不同林龄福建柏纯林与混交林生长及养分的差异
2022-05-24李秉钧王希贤荣俊冬陈礼光郑郁善
李秉钧,陈 乾,王希贤,荣俊冬,陈礼光,郑郁善
(福建农林大学 林学院, 福州 350002)
林木器官养分含量是中国林业领域一直以来较为热门的话题,大量的研究已表明[1-5],林木器官养分的动态变化是影响林分生长指标的关键因素,林木各器官养分合理分配也是林木提高自身养分利用效率来适应环境变化和应对养分匮乏的重要机制。而林龄作为指示林木生物量累积的可靠参数,是研究林木生长过程及情况,制定各种经营措施不可缺少的主要参数[6-7]。林分年龄的变化也会使得林木各器官在不同生长时期对养分的需求差异较大[8],不少学者关于林龄变化引起的林木自身各器官养分含量的动态变化开展了一些研究,薛泉宏等[9]的研究表明在林分生长初期,养分累积速率随林龄增加迅速加快,在林分生长中后期养分积累速率减慢。叶功富等[10]对不同林龄短枝木麻黄(Casuarinaequisetifolia)树体养分再吸收动态研究得出,短枝木麻黄成熟小枝N含量随着林分生长发育而显著升高,而P含量则呈降低趋势,叶片N含量随着林龄增加也增加。樊月等[11]的研究也得出,千年桐叶片C、N、P含量均表现为9年生的最高,杉木叶片N、P含量均表现为9年生的最高。可以看出,不同林龄阶段林木各器官养分变化差异较大,这也导致了不同时期林木各器官对养分的需求量有明显不同。近年来也有不少学者提出,营林模式对林木器官养分动态变化也有着显著的影响[12-14],因此,掌握混交林养分含量与分配特性,能进一步了解森林土壤的矿质营养供应状况,做到不同时期不同器官养分的精准动态监测,为研究人工混交林生态系统的养分循环奠定科学基础,对指导林业生产具有重要意义。
福建柏[Fokieniahodginsii(Dunn) Henry et Thomas]别名建柏、滇柏、杜柴等,为常绿高大乔木,属柏科福建柏属,是中国二级重点保护野生特有的单属种植物,也是福建省珍稀乡土树种,常常作为多种人工林的混交树种[15-17],主要分布在越南及中国西南部、南部至东部的海拔350~700 m的林地,在中国以福建中部最多[18]。近些年来已在福建柏材用林优良种质选育、栽培技术、人工林经营及遗传多样性等多个方面开展了不少研究并取得了一定的进展[19-24]。关于福建柏林分养分含量方面的研究主要集中在营林措施和环境因子对福建柏各器官元素含量的影响[25-26],而关于福建柏在不同年龄段各器官养分含量动态变化及与生长指标相关性的研究还鲜有报道,这使得监测养分动态变化规律和营林模式在不同时期对器官养分含量影响变得较为困难,对中国南方营造福建柏混交林后的长期林分养分管理、生态效益、材用经济效益的评价带来了一定的阻碍。探究福建柏纯林和混交林林分生长及养分随林龄变异特征,能更加直观了解不同林分条件下福建柏各阶段各器官元素含量变化,可以在福建柏各生长阶段实施不同的营林方案。
因此,本试验从福建柏不同生长发育阶段各器官养分分配层面出发,以两种人工林经营模式下的福建柏林分为研究对象,探究福建柏纯林和混交林在不同林龄阶段各器官养分含量的差异及与林分生长的相关性,解释福建柏人工林养分补偿机制和策略,旨在为福建柏混交林和纯林的营造和科学经营提供参考依据。
1 试验地概况
