萝卜乙烯合成途径基因的鉴定及对胁迫的响应
2022-05-24刘同金徐铭婕崔群香张爱慧包崇来王长义
刘同金,徐铭婕,崔群香,张爱慧,包崇来,王长义*
(1 金陵科技学院 园艺园林学院, 南京 210038;2 浙江省农业科学院 蔬菜研究所, 杭州 310021)
乙烯是一种重要的气体植物激素,广泛存在于植物的各种组织和器官。乙烯在植物生长发育和环境适应过程中发挥重要作用,不仅参与种子萌发、根毛发育、抽薹开花、果实软化和成熟、器官衰老和脱落等植物生长发育过程[1],也参与了植物对细菌、真菌、病毒、线虫和昆虫等生物胁迫的响应[2],及机械损伤、低氧、冷害和冻害、水分胁迫、盐胁迫等非生物胁迫的响应[3-4]。因此,乙烯的内源产生和信号转导途径得到研究者的广泛关注和深入研究。
乙烯生物合成起始于甲硫氨酸,在S-腺苷甲硫氨酸合成酶(S-adenosylmethionine synthesis,SAMS)催化下生成S-腺苷甲硫氨酸(SAM)。SAM是植物体内乙烯、精胺/亚精胺等生物合成途径主要的甲基供体,由ACC合酶(ACC synthesis,ACS)催化生成氨基环丙烷羧酸(1-aminocyalopropane-1-carboxylate,ACC)和5′-甲硫酰苷,前者进行乙烯合成,而后者则进入甲硫氨酸循环途径重新合成甲硫氨酸。ACC在ACC氧化酶(ACC oxidase,ACO)的催化下产生乙烯[1]。植物中SAMS、ACS和ACO均由多基因家族编码。
SAMS也被称为蛋氨酸腺苷转移酶(methionine adenosyltransferase,MAT),乌拉尔图小麦(Triticumurartu)、水稻(OryzasativaL.)和高粱[Sorghumbicolor(L.) Moench]基因组中各有3个MAT,拟南芥(Arabidopsisthaliana)、番茄(Lycopersiconesculentum)、茄子(SolanummelongenaL.)和大麦(HordeumvulgareL.)基因组中各有4个MAT,而蒺藜苜蓿(Medicagotruncatula)、向日葵(HelianthusannuusL.)和大豆[Glycinemax(Linn.) Merr.]基因组中分别有5、7和9个MAT基因[5]。研究表明MAT基因参与了植物对多种胁迫的响应。低温、ABA、H2O2和NO等非生物胁迫显著诱导紫花苜蓿MfSAMS1上调表达,在烟草中过表达该基因显著提高了其耐寒性[6]。番茄中MAT不同成员对非生物胁迫和外源激素处理的响应不同[5]。
ACS是乙烯生物合成的限速酶,ACO催化乙烯生物合成的最后一步,二者的活性影响植物体内乙烯的含量。拟南芥基因组上分别有12个ACS基因(AtACS1-12)[7]和5个ACO基因(AtACO1-5)[8]。AtACS3是假基因,ACS10和ACS12特异的对天冬氨酸和芳环氨基酸进行氨基转移,其余9个AtACS具有催化ACC生物合成的功能[7]。研究表明,ACS和ACO基因在植物生长发育、抵抗生物和非生物胁迫过程中起重要作用。梨(Pyrusspp.)基因组中分别有13个ACS和11个ACO基因,其中2个ACS和3个ACO在果实中不表达,4个ACS和3个ACO被乙烯利诱导显著上调表达,且1-甲基环丙烯(1-MCP)显著抑制其表达[9]。乙烯利、茉莉酸甲酯、水杨酸和低温显著诱导匍匐翦股颖ACO上调表达,而干旱和NaCl处理抑制其表达[10]。过表达ACO显著提高了转基因拟南芥的耐水淹能力[11]。
