忍冬属黄酮合成酶基因FNSⅡ密码子偏好性及进化分析
2022-05-27何志敏曾子豪朱肖霞余清婷胡珊巍满威刘素纯
何志敏,曾子豪,朱肖霞,余清婷,胡珊巍,满威,刘素纯
1.长沙学院生物与环境工程学院,湖南 长沙 410003;2.湖南农业大学食品科技学院,湖南 长沙 410128
《中华人民共和国药典》自2005年版起增加了金银花中黄酮类化合物木犀草苷含量的测定,但对归属山银花的4个种属(灰毡毛忍冬、红腺忍冬、华南忍冬、黄褐毛忍冬)中木犀草苷的含量未做出明确规定,所以木犀草苷成为区分金银花与山银花的一个重要化学指标,也是金银花与山银花一物一名分列的重要依据。
黄酮合成酶基因(FNSⅠ和FNSⅡ)是植物黄烷酮转化为黄酮的关键酶基因,植物内木犀草苷的含量受黄酮合成酶基因表达的调控,基因表达的高低与其在长期进化过程中形成的密码子偏好性有关。研究表明,忍冬黄酮合成酶基因FNSⅡ编码的木犀草苷酶在金银花和山银花中的酶催化活性大小是影响木犀草苷生物合成的关键因素。
目前已有许多植物的FNSⅡ基因的cDNA被克隆和表征,包括大豆、甘草、苜蓿、拟南芥、非洲菊、高粱等。在长期进化过程中,由于环境压力及突变选择,不同物种会倾向性选择不同的密码子模式,形成密码子偏好性。提高FNSⅡ基因表达效率对其功能研究具有重要意义,而其密码子偏好性与其表达效率有必然联系。通过密码子偏好性分析,不仅可以了解不同物种之间的遗传进化关系,还能对基因的外源表达选择优先宿主,通过优化密码子以提高基因的外源表达效率。
本研究对23个不同物种的FNSⅡ基因及本实验室克隆获得的29条忍冬属FNSⅡ基因序列进行密码子偏好性及进化分析,并进行忍冬FNSⅡ的外源表达系统和遗传转化受体的选择分析,为进一步开展忍冬FNSⅡ基因的功能研究及外源高表达提供依据。