试验地位于福建省安溪丰田国有林场(25°16′~20′ N,118°1′~57′ E),地处典型的亚热带季风气候区,海拔500~650 m。气候条件温和,雨量充沛,全年平均气温19.5 ℃,最高37 ℃,最低0 ℃;年均降水量1 800 mm,夏季降水较多,无霜期330 d。土壤为黄红壤,花岗岩成土母质,土层较厚,可达60 cm,土壤理化性能良好。林下植被以芒萁(Dicranopterisdichotoma)为主,淡竹叶(Lophatherumgracile)、沿阶草(Ophiopogonbodinieri)、杜茎山(Maesajaponica)、五节芒(Miscanthusfloridulus)、乌毛蕨(Blechnumorientale)、菝葜(Smilaxchina)等少有分布。该地区是福建柏的适生区、良材及良种主产区和理想的科技研究试验区。
2 试验材料与方法
2.1 试验材料
试验于2019年8月开始,选择15年生、21年生和33年生的福建柏为研究对象,分别代表中龄林、近熟林和成熟林的福建柏的3个生长发育阶段[27],采用纯林和混交林两种营林模式,其中混交林为福建柏-马尾松混交林,混交方式均为株间混交,混交比例约为7∶3,共6种林分措施,福建柏纯林与混交林样地基本情况见表1。每种措施设置3个20 m×20 m标准地,共18个标准地。标准地设置后,于2019年9月进行每木检尺,包括福建柏及马尾松树高、胸径测量,计算平均树高、平均胸径后选定标准木。每块标准地选择一株标准木进行伐倒,共18株标准木进行树干解析。
福建柏和马尾松林造林前采用炼山整地,造林当年6月进行除草培土,9月进行施肥。造林第2年5月份及10月份各劈草一次。造林后第十年进行第一次间伐,并根据土壤、植被情况进行带状深翻,福建柏和马尾松林每年进行一次施肥和除草抚育。
2.2 试验方法
2.2.1 标准木的选择、单株材积及蓄积量计算在设置的福建柏纯林和福建柏马尾松混交林标准地上进行每木调查,主要使用围径尺测定单株胸径,用测高器测定树高。根据每木检尺统计结果,算出平均胸径和平均树高,选出每块标准地上胸径和树高最接近平均值的一棵福建柏作为标准木;确定好树干东西南北四个方向后进行伐倒,先用米尺测量出树干总长度,并在0 m、0.5 m、1.3 m、1.5 m处进行分段;而后以1 m为一个区间进行截断,将最后一个不足1 m的区间段树干作为梢头,分别在每个区间的底端截取厚度4 cm左右的圆盘,在圆盘底面标记好东西线、南北线及区间号后用塑料薄膜密封带回室内;利用角磨机将各个圆盘正面打磨平整,采用LINTAB树木年轮分析仪观测树木圆盘的年轮,分别沿着东西线和南北线以2a为一个龄阶标定各龄阶位置,同时测量各龄阶四个方向上的直径,并计算其平均值。利用Casiofx-4800p解析木计算程序计算各棵标准木的单株材积的总生长量,蓄积量=标准木立木材积单位面积林地上保留株数。
2.2.2 福建柏各器官样品采集及养分含量测定对每块标准地选定标准木的根(细根为主)、枝(细枝)、叶(成熟功能叶)等器官进行东西南北方向和树干或树冠上中下混合采样,混匀后分为3个重复,每个重复鲜重约为300 g;将采集好的样品带回实验室清洗,置于105 ℃烘箱杀青15 min,然后80 ℃烘干至恒重,烘干后采用粉碎机磨碎,过2 mm筛,保存于自封袋中,用于C、N、P、K含量的测定。

表1 福建柏纯林与混交林样地基本情况
福建柏各器官全碳含量的测定采用重铬酸钾-烘箱加热法[28]测定;全氮的测定采用凯氏半微量定氮法,用全自动凯氏定氮仪(SKD-2000)测定;全磷和全钾测定参照林业行业标准LY/T 1270~1999的浓硫酸-高氯酸消煮法,全磷采用钼锑抗比色法测定,全钾采用火焰光度法测定。
2.3 数据处理与分析
采用Microsoft Excel 2010处理数据与图表绘制,采用SPSS 26进行单因素方差分析(One-way ANOVA)和LSD多重比较,分析各林分生长指标和各器官养分含量在不同林龄、不同营林模式间的差异。利用Pearson相关分析法对福建柏各器官养分含量与林分生长特性进行相关性分析。
