亏祖山秋季不同海拔土壤纤毛虫优势种生态位及其与环境因子的关系
2022-05-26王壮壮朱时应张秋涵何梦萨
王壮壮,朱时应,张秋涵,何梦萨,普 布
西藏大学理学院生命科学系高原动物学实验室,拉萨 850000
土壤原生动物具有丰富的物种多样性及巨大的生物量,在土壤生态系统的物质循环和能量流动中扮演着重要角色[1—2]。土壤纤毛虫是土壤原生动物三大类群之一,是单细胞真核生物中特化强度高,结构功能最复杂的类群,具有个体小、种类和数量繁多、繁殖速度较快、比表面积大、对环境反映灵敏,环境适应性强等特点[3—5]。土壤纤毛虫直接或间接参与土壤有机物的分解和转化,在土壤生态系统的能量流动与物质循环中发挥着十分重要作用。土壤纤毛虫被作为一种良好的环境指示生物,其环境效应参数(多样性特征和群落结构的变化等)可作为评价、监测以及预报土壤环境变化的指标[6—8]。目前国内草甸土壤纤毛虫的研究区域主要集中在沼泽化草甸、甘南亚高寒草甸和青藏高原东缘高寒草甸等地,研究内容以土壤纤毛虫的群落结构特征和多样性为主[9—11],缺乏对土壤纤毛虫资源利用情况和种间关系的系统研究。西藏高寒草甸土壤纤毛虫相关研究报道较少,对土壤纤毛虫生态位的研究尚未见报道。
西藏位于青藏高原西南部,海拔高、气候多样、生态系统脆弱,环境遭到破坏后较难恢复。由于其独特的地理环境,研究土壤动物类群多样性、种间关系及其与环境的关系尤为重要。本文以秋季拉萨河流域林周县白朗村亏祖山高寒草甸土壤纤毛虫为研究对象,旨在深入了解亏祖山高寒草甸生态系统中土壤纤毛虫优势种生态位宽度、生态位重叠值、可利用资源的变化以及与环境因子之间的关系,探讨土壤纤毛虫优势种空间资源利用情况和种间关系,为拉萨河流域亏祖山高寒草甸的土壤生物多样性保护和生态环境改善、退化生态系统的恢复提供基础数据,为亏祖山建立高寒草甸退化的预警指示系统和高寒草甸生态系统的可持续发展提供科学依据。
1 材料与方法
1.1 研究区域基本概况
西藏拉萨市林周县卡孜乡白朗村亏祖山位于东经91°07′33″—91°07′19″E,北纬29°52′23″—29°49′50″N,属于拉萨河支流澎波河谷西南部,距林周县15 km左右,山体海拔跨度在3900—5100 m之间,年日照时大于3000 h,年均气温在7.6℃,降水主要集中于每年的6—9 月,降水量在440 mm左右[12—14]。山体分布有高寒草甸草场、高寒矮灌丛到高寒草甸及部分裸露岩石,常见的植物主要有高山嵩草(Kobresiapygmaea)、钉柱委陵菜(Potentillasaundersiana)、青藏苔草(Carexmoorcroftii)、雪层杜鹃(Rhododendronnivale)、樱草杜鹃(Rhododendronprimuliflorum)、金露梅(Potentillafruticosa)、肉果草(Lanceatibetica)、青海刺参(Morinakokonorica)、垫状点地梅(Androsacetapete)和瑞香狼毒(Stellerachamaejasme)等;常见动物有荨麻蛱蝶(Aglaisurticae)、西藏飞蝗(Locustamigratoriatibetensis)、高原鼠兔(Ochotonacurzoniae)、高原兔(Lepusoiostolus)、地山雀(Pseudopodoceshumilis)、棕颈雪雀(Pyrgilaudaruficollis)、藏雪鸡(Tetraogallustibetanus)和高原山鹑(Perdixhodgsoniae)等。
1.2 样点设置和取样方法
2015年10月,在林周县白朗村亏祖山高寒草甸海拔4000—5000 m 之间,大约每100 m海拔梯度设置为一个样地,共设置11个不同海拔梯度的样地(图1),土壤理化因子随海拔的增加出现不同层次的差异,其中pH随海拔的增加呈现先增加后减少的趋势,温度随海拔的增加呈现减小的趋势,样点1总氮(TN)和全磷(TP)含量最高(样点位于山脚下、受放牧等人类活动干扰),而样点10总氮(TN)含量最少,样点5全磷(TP)含量最少,有机质含量百分比各样点间差异不明显;亏祖山植物物种丰富度随海拔呈中间高、两边低的单峰变化格局(表1)。

表1 样点的基本情况Table 1 Basic information of the research sites

图1 拉萨河流域亏祖山样点示意图Fig.1 Schematic diagram sample points on Kuizu Mountain in the Lhasa River Basin
1.3 取样方法
每个样地(20 m×20 m)用圆筒式环刀(高30 cm,直径5 cm)土壤采样器取样,采用梅花五点式采集25个0—15 cm小土样,混合后装入透气布袋,在实验室室温下自然风干,土样风干时在牛皮纸上均匀摊开,使用透气草纸覆盖自然风干。
1.4 土壤纤毛虫的培养和鉴定
自然风干的土壤样品采用定量和定性的方法进行培养和鉴定,定性研究采用“非淹没培养法”,每份取风干土样50 g,分别置于直径为15 cm 的培养皿中,将土壤浸出液加入到培养皿中,土壤完全湿润而不淹没,在25℃的恒温下培养土壤纤毛虫,培养后的第2、4、7、11、14、21、30天放置于光学显微镜下进行形态学鉴定,并记录每个物种的个体数,直到未检出新见物种为止[15—19]。定量研究采用直接计数法,其中每个样点做3次重复,并将数据整合。通过活体观察法对土壤纤毛虫进行了形态学鉴定,相关鉴定资料依据[20—26]。
1.5 理化因子的测定
土温利用曲管地温计(温度范围为-10—60℃)测定;土壤pH值采用电位测定法,用TSS-851土壤湿度酸度计测定;其它土壤理化因子委托西藏博源环境检测有限公司进行测定,土壤含氮量用凯氏法进行硝化处理后,在自动离子分析仪上测定;土壤含磷量用氢氧化钠碱熔-钼锑抗比色法测定;有机质含量采用硫酸、重铬酸钾氧化-容量法测定。
1.6 数据处理和分析