1 资料
通过美国国家生物技术信息中心(NCBI)数据库(http://www.ncbi.nlm.nih.gov)下载包括金银花、灰毡毛忍冬、烟草等23个具有完整编码区(CDS)植物FNSⅡ基因序列,另外29条序列为本实验室从已通过鉴定的忍冬属植物金银花、灰毡毛忍冬、黄褐毛忍冬、净花菰腺忍冬、红腺忍冬中克隆获得,来源信息见表1。

表1 23个物种FNSⅡ基因序列及29条忍冬属FNSⅡ克隆序列来源信息
2 方法与结果
2.1 密码子偏性分析
将NCBI下载的23个物种FNSⅡ基因序列去除5’UTR和3’UTR序列,选择CDS序列,在txt文档中建立fasta格式,导入Codon W软件,对52条FNSⅡ基因序列进行分析,获得密码子偏好性相关参数,包括相对同义密码子使用度(RSCU)、密码子第三位各碱基含量、有效密码子数(ENC)、密码子适应指数(CAI)、密码子偏爱指数(CBI)、最优密码子使用频率(FOP)等。
2.1.1 FNSⅡ相对同义密码子使用度
Codon W分析获得52条FNSⅡ序列的RSCU值并将RSCU值标准化后,获得FNSⅡ的RSCU值热图。
不同物种FNSⅡ序列的RSCU有一定差异。23个物种的FNSⅡ序列中,最优密码子为CUC,其均值为1.63。丹参、粘毛黄芩和贝加尔黄芩RSCU>2的密码子最多,均有7个,见图1。粘毛黄芩和贝加尔黄芩的密码子RSCU十分相近,两者与丹参在多个密码子上的RSCU值均大于1,在密码子偏好性上有一定相似性,表明其在进化过程中亲缘关系较近。