3 结果与分析
3.1 营林模式对不同林龄福建柏林分生长的影响
由表2可知,不同时期纯林模式下的福建柏生长指标均低于福建柏混交林,且随着生长年限的增加,差异越显著。15年生混交林福建柏胸径、树高与纯林无显著差异;21年生和33年生混交林福建柏胸径、树高则显著大于纯林(P<0.05)。在33年生时,其胸径和树高差距达到最大,混交林福建柏胸径比纯林高出13.1%,树高则比纯林增加了6%,且21年生到33年生期间福建柏的胸径生长远远快于15年生到21年生时间段,表明营造混交林可以促使福建柏林分在一定生长发育阶段,有效提高福建柏及其混交树种的胸径和树高生长量,这也使得福建柏混交林单株材积增长速度高于纯林,在33年时两种林分福建柏单株材积差值达到最大,混交林显著高出了44.5%。
不同林龄福建柏纯林总蓄积量差异不显著(P>0.05),而福建柏混交林总蓄积量则存在极显著差异(P<0.01)。当林分年龄达到33年生时,混交林总蓄积量达到381.9234 m3/hm2,比15年生和21年生福建柏混交林分别高出了67%和64%;对比两种林分的总蓄积量,混交林在生长前中期总蓄积量均高于纯林但差异不显著,而生长后期的混交林总蓄积量较纯林显著提高了65%。可以看出混交效益对福建柏林分总蓄积量具有促进作用,且在林分生长后期效果更明显。
3.2 营林模式对不同林龄福建柏各器官元素含量动态变化的影响
3.2.1 福建柏叶片各元素含量动态变化由图1可以看出,两种营林模式下的福建柏叶片C、N元素含量均随林分的生长呈现出增加的趋势。15年生的福建柏混交林C、N含量分别比纯林高出9.41%和45.27%,差异达到显著性(P<0.05)。此阶段为林分快速生长期,植株能积累较多的有机物,混交林的营造可以很大程度提高福建柏叶C、N元素的含量,积累更多养分。而随着林分年龄的增加,福建柏混交林叶片C、N含量虽然仍高于纯林,但差异较小,未达到显著性水平(P>0.05)。

表2 不同林龄和营林模式的福建柏林分生长指标对比
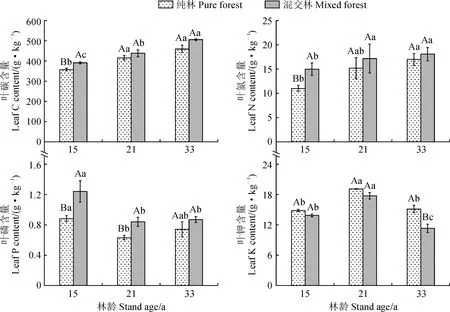
不同大写字母表示同一林龄的纯林与混交林间差异显著(P<0.05);不同小写字母表示纯林或混交林的不同林龄间差异显著(P<0.05),下同图1 福建柏叶片元素含量动态变化Different capital letters indicate significant difference between mixed forest and pure forest in the same stand age (P<0.05); Different lowercase letters indicate significant difference between different ages of pure forest or mixed forest (P<0.05), the same as belowFig.1 Dynamic changes of element contents in leaves of F. hodginsii
福建柏混交林和纯林叶片P元素含量在生长前期均达到最高,分别为1.24和0.88 g·kg-1,随着林分生长发育进程的推进均呈现出先减少后增加的趋势,15和21年间混交林P含量显著高于同龄纯林(P<0.05),而33年生时福建柏混交林与纯林的差异则未达到显著水平(P>0.05)。叶片K元素含量则与P元素表现出相反的趋势,即随着林分年龄的增加先升高后降低,在21年生时达到峰值,而福建柏混交林叶片的K含量较纯林低了7.2%,生长前中期差异不显著,但在33年生时达到显著差异;两种营林模式下的福建柏叶K含量在不同林龄间均存在极显著差异。