用Excel 2010对数据进行整理,并计算McNaughton优势度指数(Y)[27—28],利用R 4.0.5(Spaa)计算Levins生态位宽度指数(Bi)[29]和Pinaka生态位重叠值(Oik)[30],用Canoco 4.5对物种和环境因子进行去趋势对应分析(DCA)和典范对应分析(CCA),为降低各变量之间的异质性,对环境因子进行标准化处理(Z-score),采用Origin 2019b作图。公式如下:
(1)
(2)
(3)
(4)
(5)
(6)
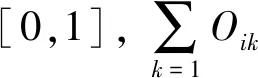
2 结果与分析
2.1 拉萨河流域亏祖山秋季土壤纤毛虫优势种
拉萨河流域林周县白朗村亏祖山高寒草甸11个样点中,共鉴定到土壤纤毛虫132种,隶属于3纲14目48科66属。其中优势种共14个,闪瞬目虫(Glaucomascintillans)和龙骨漫游虫(Litonotuscarinatus)为绝对优势种(Y>0.1),分别占所有优势种个体数的22.76%和11.38%(表2)。闪瞬目虫(Glaucomascintillans)和明显长颈虫(Dileptusconspicuus)在11个样点中均有分布(fi=1)。亏祖山土壤纤毛虫优势种丰度百分比图结果显示,闪瞬目虫(Glaucomascintillans)在样点4、5、6、7、8中所占比例较大,龙骨漫游虫 (Litonotuscarinatus)在样点6和样点11样点中所占比例较大,似肾形虫(Colpodasimulans)和多足尾枝虫(Urostylamullipes)在样点1样点所占比例最大,卵圆前管虫(Prorodonovum)在样点3样点所占比例较大(图2)。对亏祖山土壤纤毛虫优势种丰度聚类结果显示,总体上共分为两类聚集,在距离最近处样点4、5和样点7样点相似性较高聚为一类,而样点9和样点10样点相似性程度高,聚为一类(图2 )。

表2 拉萨河流域亏祖山土壤纤毛虫优势种出现频率(fi)和优势度(Y)Table 2 The occurrence frequency (fi)and dominance degree (Y)of the dominant species of soil ciliates on Kuizu Mountain in the Lhasa River Basin
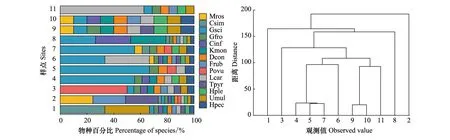
图2 拉萨河流域亏祖山土壤纤毛虫优势种丰度百分比和聚类Fig.2 Abundance percentage and clustering of the dominant species of soil ciliates on Kuizu Mountain in the Lhasa River BasinHpec:点滴半眉虫 Hemiophrys pectinata;Umul:多足尾枝虫 Urostyla mullipes;Hple:肋状半眉虫 Hemiophrys pleurosigma;Tpyr:梨形四膜虫 Tetrahymena pyrifomis;Lcar:龙骨漫游虫 Litonotus carinatus;Povu:卵圆前管虫 Prorodon