线粒体未折叠蛋白反应调控线虫衰老研究进展*
2022-05-23郑天琳张文渊刘剑峰
郑天琳 张文渊 春 雷 刘剑峰
(华中科技大学生命科学与技术学院,武汉 430074)

秀丽隐杆线虫(Caenorhabditis elegans,C.elegans),雌雄同体,雄性个体仅占群体的0.2%,是分子生物学、发育生物学、神经生物学与衰老生物学研究的经典模式生物。
随着时间的推移,生物体都将面临衰老的考验。衰老对个体而言是一个有害而复杂的过程,伴随着衰老的进行,细胞和机体动态平衡逐渐下降[1]。而衰老造成的细胞水平的机能障碍很大程度上是由蛋白质和细胞器损伤的逐渐积累导致。线粒体是生物机体内极其重要的细胞器,多数与衰老相关的疾病,特别是神经退行性疾病,都与线粒体功能紊乱有关。在衰老过程中,线粒体会因为多种因素导致自身功能下降。当线粒体的功能受损时,细胞会激活一种称为线粒体未折叠蛋白反应(mitochondrial unfolded protein response, UPRmt)的转录应答机制,从而维持线粒体蛋白质稳态[2],应对环境压力,保持机体健康。UPRmt作为一种保守的机制,在各生物中都有广泛的发现与研究。而线虫作为一种生命周期短、一般情况下可以自交繁殖、进化机制保守的模式生物,是研究UPRmt极佳的模式动物。本文主要讨论了线粒体的特征、线虫中UPRmt的信号转导机制及其对线虫寿命的影响。
1 线粒体
线粒体是膜结合的细胞器,具有外膜和内膜,将其分割成膜间空间(inner membrane space,IMS)与基质(matrix)两个部分。线粒体在生物体中有着重要而普遍的作用,它主要承担3个关键的功能,分别是提供能量、进行代谢和诱导凋亡。线粒体负责通过三羧酸循环(tricarboxylic acid cycle,TCA)协调还原态烟酰胺腺嘌呤二核苷酸NADH供体底物的合成,并负责产生来自氧化磷酸化(oxidative phosphorylation,OXPHOS)的能量,还参与了从钙稳态的调节到先天免疫反应和程序性细胞死亡的众多生物学过程[3]。
哺乳动物的线粒体含有1500 多种蛋白质,但只有13种OXPHOS的蛋白质(线虫中有12种)是由线粒体DNA(mtDNA)编码而成[4]。因此线粒体内的蛋白质组由线粒体基因编码蛋白和核基因编码蛋白共同构成。核基因编码的蛋白质首先在细胞质中翻译,再被靶向导入,并分选到线粒体的不同区室中,如进入线粒体基质内、定位在线粒体内膜或者外膜上、游离在IMS 内部等[5]。这些蛋白质在进入线粒体后会先进行高度折叠,接着与mtDNA 编码的蛋白质亚基按照固定比例组装成有功能的大分子蛋白质复合物。这种线粒体内的组装主要依靠线粒体分子伴侣[6-8]和蛋白酶[9]完成。线粒体热休克蛋白70 (mitochondrial heat shock protein 70,mtHsp70)是一个重要的分子伴侣蛋白,它与基质侧的内膜导入复合物有关,并积极介导新生蛋白的导入和折叠。