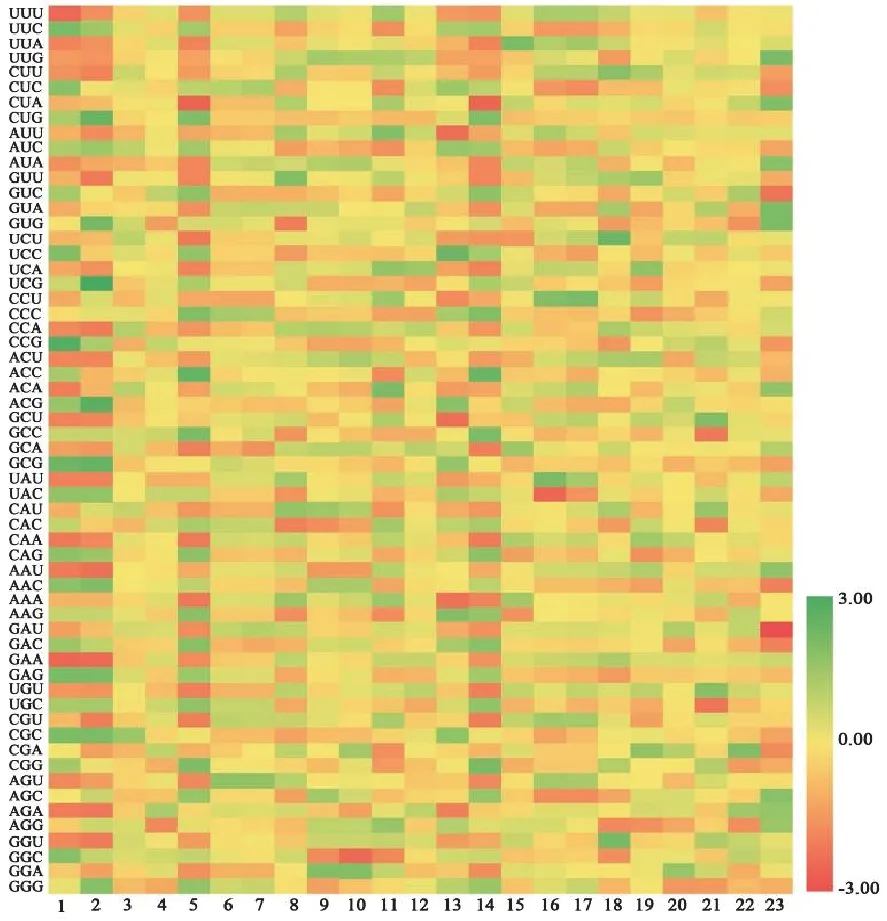
图1 23个物种FNSⅡ序列的RSCU热图
31条忍冬属FNSⅡ序列RSCU热图见图2。忍冬属的最优密码子也为CUC,其均值为2.70,表明大多数密码子在FNSⅡ中的相对使用频率较高。净花菰腺忍冬(2-5)和红腺忍冬(2-8)的最优密码子CUC的RSCU值达3.29,表明两者的FNSⅡ序列在表达时对CUC的偏好性较强。
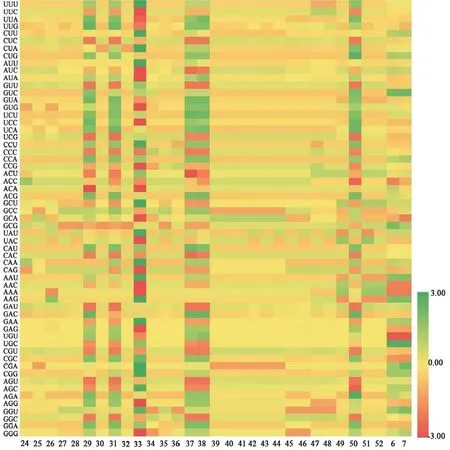
图2 31条忍冬属FNSⅡ序列的RSCU热图
2.1.2 FNSⅡ特性分析
同义密码子的差异通常体现在密码子第3位。在23个不同物种的FNSⅡ序列中,单碱基U3s、C3s、A3s、G3s平均含量分别为0.306、0.350、0.295、0.312,GC3s和GC平均含量分别为0.514和0.469。但白皮松、丹参、粘毛黄芩、蝴蝶花×荷兰鸢尾、贝加尔黄芩GC3s含量均高于70%,GC含量达56%以上,而其他物种的GC含量与GC3s含量相当,大部分在45%~50%,少部分如灯盏花、苜蓿、淫羊藿等的GC和GC3s含量低于40%。忍冬属FNSⅡ序列GC3s和GC平均含量分别为0.521和0.459,与其他物种的平均含量相当,结果见表2。表明大部分植物的FNSⅡ序列第3位碱基对G/C和A/T结尾的偏好程度相似。
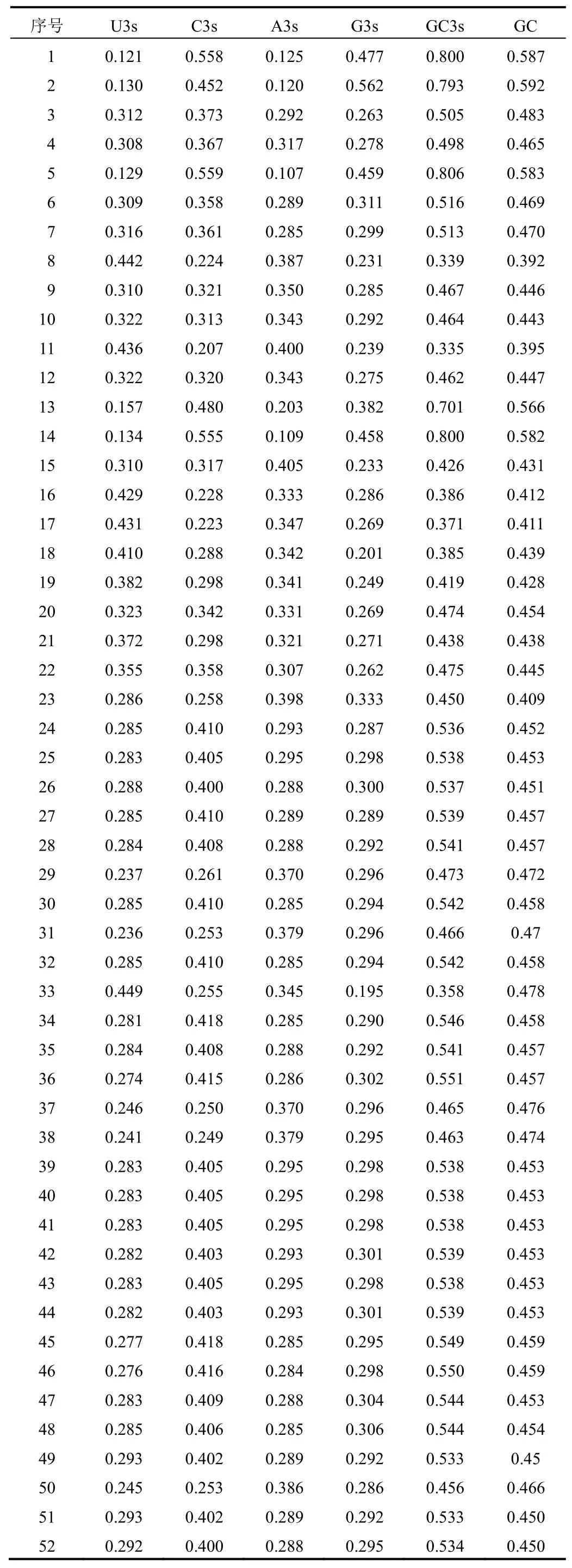
表2 52条FNSⅡ基因密码子碱基组成特性分析