ovum;Frub:美丽圆纹虫 Furgasonia rubens;Dcon:明显长颈虫 Dileptus conspicuus;Kmon:念珠角毛虫 Keronopsis monilata;Cinf:膨胀肾形虫 Colpoda inflata;Gfro:前口瞬目虫 Glaucoma frontata;Gsci:闪瞬目虫 Glaucoma scintillans;Csim:似肾形虫 Colpoda simulans;Mros:突额扭头虫 Metopus rostratus
2.2 拉萨河流域亏祖山土壤纤毛虫优势种生态位
2.2.1土壤纤毛虫优势种生态位宽度
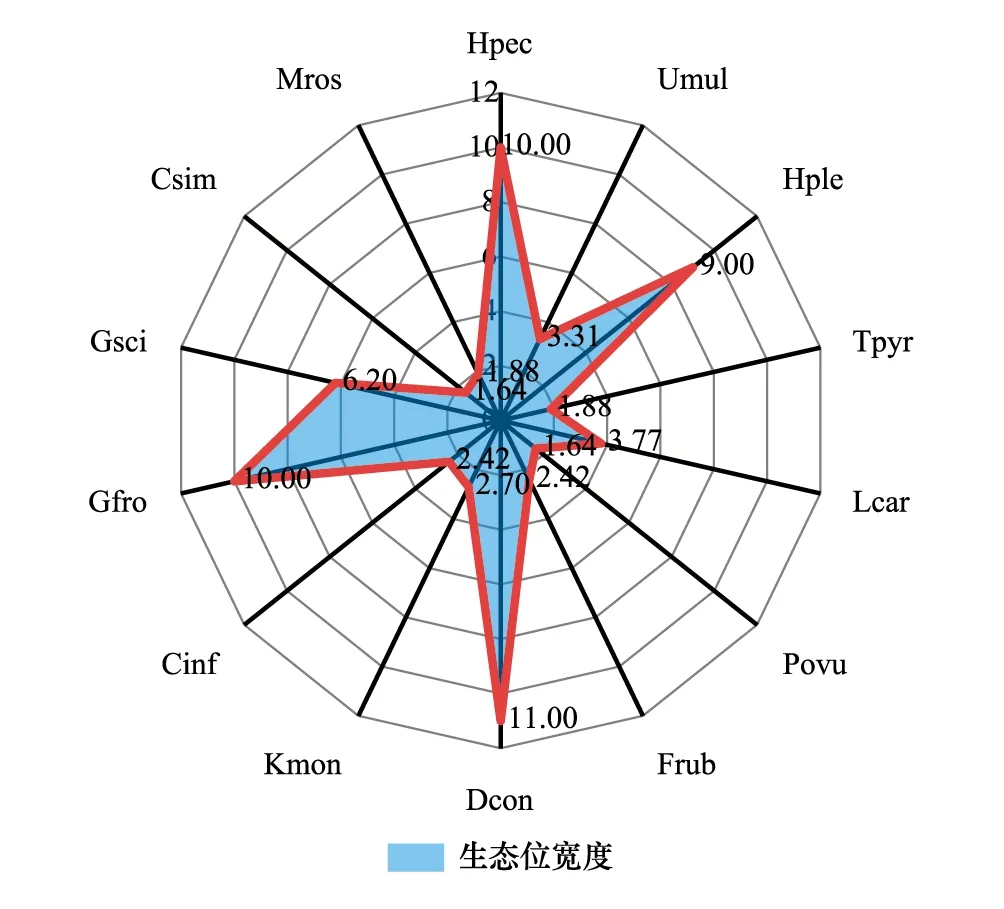
图3 拉萨河流域亏祖山土壤纤毛虫优势种生态位宽度Fig.3 Niche breadth of dominant species of soil ciliates on Kuizu Mountain in the Lhasa River Basin
在空间维度上,拉萨河流域亏祖山土壤纤毛虫优势种生态位宽度位于1.64—11之间(图3),生态位变化幅度较大,其中生态位宽度最大的是明显长颈虫(Dileptusconspicuus),生态位宽度为11;其次为点滴半眉虫(Hemiophryspectinata)和前口瞬目虫(Glaucomafrontata),生态位宽度为10,这类物种对环境适应能力较强,有着较广的生态位适应幅度。而生态位宽度最小的是卵圆前管虫(Prorodonovum)和似肾形虫(Colpodasimulans),生态位宽度为1.64,这类物种对生境有严格的要求,对环境具有极强的选择性,需在适宜的条件下才能利用资源。
2.2.2土壤纤毛虫优势种生态位重叠值
在空间维度上,拉萨河流域亏祖山土壤纤毛虫优势种生态位重叠值位于0.02—0.99之间(图4),生态位重叠值最低的是似肾形虫(Colpodasimulans)和卵圆前管虫(Prorodonovum),而生态位重叠值最高的是念珠角毛虫(Keronopsismonilata)和膨胀肾形虫(Colpodainflata)。土壤纤毛虫14个优势种的91对生态位重叠值中,有11对生态位重叠值较高(Oik>0.6),占总对数的12.09%,有38对生态位重叠值低(0.3 图4 拉萨河流域亏祖山土壤纤毛虫优势种生态位重叠值Fig.4 Niche overlap value of the dominant species of soil ciliates on Kuizu Mountain in the Lhasa River Basin 2.2.3土壤纤毛虫优势种生态响应速率 分析拉萨河流域亏祖山土壤纤毛虫优势种相对资源占用量(ΔOik)和响应速率(R)结果显示(表3),卵圆前管虫(Prorodonovum)的相对资源占用量(Oik)的变化幅度(绝对值)最大,而美丽圆纹虫(Furgasoniarubens)相对资源占用量变化幅度(绝对值)最小。