另外,线粒体编码的热休克蛋白相关蛋白1(translocon-associated protein 1,TRAP1/mtHsp90)和少量核编码的大分子伴侣热休克蛋白60/热休克蛋白10(heat shock protein 60/heat shock protein 10,Hsp60/10)复合物,也促进了线粒体基质定位蛋白质的折叠[6-8]。线粒体核糖体[10]和呼吸链成分[11]则由这些线粒体内的分子伴侣进行组装。线粒体中的多种蛋白酶也是维持其内部蛋白质稳态的重要调节因子。基质中含有Lon 蛋 白 酶 同 源 物(Lon protease homolog,mitochondrial,LonP)和人酪蛋白水解线粒体基质肽酶蛋白水解亚基同源物 (caseinolytic mitochondrial matrix peptidase proteolytic subunit,ClpP)两种主要的蛋白酶复合物[9],用来降解含有特殊结构的肽链,从而促进降解线粒体中大多数受损的蛋白质。
2 线粒体的蛋白质稳态与UPRmt
蛋 白 质 稳 态 (proteostasis, protein homeostasis)是指在特定时间点细胞内蛋白质组中的特定蛋白质合成、折叠与去折叠、修饰与降解等过程达到的一种平衡状态[12]。由于线粒体的多种功能依赖于复杂的蛋白质网络,因此蛋白质稳态对于线粒体来说至关重要。
在复杂性层次不同的生物中,线粒体蛋白质稳态防御通路的缺陷会对机体的健康和衰老产生重大影响。许多编码调控线粒体蛋白质稳态机制的基因突变会导致蛋白质稳态崩溃的加剧,最终表现为衰老相关疾病。此外,急性环境损伤,如暴露在以线粒体为靶标的杀虫剂、除草剂和抗生素中,会产生大量的错误蛋白,导致线粒体蛋白质稳态平衡的过度负载,并且扰乱线粒体稳态。
蛋白质折叠稳态是细胞适应性的核心,生命体在对压力的抗争中进化出多种反应用以保持机体的稳态,其中很重要的一种反馈机制就是未折叠蛋白反应(unfolded protein response,UPR)。这种反应首先在内质网中被发现[13],当内质网腔中积累大量的未折叠蛋白或错误蛋白,这种信号被分子伴侣识别并传导到细胞核中,通过转录因子促进内质网相关分子伴侣的转录诱导,一方面帮助未折叠的蛋白质进行折叠,一方面抑制翻译从而降低折叠的负荷[14]。在线粒体中,也存在相似的蛋白质反馈机制,称之为UPRmt。细胞采用质量控制(quality control)机制来积极监测线粒体功能,在出现线粒体损伤时启动UPRmt进行保护。UPRmt能够将线粒体应激信号传递至细胞核,以启动对蛋白酶和分子伴侣等细胞核编码的线粒体保护基因的转录[15]。UPRmt需要激活广泛的基因表达来抵消线粒体应激,同时染色质修饰相关的酶会诱导染色质结构发生改变,从而影响相关基因的转录水平。此外,UPRmt的激活也会导致大量的代谢重组,如上调抗氧化酶的表达,激活与糖酵解相关的基因并抑制与氧化磷酸化相关的基因[16],从而重置细胞代谢状态。