萝卜是中国重要的蔬菜作物,其生长发育过程中可能遭受营养元素缺乏、病虫危害、极端温度等生物和非生物胁迫。乙烯及其参与的信号转导途径在植物品质形成、抵抗多种生物和非生物胁迫过程中起重要作用[11]。本研究在全基因组水平对萝卜乙烯合成途径MAT、ACS和ACO基因家族成员进行鉴定,分析其启动子包含的顺式作用元件,并分析其在萝卜不同组织中的表达及对生物和非生物胁迫的响应,为其生物学功能的解析奠定基础。
1 材料和方法
1.1 材 料
以‘北京心里美’萝卜种子为供试材料,经10%次氯酸钠消毒后置于28 ℃恒温箱中避光催芽24 h。出芽后播种于10×10 cm育苗钵,于光照培养箱(16 h光照,22 ℃;8 h黑暗,20 ℃)中培养至三叶一心期,分别浇灌1.5% NaCl、20% PEG、2% 葡萄糖、2%果糖和2%蔗糖溶液,以蒸馏水处理为对照。每个处理8株,3次生物学重复。处理3 h后用蒸馏水将根系洗净,擦干后于液氮速冻后置于-80 ℃冰箱中保存。
1.2 方 法
1.2.1MAT、ACS和ACO基因家族成员鉴定与蛋白理化性质分析由TAIR网站(http://arabidopsis.org/)下载拟南芥MAT、ACS和ACO基因家族成员的核苷酸序列,在XYB36-2萝卜基因(https://www.ncbi. nlm.nih.gov/assembly/GCA_002197605.1/)[12]中执行本地BlastN(E-value=10-5),获得萝卜MAT、ACS和ACO基因家族成员的候选序列;将其在TAIR 数据库中进行BlastN搜索,最终筛选分别获得萝卜MAT、ACS和ACO基因家族成员序列。利用ExPASy(http://web.expasy.org/protparam/)对萝卜MAT、ACS和ACO蛋白的分子量、等电点等理化性质进行预测和分析。
1.2.2 染色体定位分析根据XYB36-2萝卜参考基因组注释文件确定MAT、ACS和ACO基因的染色体位置信息,使用MapInspect软件进行染色体定位作图。
1.2.3 系统发育树构建拟南芥MAT、ACS和ACO蛋白质序列由TAIR网站(http://arabidopsis.org/)下载,水稻MAT、ACS和ACO蛋白质序列下载于MBKbase网站(http://mbkbase.org/R498/)[13],并根据Heidari等[5]、Houben等[8]和Xu等[14]报道进行命名。使用MEGA5.05的ClustalW进行序列比对,邻接法(NJ,Neighbor-Joining)构建系统发育树,Bootstrap设为1 000次。
1.2.4 启动子顺式作用元件分析利用TBtools软件(https://github.com/CJ-Chen/TBtools/releases)[15]提取萝卜MAT、ACS和ACO基因起始密码子上游1 500 bp序列作为启动子区域,通过PlantCARE网站(http://bioinformat ics.psb.ugent.be/webtools/plantcare/html/)的Search for CARE工具进行顺式作用元件的预测。