卵圆前管虫(Prorodonovum)有最大的负△Oik,生长空间缩小,呈衰退状态,美丽圆纹虫(Furgasoniarubens)呈衰退状态。明显长颈虫(Dileptusconspicuus)有最大的正△Oik,表明其发展的空间较大,与其他优势种空间资源利用竞争较强烈,在竞争中处于优势位置,而闪瞬目虫(Glaucomascintillans)发展的空间略小于其他优势种。生态响应速率结果可反映空间维度上优势种对生境环境的生态响应状况,其中美丽圆纹虫(Furgasoniarubens)的衰退趋势较强烈(R=-3.102),卵圆前管虫(Prorodonovum)的衰退的趋势较弱(R=-0.019),闪瞬目虫(GlaucomaScintillans)的发展的趋势较为强烈(R=2.184),前口瞬目虫(Glaucomafrontata)发展的趋势较弱(R=0.070)。从总体上来看,拉萨河流域亏祖山土壤纤毛虫优势种在空间维度上生态响应速率之和为负数(-1.57),说明土壤纤毛虫优势种处于衰退阶段。 表3 土壤纤毛虫优势种相对资源占用量(ΔOik)和响应速率(R)Table 3 Relative Resource Occupation(ΔOik) and Response Rate (R) of the dominant Species of soil ciliates 土壤纤毛虫在不同环境因子的影响下发生生态位分化,通过排序的手段来解释物种-环境因子间的关系,从而对生态位测度计算得到的结果进行补充说明。首先对拉萨河流域亏祖山土壤纤毛虫优势种DCA 分析,优势种的分布可使用单峰模型,共筛选6 个环境因子海拔(Alt)、土壤温度(ST)、pH、总氮(TP)、全磷(TP)和有机质含量百分比(SOM)进行CCA 分析,应用蒙特卡拟合方法对环境因子进行显著性检验。结果显示(图5),总氮(TN)(F=1.9,P=0.034)是显著解释变量,对优势种变异的解释率为17.6%,是影响拉萨河流域亏祖山土壤纤毛虫优势种分布及生态位的主要环境因子,影响不显著的环境因子用虚线表示,结果如图5所示。第1 排序轴和第2 排序轴均与总氮(TN)呈正相关。TN与Frub、Mros、Umul、Csim、Tpyr和Hple呈正相关,而与Gsci、Povu、Kmon、Cinf、Dcon、Hpec、Gfro及Lcar及呈负相关。轴1和轴2对优势种方差的解释率分别为21.81%和16.02%,两轴累计解释物种变异的37.83%,表明轴1和轴2能较好地反映优势种与环境因子的关系。TP聚集在轴1的正轴,与轴1呈最大正相关;Hple、Umul、Lcar和Csim等与TP呈正相关,说明TP是影响其分布的因素;Alt、SOM和pH聚集在轴2的负轴,与轴2呈最大负相关。ST聚集在轴2的正轴,与轴2呈最大正相关,Tpyr、Frub和Mros等与ST呈正相关,说明ST是影响其分布的因素。 