为系统评估FNSⅡ基因的密码子使用偏好性程度,对不同物种的FNSⅡ基因密码子偏好性参数进行分析。从整体来看,各物种的ENC介于39~58,平均值为52.29,忍冬属FNSⅡ的ENC介于50~57,平均值为52.66。FOP值介于0.326~0.528,平均值为0.392,表明大部分FNSⅡ基因的密码子偏好性较弱。同时,不同植物FNSⅡ的CAI较小(0.117~0.257),且大部分植物FNSⅡ的ENC值较大。52条FNSⅡ基因的密码子偏好性参数见表3。分析结果表明,FNSⅡ基因密码子偏好性弱,且在不同植物中的表达水平可能较低。此外,不同植物FNSⅡ的ENC值存在一定差异,表明FNSⅡ基因在不同植物中的表达水平不同。
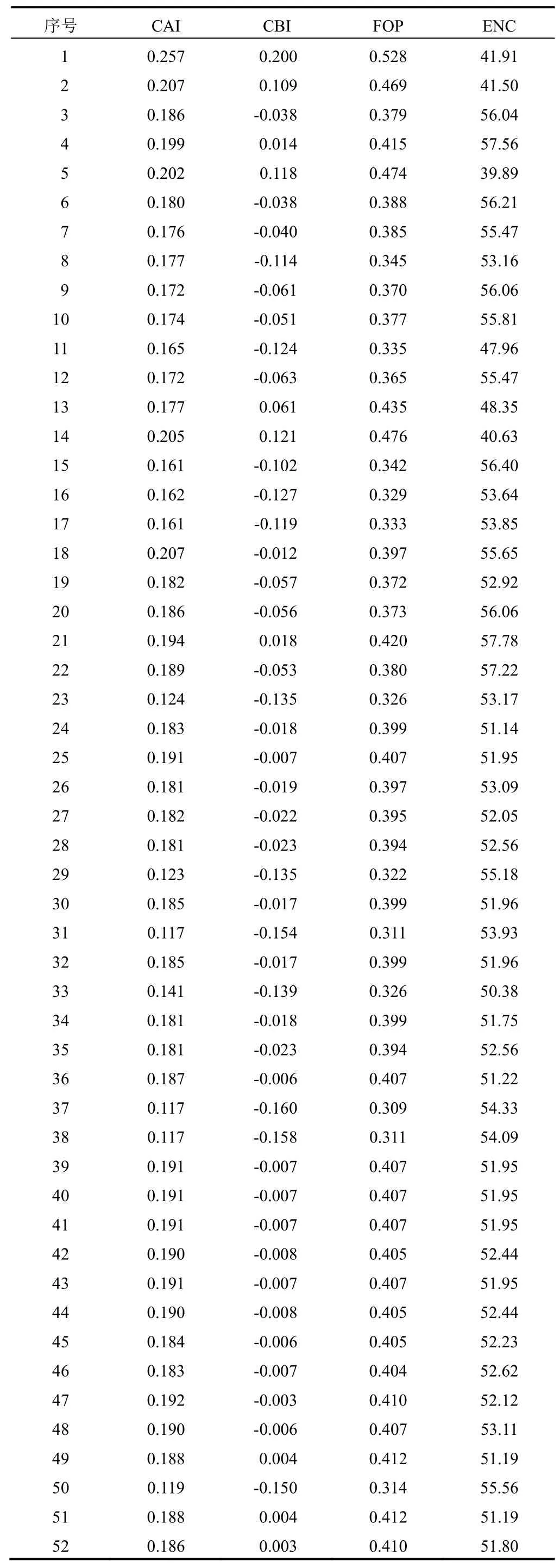
表3 52条FNSⅡ基因密码子偏好性参数
2.2 ENC-Plot分析
以GC3s为自变量,以ENC为因变量,按ENC=2+GC3s+29/[GC3s+(1-GC3s)]建立期望曲线。将所得各序列以GC3s为横坐标、ENC为纵坐标绘制散点图,不同物种的FNSⅡ基因ENC-Plot图见图3。可以看出,在期望曲线附近分布着大部分植物FNSⅡ基因,距离期望曲线最近的是灯盏花、淫羊藿、三花龙胆,该结果表明,突变压力是这些植物FNSⅡ基因密码子偏好性形成的主要原因。而自然选择主导烟草、忍冬、翠菊、苜蓿的FNSⅡ基因密码子偏好性的形成,其分布于离期望曲线较远的位置。

图3 不同物种FNSⅡ基因ENC-Plot分析
忍冬属FNSⅡ基因普遍分布于离期望曲线稍远的位置,尤其是灰毡毛忍冬、华南忍冬、黄褐毛忍冬,结果见图4,表明忍冬属植物FNSⅡ基因密码子偏好性形成主要受自然选择的影响。

图4 忍冬属FNSⅡ基因ENC-Plot分析
2.3 FNSⅡ系统进化分析
