凤阳山常绿阔叶林优势种木荷的种群结构与动态
2022-05-24周红敏王宏翔张弓乔
彭 辉,周红敏,王宏翔,张弓乔
(1. 凤阳山管理处,浙江 丽水 323000;2. 龙泉市林业科学研究院,浙江 龙泉 323700;3. 广西大学林学学院,广西 南宁 530004;4. 中国林业科学研究院林业研究所,北京 100091)
种群结构是植物种群的最基本特征,种群的结构与动态是种群生态学研究的核心之一,是揭示阐明种群更新与维持机制的关键[1]。分析种群现实特征并对其未来发展进行预测,能为该物种及其群落的科学保护及可持续经营提供理论依据。生命表、存活曲线及生存分析等是研究种群结构和动态的重要方法[2],其中,编制静态生命表可以展现种群的动态特征,利用生存分析函数可以展示种群生存规律[3],结合时间序列分析等可以进一步探讨种群生态学过程,预测其发展的趋势[4]。
木荷(Schima superbaGardn. et Champ.)是山茶科(Theaceae)木荷属(SchimaReinw.)的常绿乔木[5]。木荷习性偏阳,常与杉木(Cuninghamia lanceolata(lamb.) Hook.)等混交[6],是我国亚热带山地最重要的乡土阔叶造林树种和重要的工业用材树种之一。木荷是中亚热带常绿阔叶林的优势树种,也是常绿阔叶林恢复过程中的先锋树种[7]。在常绿阔叶林森林群落稳定和演替过程中扮演重要角色,但目前关于木荷天然种群在常绿阔叶林演替过程中的生存现状和数量动态研究较少。鉴于此,本研究以凤阳山常绿阔叶林木荷种群为研究对象,通过建立静态生命表,绘制存活曲线,并结合种群数量动态量化研究及时间序列分析等方法深度剖析木荷种群的更新特征及生态对策,以期对木荷天然种群进行科学保护,进而为凤阳山常绿阔叶林植被恢复与重建提供科学依据。
1 研究区概况
研究区位于武夷山系洞宫山山脉中段的浙江省龙泉市南的凤阳山-百山祖国家级自然保护区内(119°06′~119°15′ E、27°46′~27°58′ N) ,面积151 km2。土壤以黄壤土为主,地带性植被为中亚热带常绿阔叶林,植被垂直分布明显。区内年平均气温12.3 ℃,年日照1 515.5 h,平均相对湿度80%,年蒸发量1 171.0 mm,年降水量2 438.2 mm。保护区内有种子植物164 科、659 属、1 488 种,是南方红豆杉(Taxus wallichianavar. mairei(Lemée H. Léveillé.) L. K. Fu et Nan Li)、 伯 乐 树(Bretschneidera sinensisHemsl.)等6 种国家一级重点保护植物及多种珍稀濒危植物重要分布区[8]。