图5 亏祖山土壤纤毛虫优势种与环境因子的典范对应分析(CCA)Fig.5 The Canonical correspondence analysis (CCA) of dominant species soil ciliates and environmental factors on Kuizu Mountain Alt:海拔 Altitude;ST:土壤温度 Soil temperature;TN:总氮 Total nitrogen;TP:全磷 Total phosphorus;SOM:有机质 Soil organic matter 生态位宽度反映物种占有空间资源的多少、空间分布范围和状况以及均匀程度,是衡量物种对其所在环境的利用程度[31—32]。土壤纤毛虫群落组成出现差异,生态位宽度使土壤纤毛虫出现可利用资源谱的变化[33]。拉萨河流域亏祖山为自然放牧地,每个区域的放牧强度也有所差异,海拔较小的区域,距离村民居住地较近,牲畜及人为活动较为频繁,鼠兔的分布也明显比海拔大的区域多,水源的分布主要集中在低中 (3900—4500 m)海拔段,饮水点附近是高寒草地牲畜干扰较强的区域,因此导致各样点存在不同的小生境,空间资源分布不均匀,土壤纤毛虫优势种的生态位宽度不同。生态位宽度最大的是明显长颈虫(Dileptusconspicuus)、点滴半眉虫(Hemiophryspectinata)和前口瞬目虫(Glaucomafrontata),表明其数量较多,且在各样点中出现的频率高,表明其利用空间资源能力强,对环境适应能力强,有较强的生态适应能力。生态位宽度最小的是卵圆前管虫(Prorodonovum)和似肾形虫(Colpodasimulans),其数量相对较少,分布范围较小,占用空间资源较少,对环境的选择具有一定依赖性,空间资源利用能力较差。亏祖山土壤纤毛虫生态位宽度较宽的种间相互竞争激烈且更趋向于生境特化,可以作为土壤的指示物种,利用同一物种在不同营养状态下生态位宽度变化的特性,将物种作为土壤的指示种具有一定的意义。 生态位重叠值反映了物种之间利用资源或对环境适应能力的相似程度及种间竞争关系,体现了物种对资源的利用状况和物种间分布地段的交错程度[34—36],在生物群落中,生态位通常表现为倾向于分享其他物种的基础生态位部分,结果导致两个或者更多的物种对资源有共同需求,这些有着相似的生态特性的物种会共同利用同样的生境资源,土壤纤毛虫群落会发生显著的生态位重叠特征[37—38]。拉萨河流域亏祖山土壤纤毛虫生态位重叠值低(0.3 群落呈现发展或者衰退趋势是由于不同物种间竞争的直接结果,它是生物或者非生物因子(环境因子)相互作用的生态响应[42]。拉萨河流域亏祖山土壤纤毛虫生态位宽度大的物种,其相对资源占用量(Oik)越大,往往具有较广阔的生态资源幅度,具有较大的发展空间,大多数情况下处于发展的趋势,如明显长颈虫(Dileptusconspicuus)、点滴半眉虫(Hemiophryspectinata)和前口瞬目虫(Glaucomafrontata)属于发展型类群。而生态位宽度较小的物种,其对土壤环境适应能力较差,空间资源利用能力不强,往往属于衰退型种群,这与安瑞志等研究结果一致[43],如卵圆前管虫(Prorodonovum)和似肾形虫(Colpodasimulans)属于衰退型物种。