引起UPRmt激活的因素十分广泛,主要包括阻断线粒体翻译[17]、核DNA(nDNA)与线粒体DNA 翻译 之间 的不 均衡[17]、线 粒 体DNA 的 缺失[18]、表达错误折叠的蛋白质[19]、靶向损伤线粒体伴侣或蛋白酶[20]、过量的ROS[21]和电子传递链损伤[22]等。激活UPRmt反应对于机体而言能够保护细胞免受更广泛的线粒体应激,可以调节蛋白质稳态[23]、引起代谢重组[15-16]、协调电子传递链组分和核基因组的表达[17]、激活免疫反应[24-25]、调节发育[26]、保护心脏[27]、延长寿命等[22,28-29]。
3 UPRmt的相关信号通路与传递
线粒体作为一个有着双层膜结构的细胞器,与细胞质之间的信息或蛋白质交流有着较为严格的独特性质,不能共享细胞质中普适的维持蛋白质稳态的分子机制。因此,在这种特殊的条件下,线粒体进化出了一种细胞器之间的网络信号传递机制,主要通过线粒体-细胞核之间的信号通路进行UPRmt的信号传导,从而达到调节线粒体蛋白质稳态、应对压力、保证生存等目的。
在线虫中,压力相关转录激活因子1(activating transcription factor associated with stress-1,ATFS-1) 是研究得最透彻的UPRmt相关信号分子[18]。它是哺乳动物细胞中转录激活因子ATF5的同源异构体,是一种有着碱性亮氨酸拉链结构域(basic leucine zipper domain,bZPIP) 的bZIP 蛋白[30]。作为UPRmt的主要激活因子,ATFS-1 蛋白既包含C 端的核定位序列(nuclear localization sequence,NLS),又具有N 端的线粒体定位序列(mitochondrial localization sequence,MLS),并且在不同的压力条件下有着相应的定位策略,极其适合将线粒体应激传递给细胞核[15]。在无压力的基础条件下,由于线粒体膜电位较高,蛋白质向线粒体的运输十分充沛,ATFS-1 通过N端的MLS 序列优先进入线粒体。一旦输入线粒体,其中的蛋白酶家族成员LonP 将其迅速降解至无法检测的水平。这时ATFS-1 并不会影响线粒体内部蛋白质的正常环境。然而,当线粒体感受到外界压力,线粒体蛋白质的输入通道关闭[31],ATFS-1蛋白N端的MLS将不能正常发挥功效,导致ATFS-1 蛋白在细胞质中大量积聚,促使其通过C端的核定位序列进入细胞核中[15]。当ATFS-1顺利进入细胞核后,会激活核基因组转录从而激活UPRmt,以达到对线粒体压力进行的反馈调控。染色质免疫共沉淀测序实验表明,ATFS-1 在进入细胞核后,促进了线粒体分子伴侣基因、ROS 解毒基因、糖酵解基因、先天免疫基因等上调,抑制了TCA和OXPHOS基因的产生,从而多方面调节线粒体蛋白质的稳态[32]。