3.2.2 福建柏枝条各元素含量动态变化图2表明,枝条C含量随着林分生长呈现先下降后上升的趋势,而混交效果对3个时期福建柏枝条C含量均有一定的促进作用,但增幅效果较小,差异不显著(P>0.05)。枝条N元素含量则随林龄增加而减少,生长后期枝条N元素的含量低于生长前中期;混交林模式下的福建柏林分N元素含量在15年生和33年生时分别比纯林高出32.4%和47.1%,均具有显著性差异(P<0.05)。

图2 福建柏枝条元素含量动态变化Fig.2 Dynamic changes of element contents in branch of F. hodginsii
此外,混交效益还影响了P素在福建柏林分枝条不同时期的含量动态变化趋势。福建柏混交林林分在生长中前期枝条P素含量较纯林低了10.9%,达到显著差异性(P<0.05),但其P素含量会随着林分生长发育提高福建柏枝条中的分配,这与纯林P素动态变化趋势相反,也导致了混交林福建柏枝条P素含量在33年生时显著高于纯林(P<0.05)。两种林分模式下福建柏枝条中K元素的含量都是随着林分生长而减少,且混交林枝条的K元素含量3个时期均低于纯林,并在生长后期具有显著差异。
3.2.3 福建柏树干各元素含量动态变化图3显示,两种模式下的福建柏林分树干C含量均会随着林龄增加而上升,混交林树干C含量在21年生时显著高于纯林(P<0.05),但随着林分生长发育,两种林分树干C含量差异会逐渐减小,这表明混交效果对树干C含量的影响主要集中在生长前中期。而树干N含量在林分生长阶段会呈现先下降后上升的趋势,两种福建柏林分的福建柏树干全氮虽然随发育阶段推进的变化趋势基本一致,但变化幅度存在较大的差异。与纯林相比,15年生混交林全氮含量无显著差异(P>0.05),但21年生和33年生混交林均存在显著差异(P<0.05),分别比对应林龄的纯林增加了32.3%和137.7%。说明营造混交林后对树干N元素的影响主要在林分发育后期,可大幅度提高N素在近成熟林和成熟林福建柏树干中的分配。

图3 福建柏树干元素含量动态变化Fig.3 Dynamic changes of element contents in trunk of F. hodginsii
两种林分树干P元素含量动态变化均是随着林分年龄增加而减少,混交林树干P元素含量在3个生长阶段都低于纯林但差异不显著。而福建柏纯林的福建柏树干K含量随着林分生长一直降低,混交林则呈现完全相反的趋势:在15年生时,纯林树干K含量比混交林高了19%,差异未达到显著性(P>0.05);随着林分生长,纯林模式下的福建柏树干K含量会逐渐减少,而混交效益则会促进树干K含量的增长,在33年生时达到最大,显著高于纯林(P<0.05)。
3.2.4 福建柏根部各元素含量动态变化由图4可知,两种林分的福建柏根C含量随着林龄增加而减少,混交效益会降低福建柏C含量使其低于纯林,在33年生时达到显著性差异(P<0.05),混交林的C含量较纯林低了近10%;N元素的动态变化趋势与C元素相近,而混交效益则会在不同生长阶段显著提高福建柏根N元素的含量(P<0.05),尤其在林分生长初期,混交林根的N元素含量较纯林高出了114%。这表明随着福建柏林分生长发育,根的主要营养元素分配会降低,而混交效益则会改变福建柏林分不同时期C、N分配比例,其C/N也会呈现下降的趋势。

图4 福建柏根元素含量动态变化Fig.4 Dynamic changes of element contents in root of F. hodginsii