从生态响应速率来看,发展趋势较强的优势种为了获得更多的生境资源,通过增大生态位宽度来加强自身的生态适应能力,会逐渐代替其他衰退型的土壤纤毛虫,如闪瞬目虫(Glaucomascintillans);而衰退趋势较强的土壤纤毛虫,如美丽圆纹虫(Furgasoniarubens)等将会逐渐消亡。拉萨河流域土壤纤毛虫优势种之间以及优势种和环境之间未达到相对的动态平衡,其优势种正处衰退阶段。 土壤动物与其生存的环境之间有着相互关系,生理活动促进营养元素在凋落物和土壤系统中的循环影响土壤理化性质,而理化性质变化会影响土壤动物的分布、数量等特征[44—45]。拉萨河流域亏祖山土壤纤毛虫优势种与环境因子的典范对应分析(CCA)结果显示,轴1和轴2能较好地反映优势种与环境因子的关系。排序轴1反映了全磷的变化;排序轴2反映了pH、土壤温度、总氮、海拔和土壤有机质含量的变化,对土壤纤毛虫产生直接影响。因此,排序轴2是影响土壤纤毛虫的空间生态位分化的主要因子,而排序轴1是影响其空间生态位分化的次要因子。TN为显著解释性变量,对优势种变异的解释率为17.6%,本研究发现,土壤中总氮的含量能够保证土壤纤毛虫生存所需的营养,多数土壤纤毛虫对土壤总氮表现出明显的负向响应,这一结果与杨大星等研究一致[46],一定量的土壤养分物质是土壤纤毛虫生存所必须的,而含量过高则会产生负面的效应[47—49]。由于土壤纤毛虫迁移能力不强,过多的氮含量对部分土壤纤毛虫的群落数量特征产生明显的抑制作用。总氮是影响土壤纤毛虫优势种变异的主要驱动因子,其他理化因子不是主要影响因素。CCA结果显示拉萨河流域亏祖山土壤纤毛虫优势种受到多种环境因子的共同作用,土壤纤毛虫优势种与其生存的特殊生境环境之间形成了适应性机制,每种因子对纤毛虫种产生不同的效应,其综合影响导致不同海拔土壤纤毛虫优势种分布具有一定差异。 亏祖山不同海拔高寒草甸闪瞬目虫(Glaucomascintillans)和龙骨漫游虫(Litonotuscarinatus)为绝对优势种。明显长颈虫(Dileptusconspicuus)、点滴半眉虫(Hemiophryspectinata)以及前口瞬目虫(Glaucomafrontata)三个优势种对土壤环境适应能力较强,有较广的生态适应幅度;卵圆前管虫(Prorodonovum)和似肾形虫(Colpodasimulans)对土壤环境变化适应较为敏感,其适应能力较弱。土壤纤毛虫优势种生态位重叠值低,种间竞争弱。生态响应速率之和为负值,表明土壤纤毛虫优势种之间以及优势种和环境之间未达到相对的动态平衡,优势种正处衰退阶段;总氮是影响拉萨河流域土壤纤毛虫优势种分布与生态位的主要环境因子。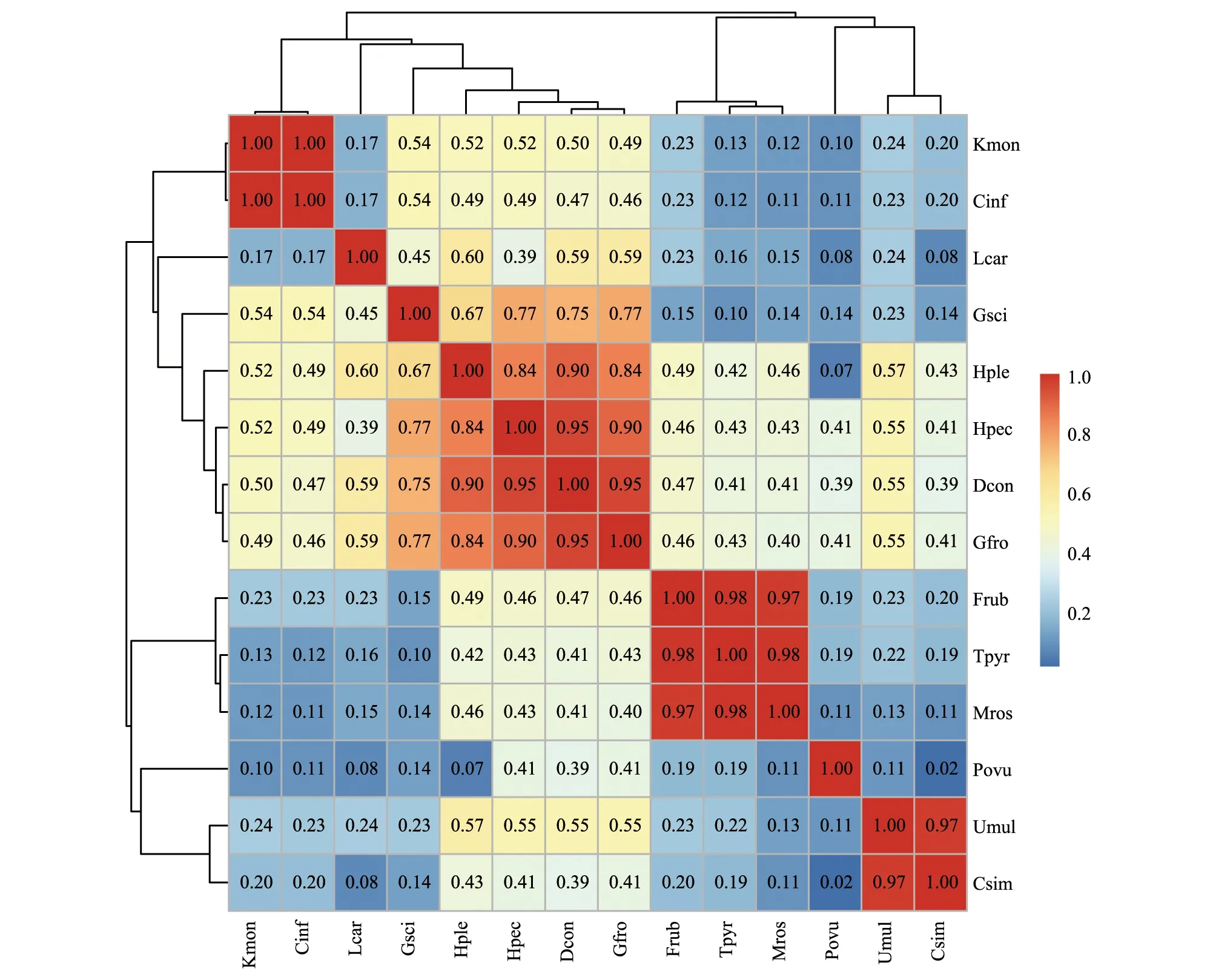

2.3 拉萨河流域亏祖山土壤纤毛虫优势种与环境因子的关系

3 讨论
3.1 拉萨河流域亏祖山土壤纤毛虫优势种生态位
3.2 拉萨河流域亏祖山土壤纤毛虫优势种环境因子的关系
4 结论