然而,ATFS-1并不是激活UPRmt的唯一转录因子。在700 多个激活UPRmt的实验中,只有400 多个是通过ATFS-1转录因子激活[15]。因此,还存在着其他多种可以激活UPRmt的信号通路。泛素样蛋白5(ubiquitin-like protein 5,UBL-5)[23]以及果蝇前心室缺陷蛋白同源物1(defective proventriculus in drosophila homolog 1,DVE-1)[33]是在线虫中最早发现的可以引起UPRmt的信号分子。UBL-5在细胞核中的累积指示着线粒体应激反应,并且与后续的蛋白质翻译后修饰相关[23]。而DVE-1 作为一种转录因子,会和辅因子UBL-5 形成物理上的相互作用,组成复合物,从而在线粒体应激时调节核内参与线粒体细胞内稳态的分子伴侣mtHsp70 和Hsp60 的基因转录[33]。后续相关研究也发现,DVE-1 与UBL-5 的联合作用也受到组蛋白甲基转移 酶2 (histone methyltransferase-like 2,MET-2)与其辅因子异常细胞谱系蛋白LIN-65 的调控[28]。正因为UPRmt的完全激活过程需要染色质的重塑,表观遗传学相关的信号通路也是UPRmt调节的重要因素。除了甲基转移酶MET-2 和其辅因子LIN-65之外,两种组蛋白去甲基转移酶十文字域蛋白JMJD-3.1(jumonji domain protein 3.1)与JMJD-1.2(jumonji domain protein 1.2)也是参与UPRmt的重要信号分子[29]。在UPRmt的状态下,组蛋白H3K9通过MET-2 大量二甲基化,DVE-1 与LIN-65 在核中聚集,细胞核紧缩,整体基因沉默[28];与此同时,JMJD-3.1 与JMJD-1.2 共同作用,将三甲基化的H3K9去除两个甲基,使之成为具有活性的一甲基化的组蛋白,释放DNA[29],招募ATFS-1 和DVE-1等UPRmt相关的转录因子,大量调节相关分子的转录与翻译,从而更好地对抗压力,维持线粒体稳态。最新研究也发现,除了组蛋白的甲基化参与了UPRmt的调节,组蛋白的乙酰化也在UPRmt的调控中起着重要的作用。在线虫中组蛋白去乙酰化酶1(histone deacetylase,HDA-1)同样也是激活UPRmt所必须的分子,它和DVE-1 一起调节UPRmt相关基因的转录[34]。同年,另一研究也发现,包含有HDA-1 的核小体重塑和组蛋白去乙酰化酶复合 物NuRD (nucleosome remodelling and histone deacetylase)通过感受线粒体压力条件下乙酰辅酶A水平的降低,诱导DVE-1的核积累,从而降低组蛋白乙酰化的水平,激活UPRmt反应且延长了线虫的寿命[35]。以上研究表明,近年来生物学界研究的热点——表观遗传,也与UPRmt有着密不可分的联系。
此外,还有一种全新且独立于ATFS-1 与DVE-1 这两个常见转录因子的信号通路,也可以激活UPRmt。鞘氨酸肌醇激酶1 (sphingosine kinase-1,SPHK-1)[36]就是这个信号通路中最重要的分子之一,在感受到线粒体压力后,SPHK-1 会在线粒体外膜有一个快速且可逆的定位,且随之催化鞘氨醇(sphingosine,SPH)转化为1-磷酸鞘氨醇(S1P)。而S1P可能会作为一种分泌信号对压力进行传递,或者作为细胞内信号分子激活UPRmt。此方面相关的下游分子机制仍不明确,但为UPRmt的激活提出了一个全新的胞内信号通路,扩展了对UPRmt的理解(图1)。
