusGurley, 1893, 埃拉斯碘泡虫Myxobolus eirasianusCech,et al., 2012, 射阳碘泡虫Myxobolus sheyangensisLiu,et al., 2016, 塔形碘泡虫Myxobolus pyramidisChen, 1958, 坠形碘泡虫Myxobolus pendulaGuilford, 1967, 卡拉瓦提碘泡虫Myxobolus kalavatiaeSzékely,et al., 2015 和内膜碘泡虫Myxobolus intimusZaika, 1965的孢子形态更接近B支中圆形或椭圆形孢子形态(图 6)。在B支中, 中华单极虫Thelohanellus sinensisChen & Hsieh, 1960和龟鳖单极虫Thelohanellus testudineusLiu,et al, 2014的孢子形态更接近A支中的梨形孢子形态(图 6)。
以往的研究和本文都表明, 黏孢子虫的系统发育与宿主类群关系密切: 相同分类阶元(如目、科)的宿主寄生的黏孢子虫之间有着较近的系统发育关系[29—32], 如本文系统发育树中的黏孢子虫均寄生于鲤形目鲤科鱼类。在寄生虫与宿主的关系中,宿主转移是人们普遍关注的问题之一, 一些寄生虫通过探索新的宿主来促进自身对环境的适应。宿主转移可能导致寄生虫宿主范围的扩大或者放弃原来的宿主, 这是在系统发育树中处在同一支系且亲缘关系较近的寄生虫对应的宿主亲缘关系却较远的首要原因[33]。如本文系统发育树中的穆勒碘泡虫Myxobolus muelleriBütschli, 1882除了感染鲤形目鲤科鱼类, 还能感染鲈形目杜父鱼科鱼类和鲻形目鲻科鱼类。另外, 中华单极虫除了感染鲤形目鲤科鱼类, 还能感染鲈形目鳢科鱼类、鲈形目虾虎科鱼类及鳗形目鳗科鱼类。通过黏孢子虫的系统发育与宿主类群关系的分析推测, 穆勒碘泡虫和中华单极虫的原始宿主应为鲤科鱼类, 其他类群应是这两种黏孢子虫通过宿主转移方式获得的新宿主类群, 从而扩大了寄生虫的宿主范围。