1.2.5MAT、ACS和ACO基因转录组表达分析根据前人发表的XYB36-2萝卜芽期、破肚期、膨大前期、膨大盛期和成熟期共5个不同发育时期的肉质根和5个不同组织(薹、愈伤组织、花、叶和角果)的转录组数据[16-17],分析MAT、ACS和ACO在萝卜不同组织中的表达;利用Tkachenko等[18]报道的抗、感根癌农杆菌萝卜自交系接种根癌农杆菌7 d后下胚轴的转录组数据,分析MAT、ACS和ACO对生物胁迫的响应;利用前人报道的镉胁迫(0.876 mmol/L CdCl2·2.5H2O)、铬胁迫(600 mg/L K2Cr2O7)和铅胁迫[1 000 mg/L Pb(NO3)2]萝卜根的转录组数据,分析MAT、ACS和ACO对重金属胁迫的响应[19-21];利用4 ℃低温处理3 d的萝卜幼苗转录组数据分析MAT、ACS和ACO对冷胁迫的响应[22]。转录组表达数据经log2均一化处理后,利用TBtools软件进行表达热图的绘制[15]。
1.2.6MAT、ACS和ACO基因荧光定量PCR(qRT-PCR)表达分析使用华越洋生物科技有限公司的通用植物RNA提取试剂盒进行总RNA的提取,使用天根生化科技有限公司的FastKing RT Kit(With gDNase)反转录成cDNA。qRT-PCR采用TaKaRa公司的SYBR Green qPCR试剂盒,体系和反应程序参见试剂盒说明书。引物序列见表1,以GAPDH作为内参,生物学和技术重复均为3次,采用2-ΔΔCT方法进行相对定量分析。使用SPSS17.0软件进行显著性分析,并利用Excel绘图。

表1 本实验所用引物
2 结果与分析
2.1 萝卜MAT、ACS和ACO基因家族成员鉴定与蛋白理化性质分析
利用生物信息学方法,在萝卜基因组中分别鉴定出8个RsMAT、16个RsACS和7个RsACO基因(表2),其CDS序列全长分别为1 173~1 182、1 017~1 656和933~966 bp,分别编码390~393、338~551和310~321个氨基酸,RsMAT和RsACO基因序列长度比较保守,RsMAT和RsACS序列长度显著大于ACO。RsMAT、RsACS和RsACO基因家族成员蛋白分子量分别为42.54~43.2、38.58~60.03和35.40~36.47 kD,理论等电点分别为5.36~6.08、5.19~8.21和4.96~7.83,亲水性分别为-0.333~-0.243、-0.361~0.122和-0.575~-0.459,脂肪指数分别为80.08~84.05、81.11~91.88和74.04~81.03。
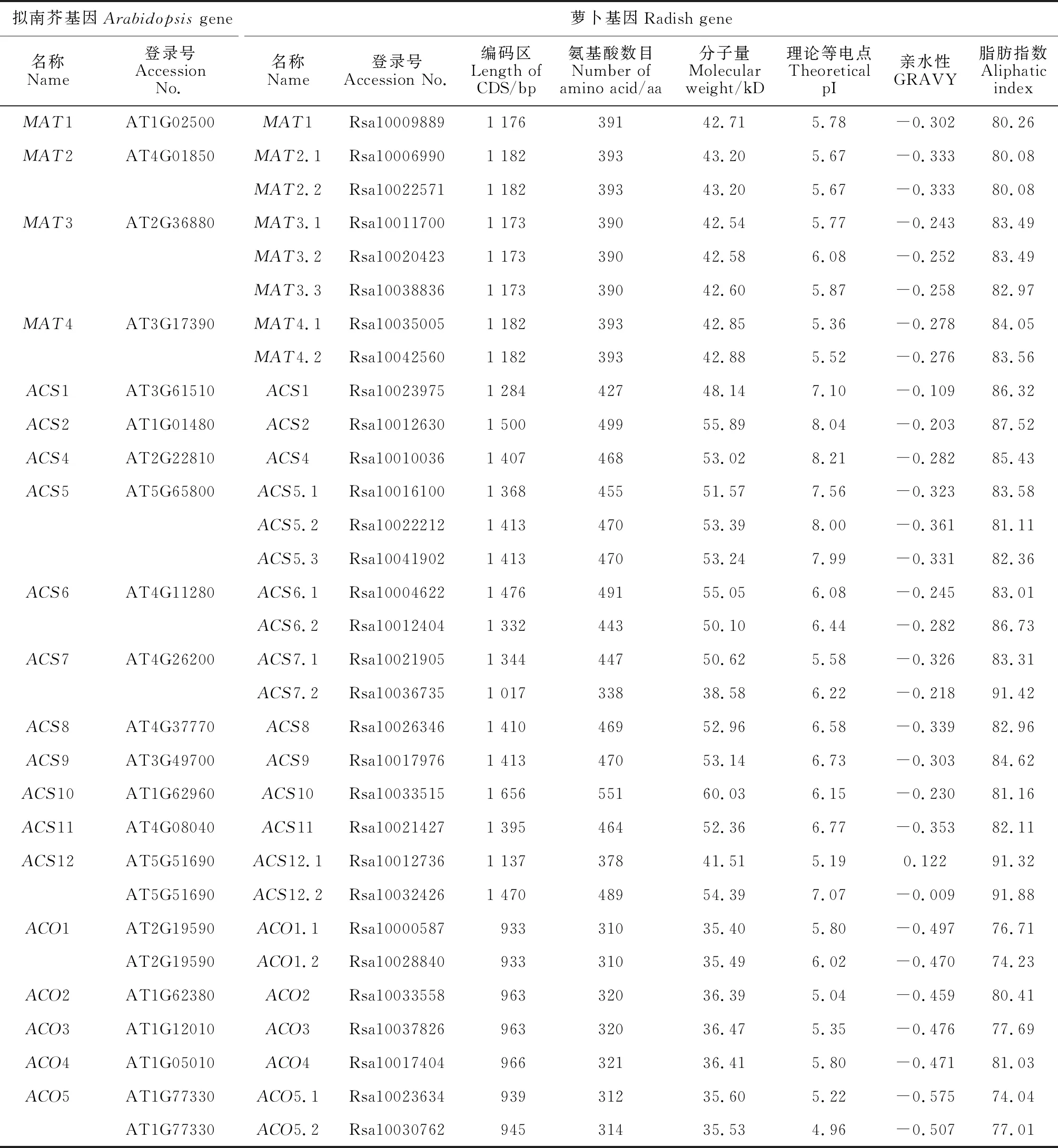
表2 萝卜与拟南芥MAT、ACS和ACO基因家族成员信息
2.2 萝卜MAT、ACS和ACO基因的染色体定位
参考XYB36-2基因组的注释信息将乙烯生物合成途径的30个基因定位于萝卜的8条染色体,1个基因(RsACO1.1)定位于Scaffold2443(图1)。它们在染色体上的分布十分不均匀,1、2和9号染色体上各分布有5个,3和4号染色体各分布有4个,5和6号染色体各有3个,7号染色体上有1个,而8号染色体上无MAT、ACS和ACO分布。染色体定位分析(图1)未发现萝卜MAT、ACS和ACO基因存在串联重复,其基因数目的增加是由于萝卜基因组多倍化的结果。
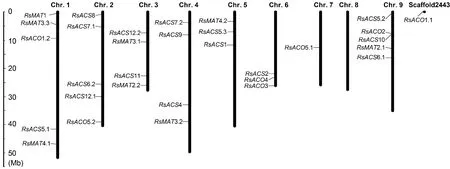