在全面考察的基础上,依照美国CTFS (the Center for Tropical Forest Science) 的森林生物多样性监测规范,在凤阳山典型常绿阔叶林内设立80 m × 100 m 的固定样地(海拔1 300 m,坡度20°)。样地内划分为5 m × 5 m 的小样方进行每木检尺,调查胸径(DBH)≥1 cm 的所有乔木树种,挂牌并记录每棵树的树种、位置坐标(x,y)、胸径、树高、冠幅等因子;灌草层记录各物种的高度、盖度、多度等指标。
2 研究方法
2.1 划分木荷种群龄级
在相同的环境条件下,同一树种的径级和龄级对环境的反应规律具有一致性[9]。因此,在木荷种群年龄难以准确获取的情况下,参照亚热带常绿阔叶林相关研究,结合凤阳山0.8 hm2样地内实际情况[10],以径级结构代替年龄结构来分析木荷种群的数量动态变化。按照DBH 的大小等级,把径级I(DBH < 5 cm)作为幼树,之后按DBH 大小以5 cm为一级划分(上限排外),把径级II (5 cm≤DBH <10 cm)作为小树、径级III -径级Ⅴ(10 cm≤DBH < 25 cm)作为中树、径级Ⅵ-径级Ⅸ (25 cm≤DBH < 45 cm)作为大树,共分为4 个生长阶段[11]。
2.2 编制静态生命表
因现实中无法对某一种群的生活史进行跟踪分析,特定时间生命表只能根据同一时间的种群个体径级进行编制,进而反映世代重叠的年龄动态历程[12]。在依据自然种群的不同龄级进行分析时,多因采取空间代替时间、径级代替龄级等会导致死亡率出现负值的情形。大多数研究会对数据进行均滑处理,但因此反而忽略了种群真实的生态过程。为此,本研究采用各龄级实际个体数编制静态生命表,并绘制种群结构图。计算公式如下:
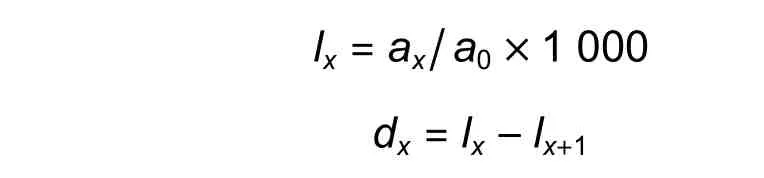
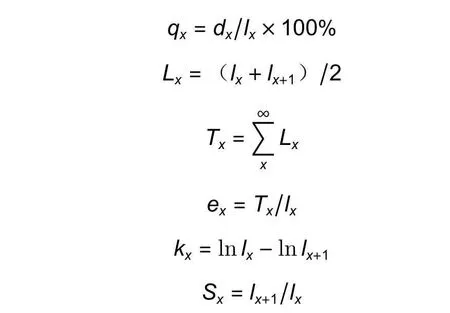
式中:ax是x龄级内的个体数,a0是ax的初始值,lx是在x龄级标准化后的存活个体,一般为1 000,dx和qx是从x龄级到x+ 1 龄级间标准化的死亡数和死亡率,Lx是x到x+ 1 龄级的存活个体,Tx是从x龄级起的个体总数,ex是x龄级的期望寿命值,kx是损失度(消失率),Sx是存活率。
2.3 拟合存活曲线