两种林分福建柏根的P元素含量动态变化趋势基本一致,在生长中期时P元素含量会减少,随着林分生长发育进程的推进,P元素含量在33年生时达到另一个峰值。可以看出,混交林P元素在3个生长期均高于纯林,并随着林分年龄的增长,差异越显著。纯林K元素在21年生达到峰值,生长后期则显著降低,而混交效益则会在生长前期和后期促进根K元素的含量,使其较高于纯林。
3.3 福建柏各器官养分与林分生长特性的相关性分析
由表3可以看出,福建柏叶和树干的C含量与胸径、树高、材积和林分总蓄积均呈现显著正相关,而根的C含量则与生长指标呈显著负相关,这说明福建柏林分叶和树干C含量的增加能促进胸径、树高、材积和总蓄积的生长,同时降低根C元素的比例;而树干和根N元素含量的增加可以显著提升福建柏总蓄积。

表3 福建柏各器官养分与林分生长特性的相关性分析
P元素含量对福建柏生长指标的影响主要集中在树干和根。树干P含量与福建柏胸径、树高、材积和总蓄积呈极显著负相关;而根P含量则与胸径、材积和总蓄积呈显著正相关。叶、枝条、根的K元素含量与胸径、树高、材积和总蓄积具有显著负相关性,这也表明这些福建柏器官过高的P、K元素含量可能会导致福建柏生长受限。
4 结论与讨论
本研究中,不同时期纯林模式下的福建柏生长指标均低于福建柏混交林,且这种差异随着林龄的增加而更明显,表明了营造混交林可以在一定生长发育阶段有效提高福建柏及其混交树种的胸径和树高生长量,使得福建柏混交林单株材积增长速度高于纯林,在33年生时生长指标差异均达到显著性水平。罗毓明等[29]的研究表明,林分主要树种种类越丰富,林分结构越稳定,具有更佳的保育和维持林下生物多样性功能,其林木生长效果也更优于树种种类较少的林分。张瑞秀[30]的研究结果也验证了混交造林能显著提高福建柏单株生长量,提升林分单位蓄积量,这与本试验的研究结果相一致。此外,不同林龄福建柏纯林总蓄积量差异不显著,而福建柏混交林总蓄积量在生长中后期存在显著差异,这可能是由于纯林中种内竞争压力大,有限的环境资源条件下,密度效应大于林分生长效应,大量个体被淘汰掉,导致林分总蓄积量没有明显增加;而福建柏混交林在林分生长发育前期并没有明显表现出混交效果和优势,种间竞争比较激烈,林分总蓄积在此阶段并没有表现出显著增加,到了生长中后期,各树种充分利用环境中不同高度层次的光照、温度和水资源,种间竞争减小,各树种个体生长迅速,林分蓄积量快速增加,带来较大的经济收益,这也与刘卓[31]的研究结果相似。总体而言,混交林林分树种多样性更有利于林分的生长发育和生物量积累,使其在不同时期生长指标均优于纯林。此外,有研究表明[32-33],混交林的营林方式相比于纯林很大程度提高了林木的生态功能,其中合理的树种组成、各树种之间组成系数分配较均匀的林分结构相对较稳定。
森林生态系统的碳固存格局与森林的生长发育过程密切相关,净碳固存取决于林龄差异[34-35]。随着林龄的增长,森林植被碳库通常呈增加趋势,但人工林树体各器官全碳分配格局随林龄的变化存在较大的差异[36-38]。在本研究中,两种林分的福建柏叶和树干C含量随着林龄的增加而提高,根C含量呈现出相反的趋势,造成这种养分分配比例的原因可能是因为福建柏在生长中后期会将养分和水分运输到生理活动旺盛的部位,通过降低根C含量来减少木质化的程度,从而增加吸收水分效率。而枝条的C含量则在21年生时最低,这可能是由于在近熟林阶段,福建柏老枝自然枯死和整枝的作用,新的功能枝处于生长期,积累的全碳少,导致该林龄阶段枝条碳含量较低。朱育锋等[39]研究也表明,近熟林和成熟林的枝条C含量低于中龄林和过熟林。此外,营林模式也是影响福建柏各器官C元素分配的关键因素,混交效益在3个时期均能促进福建柏叶片和树干C元素的含量,叶片是进行光合作用、合成有机物的重要场所,对C素的需求量较高[40],因此混交效益在一定程度上也能提高福建柏林分光合作用强度。而福建柏混交林3个时期根的C元素含量均低于纯林,且在生长后期具有差异显著性,说明混交效益可以通过降低根的C元素含量来减少木质化的产生,提高水分的运输水平,更有利于福建柏后期生长发育的需求,这也与前人的研究结果相似[39]。