左侧比例尺表示染色体长度图1 萝卜MAT、ACS和ACO基因在染色体上的分布The scale on the left provides the relative length of chromosomeFig.1 The distribution of MAT, ACS and ACO genes on radish chromosomes
2.3 萝卜MAT、ACS和ACO的系统进化分析
利用萝卜(Rs)、拟南芥(At)和水稻(Os)的MAT、ACS和ACO蛋白质序列构建系统进化树,结果可将其分别分成3类。MAT家族第Ⅰ类群包括3个RsMAT、2个AtMAT和全部的OsMAT,第Ⅱ类群为AtMAT4、RsMAT4.1和RsMAT4.2,第Ⅲ类群为AtMAT3及其在萝卜中的3个同源基因(图2,A)。萝卜、拟南芥和水稻ACS(图2,B)以及ACO(图2,C)家族成员在3个类群中均有分布。ACS和ACO的编码基因在3个物种虽然基因数目有较大差异,但在3个类群中均有成员分布,表明ACS和ACO进化上比MAT保守,可能不同类群的成员在植物生长发育过程中发挥不同的功能。

图2 萝卜(Rs)、拟南芥(At)和水稻(Os)的MAT(A)、ACS(B)和ACO(C)系统进化树Fig.2 Phylogenetic tree for MAT (A), ACS (B) and ACO (C) of radish (Rs),Arabidopsis thaliana (At) and rice (Os)
2.4 萝卜MAT、ACS和ACO基因的启动子顺式作用元件分析
为了解萝卜MAT、ACS和ACO基因的表达调控机制,对其启动子序列进行顺式作用元件预测,3个基因家族分别发现39、41和38类顺式作用元件(图3)。萝卜MAT、ACS和ACO基因启动子中含有23种与光响应相关的顺式作用元件。除RsACO1.1和RsACO3,其余基因启动子中均至少含有一种响应植物激素的顺式作用元件。本研究还发现萝卜乙烯生物合成途径基因启动子中存在多种响应生物及非生物胁迫的顺式作用元件,如响应脱水、低温和干旱的DRE元件,响应低温的LTR和干旱的MBS元件,厌氧诱导的ARE和GC-motif元件,以及参与抗性和胁迫响应的TC-rich repeats。推测萝卜乙烯合成途径基因参与了其对生物和非生物胁迫的响应。此外,部分基因启动子区还有ATBP-1和MYBHv1结合位点,分生组织、种子和胚乳特异表达的顺式作用元件,及响应昼夜节律和细胞周期的元件,表明萝卜部分MAT、ACS和ACO基因表达可能具有时空特异性。
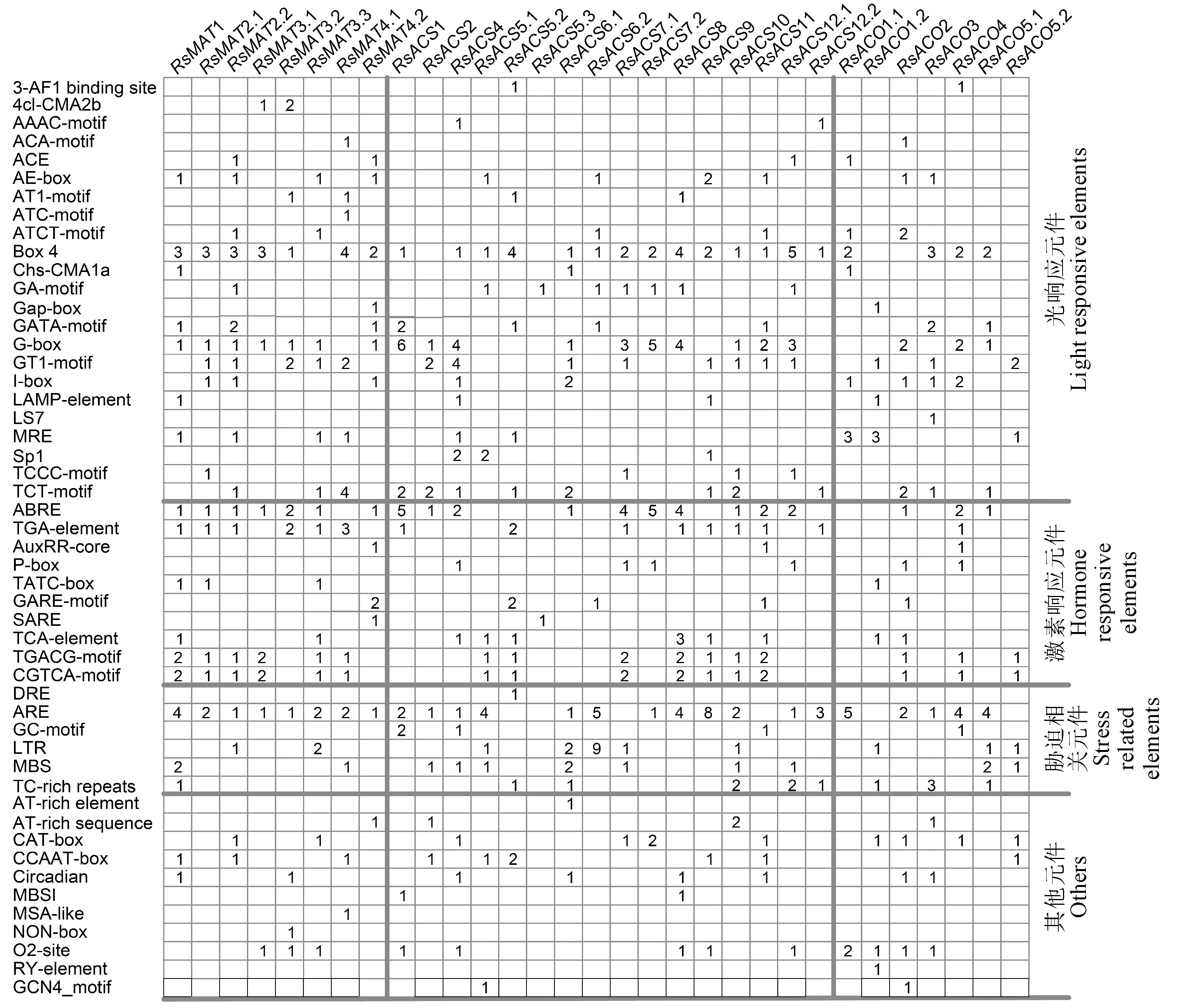
图3 萝卜MAT、ACS和ACO基因启动子区域顺式作用元件的种类及数目Fig.3 The types and number of cis-acting elements in the promoter regions of the MAT,ACS and ACO genes in radish
2.5 萝卜MAT、ACS和ACO基因的组织表达分析