将列入所有序列的fasta文件导入Clustal X软件,进行序列比对后,选择bootstrap N-J tree聚类分析(bootstrap=1 000),输出.phd的结果文件,将该文件导入MEGA4.0软件,构建系统发育树,结果见图5。通过邻接法构建的基于52条FNSⅡ CDS序列系统进化树主要分为忍冬属和非忍冬属两大分支,符合一般植物分类的规律。当中可以发现忍冬属植物中大部分聚为一个大类,各FNSⅡ按植物亲缘关系远近聚为一类,但除黄褐毛忍冬和华南忍冬外,其他忍冬属植物之间有混合交错分布而聚为一类的现象,而华南忍冬甚至与非忍冬属植物聚为一类。同比之下,非忍冬属植物较符合分类规律,同种属的2种大豆,粘毛黄芩和贝加尔黄芩,同为菊科的野菊花、灯盏花、大蓟等分别聚为一类,亲缘关系较远的三花龙胆、山茶树、苜蓿则独自分为一类。
2.4 聚类分析
利用SPSS18.0软件,以52条FNSⅡ序列RSCU为变量,采用组间连接法,进行密码子偏好性聚类分析。基于FNSⅡ基因RSCU的聚类分析结果(见图5)与基于CDS序列的系统进化分析结果(见图6)基本一致。在RSCU聚类中,绝大多数忍冬属植物聚为一个大类,但在另一小分支,灰毡毛忍冬、净花菰腺忍冬、黄褐毛忍冬、红腺忍冬及烟草聚为一类,灯盏花、非洲菊等菊科植物聚成一类,但夹杂了小豆蔻和3个忍冬属样本;黄芩属植物、丹参、蝴蝶花×荷兰鸢尾、白皮松聚为一类;大豆、山茶树、淫羊藿、三花龙胆聚为一类。表明其存在核酸序列一致性较高的FNSⅡ基因。同为忍冬属植物的FNSⅡ基因分布于多个类别,其中14个灰毡毛忍冬的FNSⅡ基因分布于多个大类,与其他植物聚成一小支,与系统进化树的归类情况相似但存在一定差异。