种群各龄级个体的存活状况及过程可以通过存活曲线直观反映[13],一般以龄级为横坐标,以实际存活个体数ax和存活个体对数值l nax为纵坐标,绘制种群存活曲线图,采用Hett 等[14]提出的数学模型,依据Deevey 对存活曲线的分类,若指数方程Nx=N0e−bx拟合效果好,存活曲线符合DeeveyⅡ型描述,种群中各龄级个体的死亡率相同,为对角线型。若符合幂函数Nx=N0x−b方程,则存活曲线为DeeveyⅢ型[15],种群中的幼龄个体具有较高的死亡率,随着龄级增加死亡率降低且稳定。
2.4 量化种群动态
根据陈晓德[16]的方法,以种群两个相邻龄级个体的数量变化指数(Vn)、种群年龄结构数量动态变化指数(Vpi)及考虑外部干扰时种群年龄结构动态指数(V′pi)等客观准确的量化分析木荷种群的动态变化。公式为:

式中:Vn是种群的n龄级到n+ 1 龄级个体数量变化动态,Vpi代表整个种群的数量动态变化指数,Sn和Sn+1分别是相邻龄级的各组的个体数,k表示种群的龄级数量,max/min()表示取其极大/极小值,Vn、Vpi取值在[−1,1]之间,取负值、零、正值时分别表示种群相邻两个龄级间(整个种群)是衰退、稳定、增长的动态变化。当有外部干扰时,用i表示整个种群年龄结构动态,其还与龄级数量(k)及各龄级个体数(Sn)相关,则:也是衡量随机干扰对种群结构动态影响的指标,
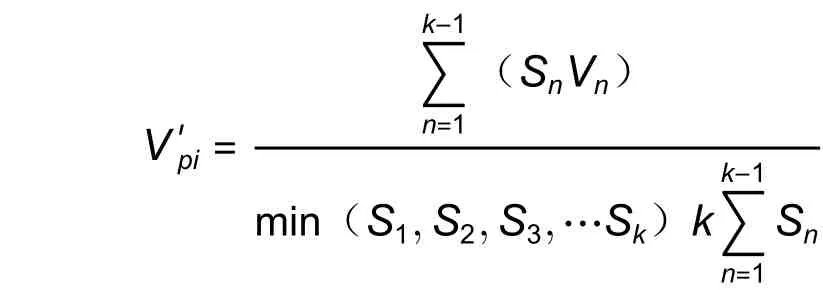

2.5 时间序列模型
本研究中运用时间序列分析中的一次平均推移法,对木荷种群的年龄结构在未来2、4、6、8 个龄级发展趋向进行预测[17],模型为:
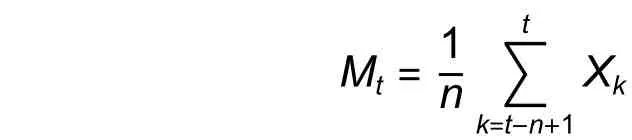
式中:n为需预测的未来时间年限,t为龄级,为未来n年时t龄级的种群存活数量,是近期的n个观测值在t时刻的平均值,称为第n周期的移动平均,Xk为当前k龄级种群的存活数。
3 结果与分析
3.1 木荷种群年龄结构
本研究样地中共有木荷692 株,种群密度为0.086 5 株·m−2,幼树和小树的比例较高(图1),占整个种群的84.97%。小树阶段(径级II)个体量急剧下降,占整体的20.8%,表明样地中木荷种群存在激烈的种内和种间竞争,幼龄个体被大量淘汰,只有少数个体能进入下一生长阶段。总体看,木荷年龄结构呈典型“倒J 形”种群,属于增长型种群。
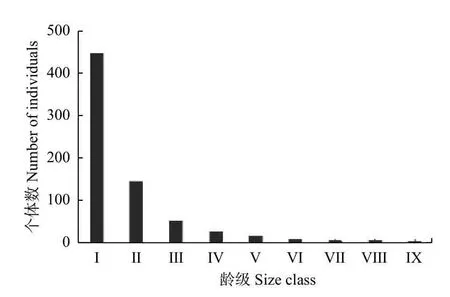
图1 凤阳山木荷种群的龄级结构Fig. 1 Size class structure ofSchima superbapopulation
3.2 种群生命表