利用转录组数据对其在肉质根的5个不同发育时期,以及叶片、薹、花、角果和愈伤组织中的表达进行了分析,根据表达趋势进行聚类分析将31个乙烯合成途径基因分为3类:第一类包含萝卜MAT2.1、MAT2.2、MAT3.1、MAT3.2、MAT3.3、MAT4.1、MAT4.2、ACS6.1和ACO2共9个基因,这些基因在不同组织中的表达量均较高;第二类包含萝卜ACS1、ACS2、ACS4、ACS5.1、ACS5.2、ACS5.3、ACS7.1、ACS7.2、ACS8、ACS9、AACS11、ACS12.2和ACO1.2共13个基因,这些基因在不同组织中的表达量均较低;第三类包含萝卜MAT1、ACS6.2、ACS10、ACS12.1、ACO1.1、ACO3、ACO4、ACO5.1和ACO5.2共9个基因,这些基因呈现组织或发育时期特异性表达(图4)。
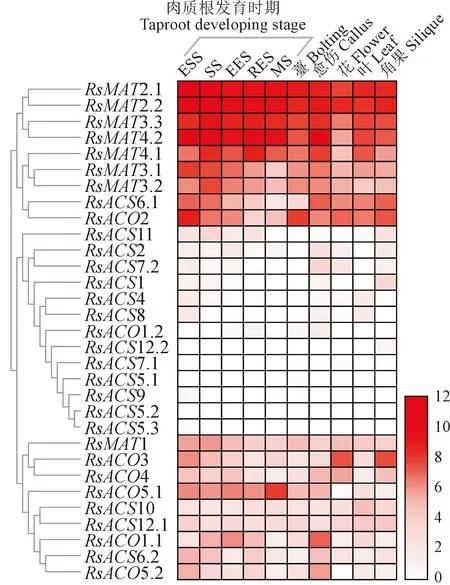
ESS.芽期;SS.破肚期;EES.膨大前期;RES.膨大盛期;MS.成熟期。图例表示标准化的FPKM值。红色表示高表达水平,白色表示低表达水平,下同图4 萝卜MAT、ACS和ACO基因的组织表达模式ESS. Seedling stage; SS. Splitting stage; EES. Early expanding stage; RES. Rapid expanding stage; MS. Mature stage. Legends represent the normalized FPKM values. Red means high expression level, while white means low expression level, the same as belowFig.4 The expression profiles of radish MAT, ACS and ACO gens in various tissues
2.6 萝卜MAT、ACS和ACO基因对生物胁迫的响应
为明确萝卜MAT、ACS和ACO基因对生物胁迫的响应,利用转录组数据比较了其在抗、感根癌农杆菌萝卜高代自交系接种A.tumefaciens7 d后下胚轴中的表达情况(图5)。接种后,萝卜MAT基因在抗病材料中除MAT1外均上调表达,而感病材料除MAT4.1和MAT4.2外均下调表达。抗病萝卜ACS2、ACS7.1和ACS7.2受A.tumefaciens侵染显著上调表达;而感病材料ACS4、ACS6.2、ACS8和ACS11显著上调表达。抗、感萝卜RsACO基因对A.tumefaciens侵染的响应不同,抗病材料ACO1.1、ACO1.2和ACO4受其侵染显著上调表达,其余基因的表达水平无显著变化;而感病材料中除ACO5.2在其侵染前后差异不显著、ACO5.1显著上调表达外,其余成员均显著下调表达。
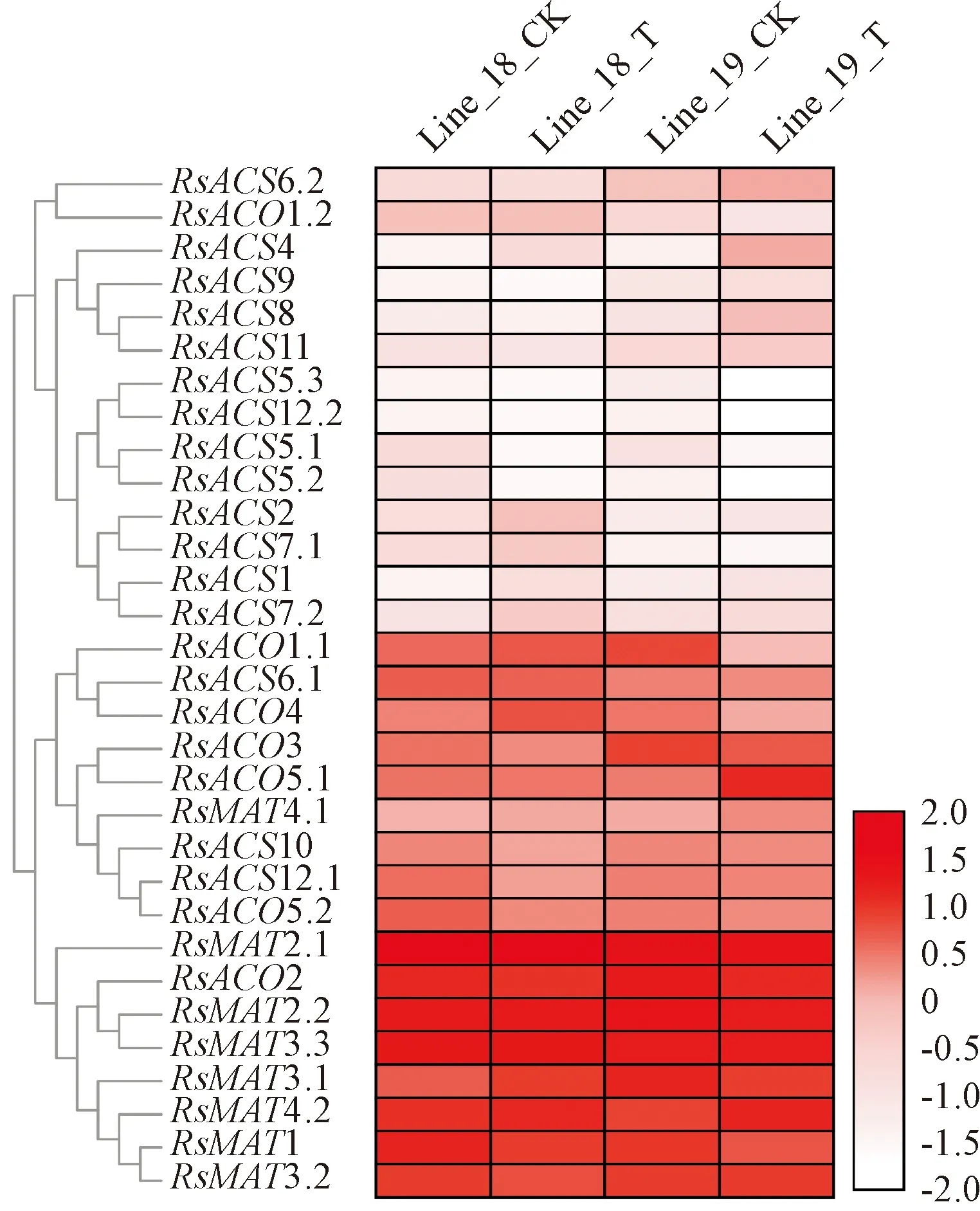
Line_18和Line_19分别为抗和感根癌农杆菌萝卜高代自交系。CK和T分别表示接种LB培养基和根癌农杆菌7 d后的样品图5 抗和感根癌农杆菌萝卜接种根癌农杆菌7 d 后下胚轴MAT、ACS和ACO基因的表达Line_18 and Line_19 is resistance and susceptible radish inbred lines to A. tumefaciens, respectively. CK and T represent 7 d after incubated with LB medium and A. tumefaciens, respectivelyFig.5 The expression profiles of MAT, ACS and ACO genes in 7 d after incubated with A. tumefaciens in radish hypocotyls