同种植物的FNSⅡ基因被聚类到不同分支上,说明其FNSⅡ基因具有密码子偏好性差异。以上结果表明,FNSⅡ基因的密码子偏好性差异大小与物种亲缘关系远近有一定关系,反映了物种的进化关系,但基于RSCU的聚类在反映物种进化关系准确度方面逊于基于CDS序列的系统进化树。
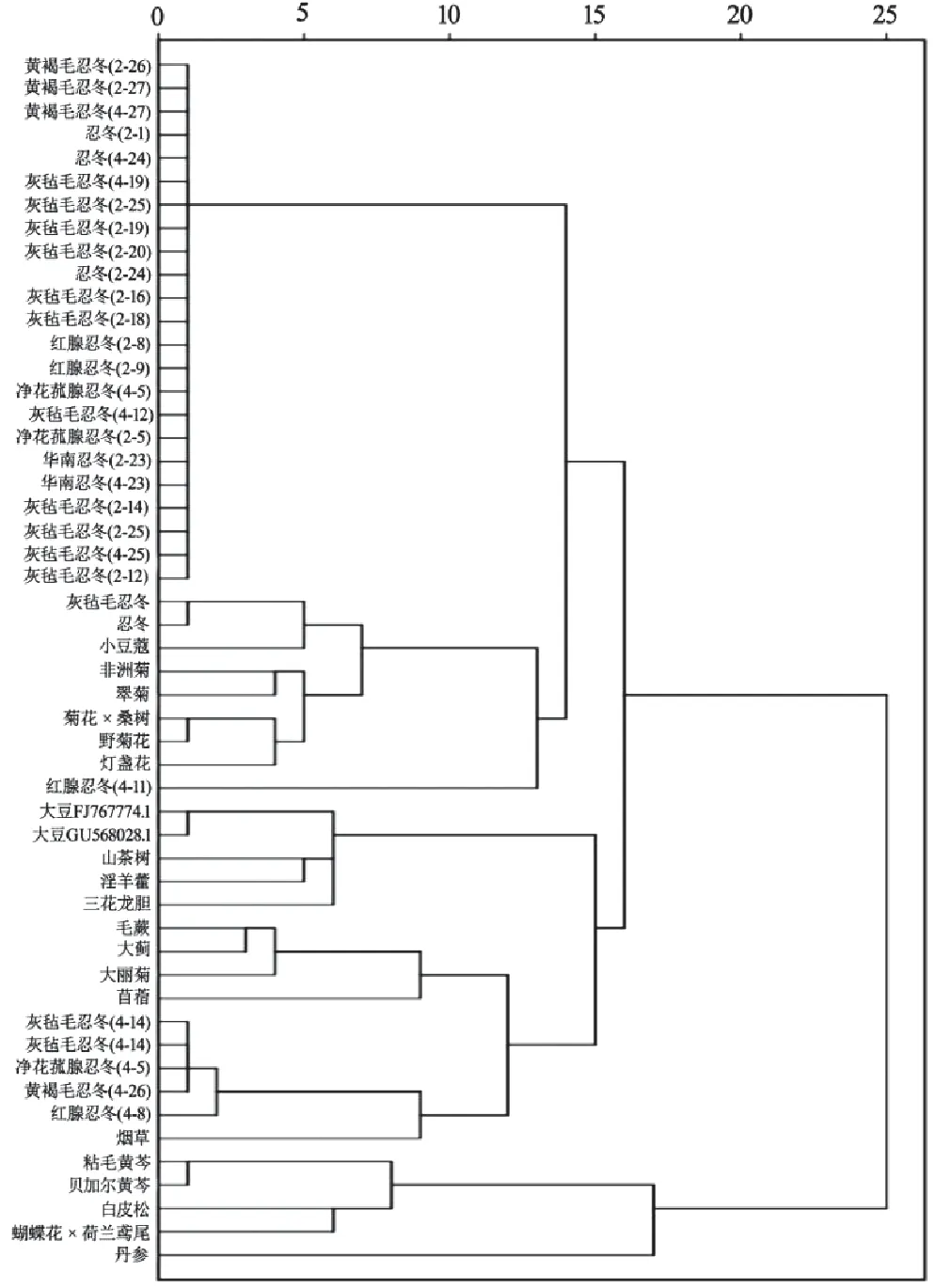
图5 FNSⅡ的RSCU聚类分析树状图

图6 FNSⅡ系统进化分析
2.5 忍冬FNSⅡ与模式生物密码子偏好性比较
通过比较忍冬()与模式生物拟南芥()、烟草L.()、大肠杆菌()和酵母()FNSⅡ基因的密码子使用频率,分析频率差异性较大(频率比值<0.5或>2)的密码子个数,以确定最佳遗传转化受体和外源表达系统,结果见表4。大肠杆菌、酵母与忍冬FNSⅡ基因差异较大的密码子个数分别为34、31,说明忍冬FNSⅡ的最佳外源表达是酵母,但实际应用时需要对外源表达的FNSⅡ密码子进行优化以提高表达效率。拟南芥、烟草与忍冬FNSⅡ的差异性密码子数分别为26、27,说明两者均可作为忍冬FNSⅡ的遗传转化受体。
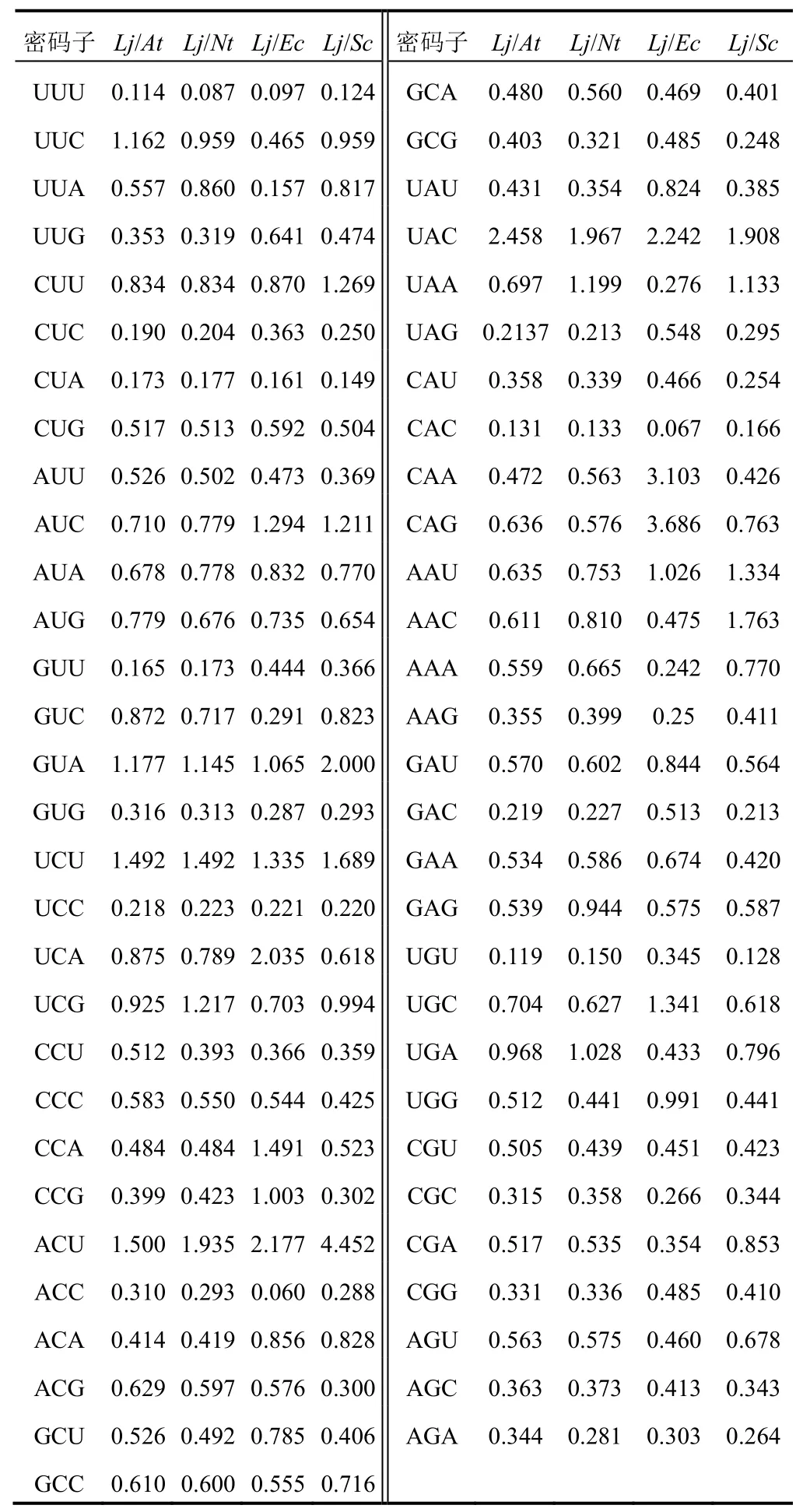
表4 忍冬FNSⅡ与模式生物基因组密码子使用频率比较
3 讨论
本研究首先对23条不同种植物及29条本实验室克隆的忍冬属FNSⅡ基因进行密码子偏好性整体分析,发现不同种植物FNSⅡ及忍冬属FNSⅡ的最优密码子为CUC。通过分析不同物种RSCU>2的偏好性密码子发现,丹参、粘毛黄芩和贝加尔黄芩中均有7个,而忍冬属植物除红腺忍冬(4-11)有5个外,其余均为2个,表明忍冬属植物FNSⅡ基因对优势密码子的使用偏好性较弱。