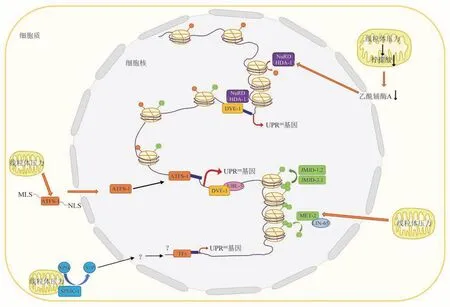
Fig.1 Cell-autonomous UPRmt-related signaling pathways图1 细胞自主性UPRmt相关信号通路
4 组织间的UPRmt传导通路
上述有关信号通路的研究都是发生在同一细胞内,是细胞自主性UPRmt的表现。然而,对于线虫线粒体功能研究表明,UPRmt信号不仅在那些正在经历线粒体应激的细胞内发生,还会在组织间进行传递,同时产生一种细胞非自主性的UPRmt。在最初的研究中发现,当特异性地降低或敲除线虫神经元中线粒体电子传递链(electron transport chain,ETC)复合物亚基时,不仅会激活神经组织自身的UPRmt,还会引发肠道组织的UPRmt[22]。由于技术原因,当时并未发现是哪些因子引起组织间的UPRmt,于是类比细胞因子“cytokine”,研究者将这些由于线粒体应激而产生的未知因子命名为“mitokine”。然而,随着线粒体稳态、线粒体UPRmt研究的深入,目前已经发现多种分泌因子参与了组织间UPRmt的信号传递。
5-羟色胺(5-HT)是一种抑制型的小分子单胺类神经递质,在神经元中有着广泛的分布。它是参与组织间UPRmt信号传递的重要分泌因子之一[37]。在神经组织中特异性表达多聚谷氨酰胺40(polyQ40)蛋白,可以诱导神经细胞产生UPRmt信号,激活突触囊泡外泌相关的钙依赖分泌激活因子同源物UNC-31和5-HT合成的关键酶——5-羟色氨酸羟化酶TPH-1,分泌并释放5-HT 进行组织间的信号传导,从而引起下游肠道内细胞非自主性UPRmt的激活[37]。
除了5-HT这种小分子神经递质依赖的信号传递途径外,神经肽也在细胞非自主性UPRmt中起重要作用。3个感受神经元ASK、AWC、AWA和1个中间神经元AIA会组成感知和传递线粒体应激的神经通路,在这个神经通路中特异性地引发UPRmt后,FMRF(苯丙氨酰-甲硫氨酰-精氨酰-苯丙氨酰胺,H-Phe-Met-Arg-Phe-NH2)类似神经肽FLP-2随之分泌释放,传递神经细胞中产生的线粒体压力信号,从而诱导肠道细胞非自主性UPRmt的产生[38]。
此外,胞间分泌的其他因子也可以参与细胞非自主性UPRmt的信号传递。Wnt信号通路常用于临近细胞之间的通信(旁分泌)或同个细胞自身的通信(自分泌),是一系列多功能且常见的分泌因子[39]。逆转运复合体中的组分液泡蛋白分选同源物VPS-35 与Wnt 分泌因子MIG-14 组成囊泡复合物,运输并分泌Wnt蛋白配体EGL-20,EGL-20蛋白再与肠道细胞膜上的Frizzled 受体相结合,共同稳定β连环蛋白(β-catenin),使之进入肠道细胞核内,从而与转录因子共同诱导肠道细胞产生细胞非自主性的UPRmt[40]。