2.7 萝卜MAT、ACS和ACO基因对非生物胁迫的响应
图6显示,重金属胁迫均显著抑制RsMAT1、RsACO5.1的表达,促进RsMAT4.2、RsACO1.1和RsACO4的表达;铅和镉处理诱导RsMAT3.2和RsACO2上调表达,但铬诱导其下调表达;此外,镉显著诱导RsACS6.1、铅显著诱导RsACO3上调表达(图6,A)。4 ℃低温处理显著抑制RsMAT2.1、RsMAT2.2、RsMAT4.1、RsACO1.1和RsACO5.2的表达(图6,B)。
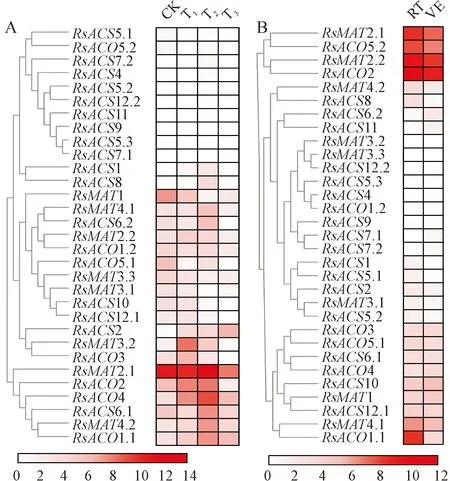
CK.对照;T1.铅胁迫;T2.镉胁迫;T3.铬胁迫;RT.室温处理;VE.4 ℃处理3 d图6 重金属(A)和低温(B)胁迫对萝卜MAT、ACS和ACO基因表达的影响CK. Control; T1. Lead stress; T2. Cadmium stress; T3. Chromium stress; RT. Room temperature treatment; VE. 4 ℃ for 3 d treatmentFig.6 The expression profiles of MAT, ACS and ACO genes in radish with heavy metal (A) and low temperature (B) stress
同时,选取转录组分析获得的响应生物和非生物胁迫显著上调或下调表达的11个基因,通过qRT-PCR方法研究其对1.5% NaCl和20% PEG-6000两种非生物胁迫的响应(图7)。所有样品中均未检测到RsMAT1表达,其余基因响应一种或多种处理表达量发生显著变化。NaCl和PEG-6000处理均显著抑制RsMAT4.1和RsACO4的表达,但显著促进RsACO5.1和RsACO5.2的表达。此外,NaCl处理还诱导RsMAT2.2、RsMAT4.2、RsACS2和RsACS6.1显著上调表达,RsACO1.1显著下调表达,但对RsMAT2.1表达无显著影响;PEG-6000处理还显著抑制RsMAT2.1和RsMAT4.2的表达。
另外,为了解糖是否参与对上述差异表达的基因的调控,利用qRT-PCR分析了葡萄糖、果糖和蔗糖处理对上述差异表达基因的影响,发现3种糖处理均显著抑制RSMAT2.1、RSMAT2.2、RSMAT4.1和RSMAT4.2的表达;均诱导RSACO5.1和RSACO5.2显著上调表达;但对RsACS2和RsACS6.1表达无显著影响(图8)。此外,葡萄糖和果糖处理显著促进RSACO1.1的表达,但蔗糖对其表达无显著影响;果糖显著促进、蔗糖显著抑制RSACO4的表达,但葡萄糖对其表达无显著影响。
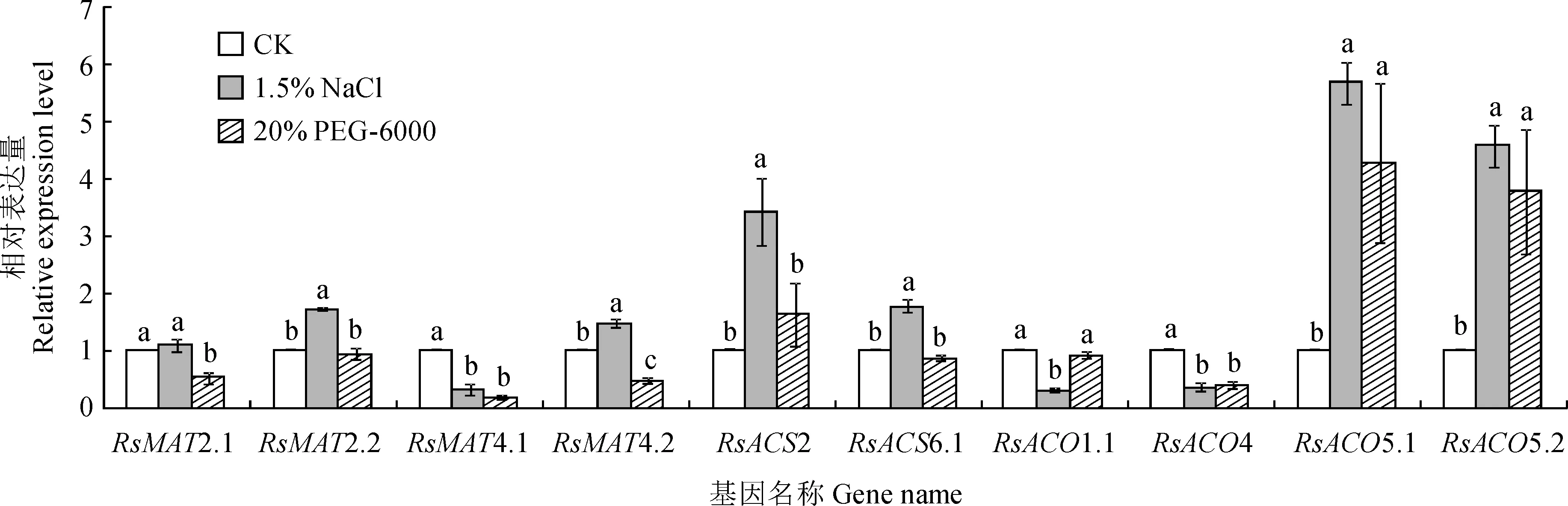
不同小写字母表示基因表达量在处理间存在显著差异(P<0.05),下同图7 萝卜MAT、ACS和ACO基因响应NaCl和PEG-6000胁迫的qRT-PCR分析Different lowercase letters indicate significant difference in gene expression among treatments (P<0.05), the same as belowFig.7 The expression profiles of MAT, ACS and ACO genes in radish response to NaCl and PEG-6000 treatment by qRT-PCR