对木荷种群静态生命表的分析(表1)可知:木荷种群不同龄级个体存活数差别较大,特别是Ⅰ-Ⅱ龄级,存活个体下降迅速,在环境及竞争因素作用下淘汰了近68%的个体。期望寿命()反映了龄级内种群个体的平均生存能力,木荷种群在Ⅲ-Ⅳ龄级期望寿命较高,死亡率较低,在Ⅶ龄级后个体生命期望值较低。整体上,种群期望寿命虽呈现出小幅波动,但期望值总体偏低,呈下降趋势。木荷种群的死亡率()二者有相同的变化趋势,均在Ⅵ径级出现高峰,死亡率为71.4% 消失率为1.253,而非呈单调递减趋势。
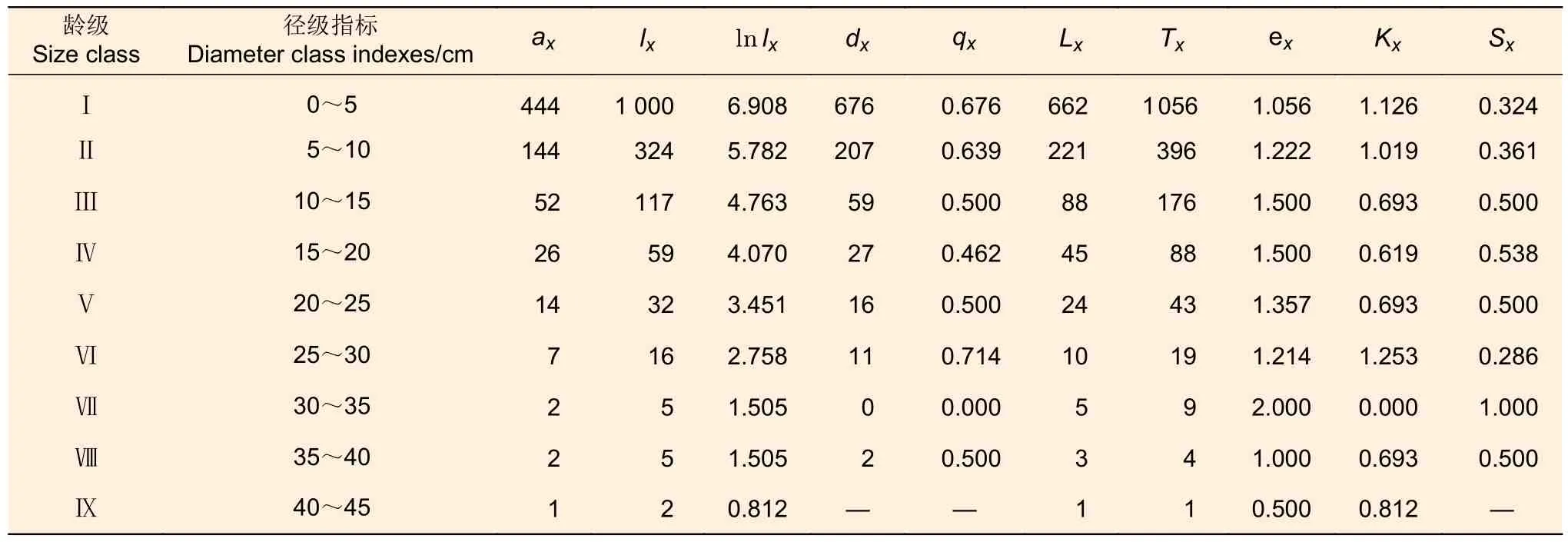
表1 木荷种群静态生命表Table 1 Static life table ofSchima superbapopulation
3.3 种群存活曲线
存活曲线有3 种类型,Ⅰ 型是凸曲线,该类型早期死亡率低,但达到平均生理年龄时,会在短期内个体全部死亡,Ⅱ型、Ⅲ型分别为直线型和凹线型,Ⅱ型特征为种群各年龄段死亡率基本相同,Ⅲ型是前期种群个体死亡率高,一旦进入某一龄级死亡率低而稳定。因此,分别以实际存活个体数ax和其对数值 lnax做存活曲线图(图2)。由图2可知:ax做的存活曲线图在凹型和直线型之间,lnax做图近似于直线型,因此,采用指数方程Nx=N0e−bx和幂函数Nx=N0x−b方程,进行拟合效果检验,结果见表2。对比4 个拟合方程可知:以ax和l nax为因变量拟合的指数模型中F值和R2均明显高于相应的幂函数模型,因此,认为木荷种群的存活曲线更符合Deevey Ⅱ型描述,这表明木荷种群各龄级具有近乎相同的死亡率,与在静态生命表中的分析一致。
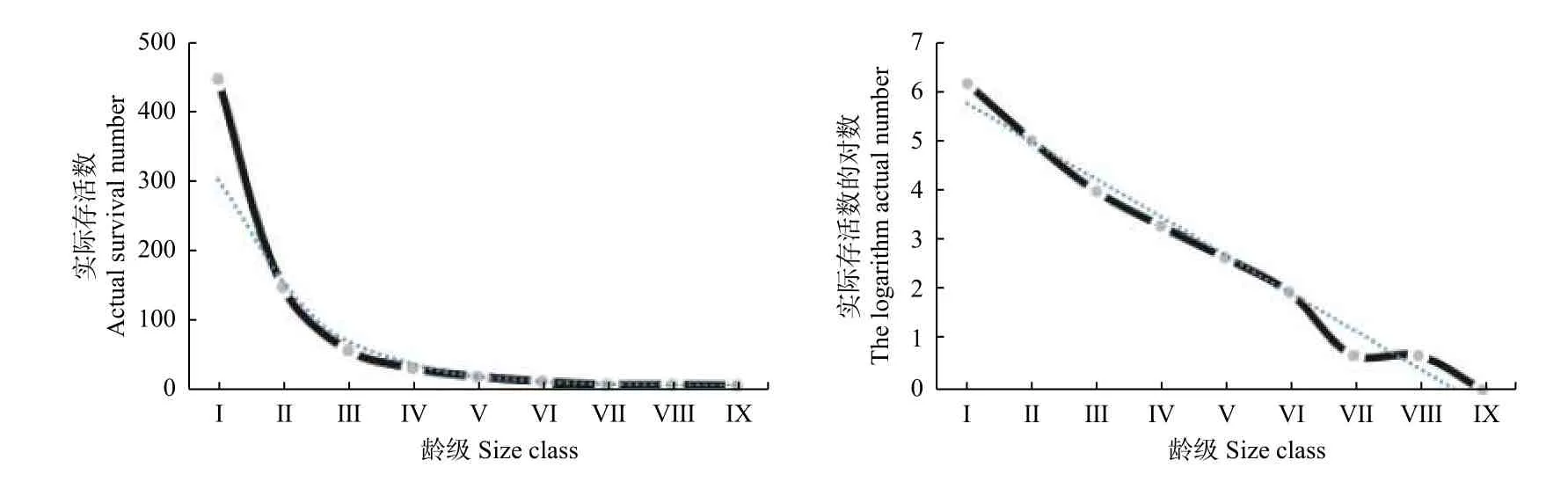
图2 凤阳山木荷种群的存活曲线Fig. 2 survival curve ofSchima superbapopulation

表2 木荷种群存活曲线方程Table 2 The equation of survival curve forSchima superbapopulation
3.4 种群动态与时间序列
木荷种群相邻龄级数量变化动态指数随着龄级的增大表现不同(表3),种群动态指数大多为正值,说明种群多数时期属于增长的结构动态,只是在V7时处于稳定阶段,均大于0,说明即使受到外界随机干扰时,种群仍能处于增长状态,但对随机干扰有较高的敏感度。

表3 木荷种群存动态变化指数Table 3 Dynamic index ofSchima superbapopulation