金银花和山银花FNSⅡ的密码子RSCU都具有很高的相似性。进一步分析ENC和FOP,发现23个不同种植物FNSⅡ基因的ENC均值为52.29,忍冬属植物FNSⅡ基因的ENC均值为52.66,52个样本的FOP均值接近0.36,表明所选52个样本FNSⅡ基因的密码子偏好性普遍较弱。上述ENC和RSCU分析结果均表明植物FNSⅡ密码子偏好性较弱。CAI与基因表达水平通常呈正相关,而ENC则相反。本研究52条FNSⅡ序列的CAI均小于0.3,而ENC均大于50,表明FNSⅡ基因表达水平较低,而低水平表达的基因往往对密码子的偏好性也较低,与前述分析结论一致。ENC-plot分析结果表明,突变压力是大部分植物FNSⅡ基因偏好性形成的主因,仅有少数如烟草、忍冬、翠菊、苜蓿的FNSⅡ基因主要受自然选择影响或其他因素的影响,如基因长度、结构等。
亲缘关系较近的物种在密码子使用偏好性方面趋向于相同或相近。本实验室克隆的忍冬属FNSⅡ基因在各种属间的同源性高达95%以上,在密码子使用偏好性方面具有很高的相似性。基于FNSⅡ序列的进化分析与传统的植物分类结果基本一致,表明FNSⅡ基因与植物的进化过程密切相关。同为忍冬属的不同种有混合交错聚为一支的现象,如忍冬与灰毡毛忍冬、黄褐毛忍冬都有聚为一支的现象。其原因除与FNSⅡ序列的高同源性有关外,可能还与该基因影响的木犀草苷在忍冬、灰毡毛忍冬及黄褐毛忍冬中的含量有关,目前在大部分山银花中未检测到木犀草苷,但通过超高效液相色谱和高效液相色谱-串联质谱能在灰毡毛忍冬和黄褐毛忍冬中检测到少量木犀草苷。