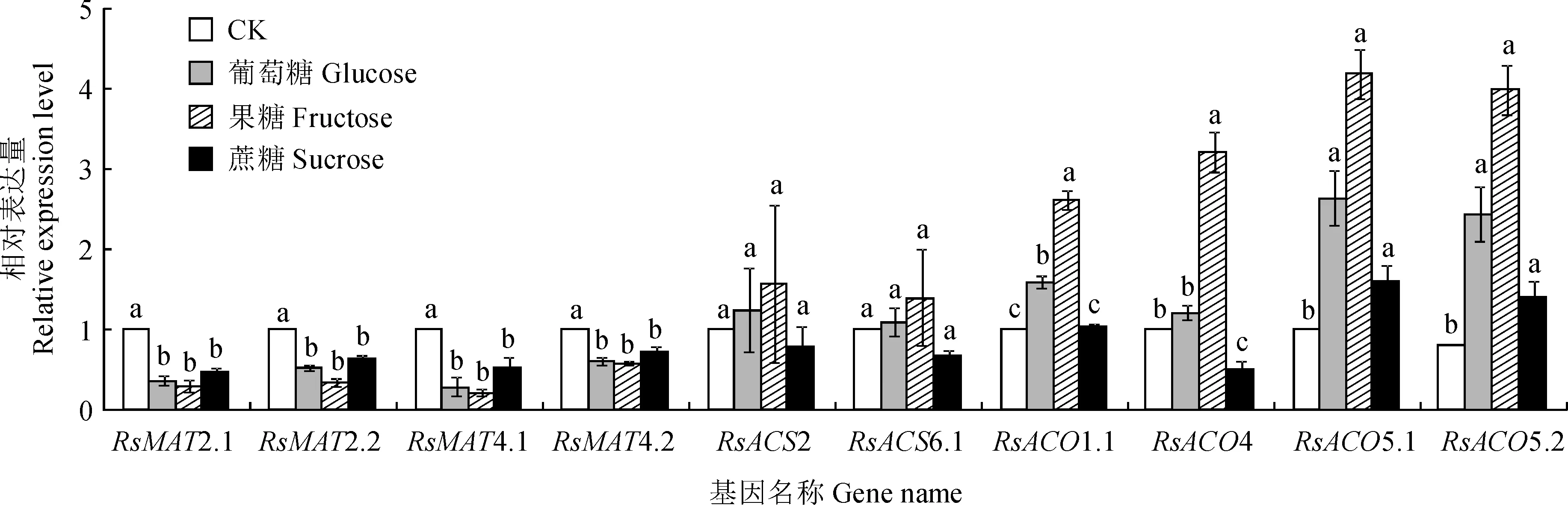
图8 萝卜MAT、ACS和ACO基因响应葡萄糖、果糖和蔗糖处理的qRT-PCR分析Fig.8 The expression profiles of MAT, ACS and ACO genes in radish response to glucose,fructose and sucrose treatments by qRT- PCR
3 讨 论
本研究基于XYB36-2萝卜参考基因组数据[12],采用生物信息学分析方法,在萝卜基因组中共鉴定出8个RsMAT、16个RsACS和7个RsACO基因。RsMAT基因家族成员数目多于乌拉尔图小麦、水稻和高粱(各3个)、拟南芥、番茄、茄子和大麦(各4个)、向日葵(7个)和蒺藜苜蓿(5个),但少于大豆(9个)[5];RsACS基因家族成员数目多于黄瓜(8个)[23]、甜瓜(11个)[24]、拟南芥(12个)[7]、梨(13个)[9]和香蕉(MusananaLour.,14个)[25],但少于苹果(MaluspumilaMill.,19个)[26];RsACO基因家族成员数目多于拟南芥(5个)[8],与番茄、苹果和水稻中数目一致[8],但少于梨(11个)[9]中相应基因家族成员数目。萝卜MAT2、MAT3、MAT4、ACS5、ACS6、ACS7、ACS12、ACO1和ACO5在基因组有2~3个拷贝,而其余基因均为单拷贝,表明萝卜MAT、ACS和ACO基因家族成员在基因组多倍化之后均出现了基因丢失。
本研究发现萝卜MAT、ACS和ACO基因启动子序列中含有与光响应相关的顺式作用元件最多,其次是与植物激素、生物及非生物胁迫响应相关的顺式作用元件。赵薇等[24]发现甜瓜ACS基因启动子序列中,主要含有参与防御和胁迫响应的顺式作用元件和参与响应各种激素的顺式调控元件。Wang等[27]对12种陆生植物ACO基因启动子进行分析发现光响应元件数目最多,推测其可能参与了光调控途径;此外也包含大量的植物激素、生物和非生物胁迫及生长发育相关元件。前人多项研究均表明乙烯与其他植物激素均有交叉作用,共同影响植物生长发育及对生物和非生物胁迫的响应[28],本研究同样发现乙烯合成途径基因启动子含有响应脱落酸、生长素、赤霉素、水杨酸、茉莉酸甲酯的顺式作用元件。上述结果表明,不同植物中乙烯合成途径基因家族成员的表达调控可能具有一定的相似性。本研究发现多数基因的表达和该基因启动子的顺式作用元件的种类有显著关系,如本研究鉴定的31个萝卜乙烯合成途径基因启动子至少含有一个响应光照的顺式作用元件,组织表达分析也发现多个基因在叶片中具有较高的表达量;但二者也并非完全对应,如RsACS4启动子中虽然含有多达16个(9类)参与光响应的顺式作用元件,但该基因在本研究所分析的组织中不表达或表达量很低。