以各龄级的实际存活数作为基数,以时间序列的一次平均推移法对木荷种群未来2、4、6、8 个龄级后的存活个体数量进行预测,从表4 可知:在未来4 个龄级后,随着时间推移各龄级个体数量均逐渐增加,且增加的趋势增大,其中,在Ⅵ龄级,个体数量经过2、4、6 个龄级后分别增加了57.1%、257.1%、1 542.9%,而Ⅷ龄级经过4、6、8 个龄级后个体增加了2、7.5、42 倍。说明今后一段时期的演替过程中,木荷种群仍会在常绿阔叶林中占据优势地位。

表4 木荷种群存动态变化指数Table 4 Time sequence ofSchima superbapopulation
4 讨论
优势物种是塑造群落结构的骨架[18],对群落的外貌和类型具有重要调控作用。种群结构动态变化规律,反映了其对环境因子的适应[19]。木荷喜光,种子有种翅,在样地中广泛分布。在凤阳山常绿阔叶林当前的演替阶段中,木荷种群为典型的倒“J”形,属于增长型种群,其中,龄级Ⅰ和龄级Ⅱ个体占种群数量的比例较大,说明木荷拥有大量的幼苗储备,这与李立等[11]的研究结果一致。小树阶段(龄级Ⅱ)竞争最激烈,个体出现死亡高峰,该阶段植物对其对环境资源,特别是光照、水分、土壤养分及生长空间的需求激增,是更新和演替过程中最敏感的阶段[20],受Jazen-Connell 效应制约,同种个体的竞争激烈,自疏明显,结果为其他物种入侵和定居提供空间,从而提高、维持群落的多样性。
静态生命表及死亡率和消失率的分析结果显示,种群的期望寿命随着龄级增大呈下降趋势但略有波动,死亡率和消失率有着相同的变化趋势,在Ⅵ径级出现高峰,可能是因为木荷种群已经开始进入生理死亡年龄阶段。存活曲线更符合Deevey-Ⅱ型描述,说明外界的干扰可能一直存在,导致种群各年龄段具有相似的死亡率。相邻龄级的动态指数均大于零(V7=0),为正值,说明木荷种群虽有轻微波动但一直处在增长动态变化中,种群动态指数和受外界干扰时的动态指数(Vpi和)均为正值,说明木荷种群为增长型种群,但对外界干扰敏感指数较高,暗示种群仍面临较大的外界干扰。
时间序列是分析预测种群未来年龄结构的重要手段[21],研究发现,在经历2、4、6、8 个龄级后,木荷种群个体数量均呈现上升趋势,特别是幼龄个体充足是,增长的潜力更大,说明充足的幼苗储备是各龄级个体增加的前提保障,幼苗多则种群增长潜力大。
5 结论
木荷作为中亚热带常绿阔叶林的优势种,对维持群落稳定具有重要意义,与清凉峰保护区木荷稳定型种群相比[22],凤阳山保护区的木荷种群与古田山木荷种群更相似,均为增长型种群,更新良好。凤阳山木荷种群的年龄结构合理,但是对外界环境变化敏感。一旦环境遭到破坏,必将严重影响到木荷的正常生长发育。因此,必须巩固加强现有的群落结构和生境保护,避免人为干扰造成的破坏。