以主成分含量为变量对5个忍冬属种质资源进行的系统聚类分析结果显示,灰毡毛忍冬和黄褐毛忍冬与忍冬种质差异较其他山银花种属小。另外,华南忍冬与非忍冬属植物归为一个大类,存在与传统的植物分类规律冲突的现象。已有研究表明,华南忍冬野生居群具有较高水平的遗传多样性,居群间遗传分化程度较高,地理距离与遗传距离之间具有显著相关性,地理隔离效应是导致居群间遗传分化的重要因素。所以,推测其可能受到生长的地理环境影响,导致华南忍冬的FNSⅡ基因在分化过程中发生了一定程度的变异。
在进化过程中,相同物种或相近种属会形成相似的密码子偏好性,密码子偏好性能在一定程度上反映不同物种间亲缘关系的远近。基于RSCU的聚类分析结果显示,大部分忍冬属植物的FNSⅡ基因聚为一支,表明亲缘关系较近的物种有相似的密码子偏好性。少部分忍冬属植物被聚为其他大类的分支。14个灰毡毛忍冬与4个红腺忍冬的FNSⅡ基因距离较远,分别与其他多种植物聚为一支,说明其FNSⅡ基因的密码子偏好性差异较大。同种植物的FNSⅡ基因被聚类到不同分支上,说明在进化过程中由于突变或自然选择等压力导致地域差异,同种属FNSⅡ基因之间的密码子偏好性不同。RSCU聚类在一定程度上能够反映出物种进化的关系,但容易受到更多因素的影响,比较适合在较小的分类单元里提供较为可靠的分类依据,因此,与RSCU聚类相比,基于CDS序列的系统进化树分析的准确度更高。
通过对比分析忍冬与模式生物的密码子使用频率,发现酵母为忍冬的最佳外源表达系统,但为达到较高水平的蛋白表达需要对密码子进行优化,而拟南芥和烟草均可作为其遗传转化受体。本研究可为FNSⅡ基因的分类与演化表达调控及植物遗传育种等研究提供科学依据,对优化金银花和山银花鉴别方法的研究提供指导。