在进化过程中,不同物种的同源基因多数保留了相同或相似的生物学功能,基因的表达模式也往往与其功能密切相关。本研究通过分析萝卜不同组织的转录组数据发现:萝卜MAT2.1/2.2/3.1/3.2/3.3/4.1/4.2、ACS6.1和ACO2在不同组织中的表达量均较高,推测这些基因可能是维持植物正常生长发育所必需的。前人的研究也表明MAT催化甲硫氨酸和三磷酸腺苷反应生成的S-腺苷甲硫氨酸[29],不仅参与植物乙烯合成途径,还是主要的甲基供体以及生物素和多胺合成的前体物质[30],此外还具有转氨基[31]和硫基[32]的作用,是维持植物生长发育所必需的。ACO1.1/3/4/5.1呈组织或发育时期特异性表达,这些基因可能参与植物特定生长发育阶段或特定组织器官的发育。拟南芥ACO1主要在根中表达,其缺失突变体根毛数量显著增加[33],本研究发现其萝卜同源基因在破肚期、膨大前期、膨大盛期表达量较高,可能调控萝卜根毛的发育。RsACO3在花和角果中的表达量最高,RsACO5.1在萝卜肉质根尤其是成熟期表达量最高。研究表明,抑制植物内源ACO表达可抑制乙烯合成而延迟花器官衰老[34],果实成熟过程中普遍伴随乙烯含量的增加,因此推测RsACO3和RsACO5.1可能分别控制花和角果、肉质根乙烯合成,进而调控其衰老和成熟,其具体生物学功能有待进一步的功能验证。
前人研究发现植物中存在大量响应生物胁迫的ERF型转录因子,ERF转录因子通过响应乙烯信号转导途径而调控下游基因的表达,进而提高植物对生物胁迫的抗性[4]。郝丽芬等[35]研究表明,油菜乙烯合成途径基因被黑胫病菌侵染强烈诱导表达。本研究也发现,A.tumefaciens侵染诱导抗根癌农杆菌的萝卜中多数MAT和ACO基因上调表达而感病材料下调表达,表明乙烯合成通路基因在萝卜对生物胁迫的响应过程中发挥重要作用。本研究发现,低温处理显著抑制乙烯生物合成途径基因的表达。前人研究结果也表明,低温处理显著抑制了苹果果实[36]和水稻植株[37]乙烯的合成,过表达乙烯合成基因或外施ACC会提高乙烯含量进而降低植物的耐寒性[4]。邓丽等[38]也发现短时间4 ℃低温抑制溶质型桃果实内源乙烯的释放。本研究还发现同一基因的不同拷贝可能参与不同非生物胁迫的响应。4 ℃低温处理显著抑制RsMAT4.1的表达,重金属胁迫促进了其另一个拷贝RsMAT4.2的表达;重金属胁迫和4 ℃低温处理分别抑制RsACO5.1和RsACO5.2的表达。前人研究也发现,拟南芥MAT4受生物胁迫显著上调表达,而响应非生物胁迫显著下调表达[5];拟南芥ACS基因家族不同成员受不同因素的调节[39-40],与本研究结果一致。前人研究发现镉胁迫通过上调拟南芥ACO2和ACO4、ACS2和ACS6的表达而促进乙烯的合成[41],本研究结果与之一致,但值得注意的是,RsACS6受铅和铬胁迫、RsACO2受铬胁迫显著下调表达。上述基因在植物抵御生物和非生物胁迫中的功能有待深入研究。糖作为一种信号分子和渗透调节物质广泛参与植物对胁迫的响应,低浓度外源蔗糖能缓解干旱对水稻种子的萌发的抑制作用[42],NaCl胁迫增加了流苏幼苗叶片可溶性糖含量[43],外源糖处理显著影响乙烯合成途径基因的表达[44],本研究通过qRT-PCR对发现,除RsMAT2.2外,其余9个基因响应蔗糖和PEG-6000处理的表达趋势一致;除RsMAT2.2和RsACO4外,其余8个基因响应果糖和PEG-6000处理的表达趋势一致。表明果糖和蔗糖可能参与了萝卜对PEG-6000胁迫的响应。
本研究通过对萝卜MAT、ACS和ACO基因的全基因组鉴定及表达分析,为进一步探讨这些基因的生物学功能奠定了基础。