随着林龄的增长,两种经营模式的福建柏叶N含量呈现出上升的趋势,枝条和根的N含量均有所下降,树干N含量则呈现先下降后上升的趋势,不同器官N元素的变化趋势和含量差异较大,这可能与N元素本身的特性有关,N在林木生长过程中容易移动,不同林龄阶段各器官的生长以及生理活动需要消耗的N素量存在明显的差异[27]。由于N素是构成叶片光合结构的重要元素,代谢活动较旺盛,因此叶片N素养分含量最高,而干材以木质为主,生理功能较弱,所含的大部分养分被利用或转移,故而N素养分含量较低[41]。混交模式下的福建柏在3个阶段各器官N含量均高于纯林,这也表明了混交效益能促进福建柏各器官N元素的积累,其中对根的N元素含量影响最显著,这可能是营造混交林可有效改善土壤理化性质,提高土壤肥力,进而促进福建柏根系对N素的吸收、利用和转移[42-43]。而不同时期混交增益效果也有一定差异,在生长前中期,福建柏混交林叶片的N元素显著高于纯林,随着林分年龄的增加虽然仍高于纯林,但差异不显著,这也说明了福建柏林分生长后期叶片有机物积累已近饱和,营林模式对其影响效果较小。
福建柏各器官P含量随林龄的变化趋势差异较大,叶和根在生长前期能积累较多的P元素,以后逐渐呈现下降趋势,到生长后期其P元素含量又有一定的上升,而树干P含量则随林龄增加而减少。这可能由于混交林在发育中后期阶段,福建柏的叶和根系生理活动较强,需要更多的营养元素,而树干又以木质为主,生理功能较弱,将P素养分更多地转移到生理活动强的器官利用[44-46]。两种模式福建柏林分枝条的P素含量变化趋势差异较大,15年生纯林枝条P含量显著高于混交林,但会随着林龄增加而减少,而混交效益则改变了福建柏枝条P元素的含量和变化趋势,使其在生长后期阶段显著高于纯林。此外,营造混交林能促进相同时期福建柏叶和根P元素的积累,降低树干P元素的比例,这也优化了混交林不同时期器官P素养分的比例构造,促进林木在中龄阶段的生长,满足生长旺盛部位养分的需求。
不同时期福建柏各器官K元素含量变化趋势有显著差异,这可能与K元素移动性较强的特点有关[47],两种林分模式下福建柏枝条中K元素的含量都是随着林分生长而减少,且混交林的K元素低于纯林,在生长后期达到显著差异。这也说明了营造福建柏马尾松混交林可以在一定林分发育阶段降低K元素在枝条中的分配量,将更多的K元素分配到其他器官,最大程度维持树体生长生理活动正常进行。随着林分生长,纯林模式下的福建柏树干K含量会逐渐减少,而混交效益则会促进树干K含量的增长,并在33年生时达到最大,显著高于纯林。混交效益对树干P含量的影响不显著,却能改变K元素在树干中动态变化比例。
通过相关性分析可知,福建柏林分叶和树干C含量的增加对福建柏胸径、树高、材积和总蓄积的生长有显著促进作用,而树干和根N元素含量的增加可以显著提升福建柏总蓄积。树干P含量与福建柏胸径、树高、材积和总蓄积呈极显著负相关。根P含量水平与胸径、材积和总蓄积具有显著正相关性。叶、枝条、根的K元素含量则与胸径、树高、材积和总蓄积具有显著负相关性,这也表明这些福建柏器官过高的P、K元素含量可能会导致福建柏生长受限。
综上所述,不同时期福建柏各器官养分变化差异较大,可根据各器官养分动态变化规律进行精准高效施肥;混交林模式下福建柏林分各项生长指标优于纯林,从元素分配比例上也更优化,能更好地将养分分配到林木生长各阶段需求量较高的器官中。