虽然机理并未清晰,EGL-20介导的细胞非自主性UPRmt产生的过程也是依赖5-HT的[40]。最新研究还发现,促卵泡激素G蛋白偶联受体1(follicle stimulating hormone receptor 1,FSHR-1)在神经元中的特异性表达也可以经由一系列信号传导,激活肠道细胞中的SPHK-1,促进SPH向S1P的转化,从而激活肠道细胞非自主性的UPRmt反应[41]。
经过系列实验,已经鉴定出了多种神经到肠道组织间通讯的信号分子,并且检测到了肠道组织中细胞非自主性UPRmt的激活,然而肠道组织感受UPRmt信号的表面受体及其信号传递机制仍不清晰。
以上研究均为细胞非自主性UPRmt信号在神经元与肠道细胞之间的传递。除此之外,这种通讯也可以在线虫生殖细胞与肠道细胞之间进行。在超级长寿的daf-2:rsks-1双突变线虫的生殖细胞中存在一系列蛋白质翻译水平的改变,导致线粒体中细胞色素C蛋白CYC-2.1下调,进而激活肠道细胞中的UPRmt,并显著延长线虫寿命[42]。然而,这种生殖细胞到肠道细胞之间进行传递的“g-mitokine”(germline-mitokine)仍未发现。
综上所述,细胞非自主性UPRmt的分子机制仍有许多不明确之处,不同组织之间可能会有更多全新的细胞非自主性传递机制,下游组织细胞表面受体的激活机制有待研究,还有更多线粒体相关的分泌因子“mitokine”亟待发掘(图2,3)。
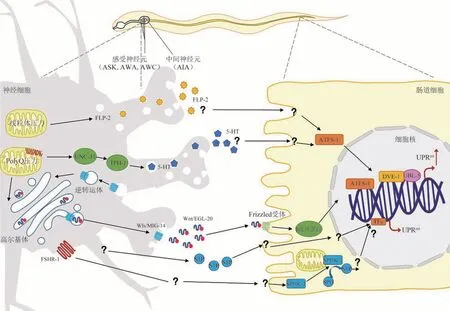
Fig.2 Cellular non-autonomous UPRmt-related signaling pathways between nerval and intestinal cells图2 神经细胞与肠道细胞间的细胞非自主性UPRmt相关信号通路
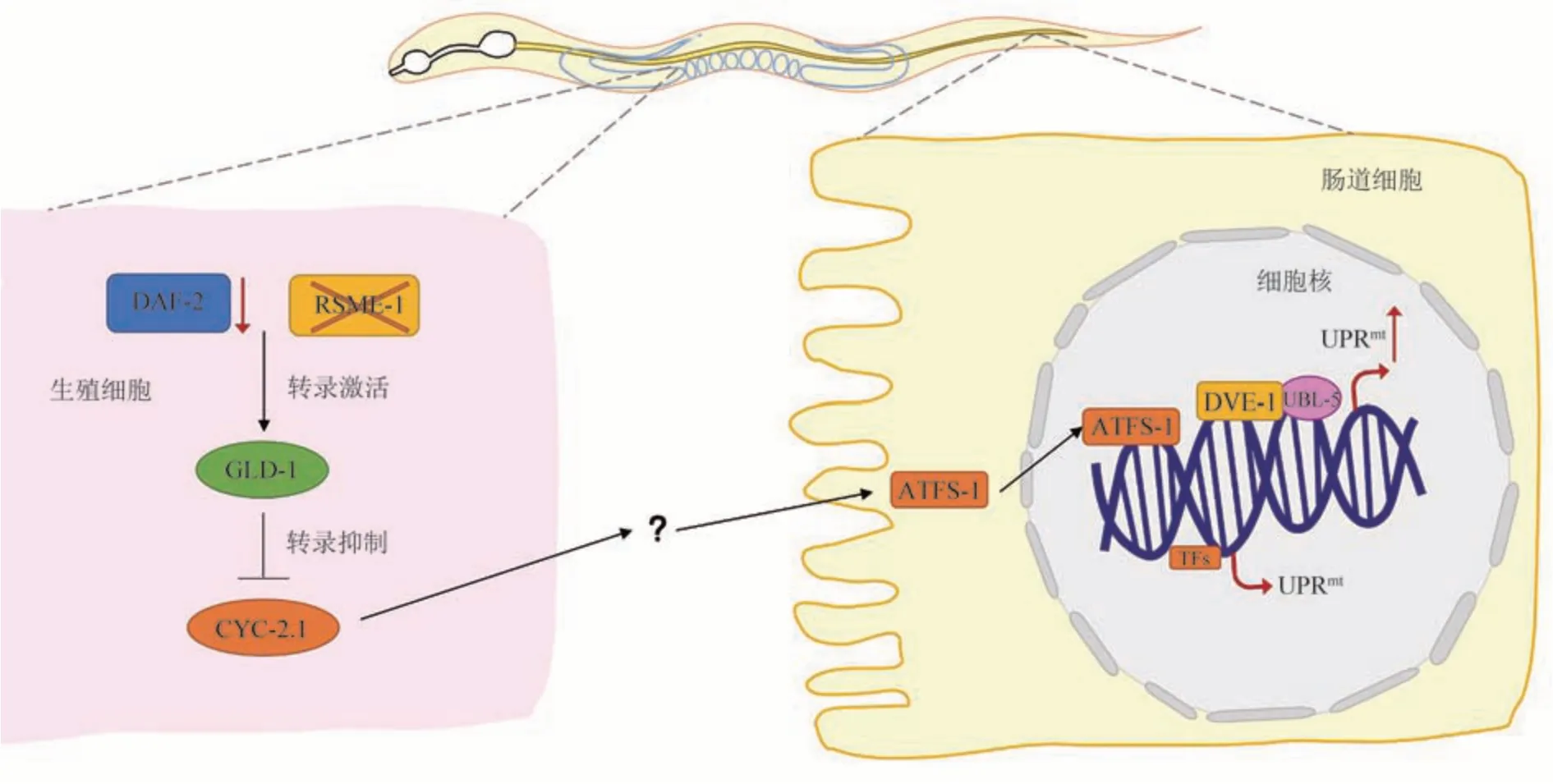
Fig.3 Cellular non-autonomous UPRmt-related signaling pathway between germline and intestinal cells图3 生殖细胞与肠道细胞间的细胞非自主性UPRmt相关信号通路
5 UPRmt与线虫寿命
UPRmt在细胞与组织水平的主要作用是缓解线粒体压力,投射在整个机体中,则会产生多种保护性功能。多项研究证明,引起UPRmt产生的相关信号分子在延长寿命方面发挥了重要作用。例如,过表达组蛋白脱甲基酶JMJD-2.1可以激活UPRmt并延长线虫的寿命[29],过表达Wnt/EGL-20可以激活细胞非自主UPRmt,并延长线虫寿命[40]。
适度进行线粒体平衡的扰动可以延长线虫的寿命,且与UPRmt的激活存在很强的相关性。其中重要的一点就是改变了线粒体内nDNA 与mtDNA 翻译蛋白之间的比例,产生了翻译的不均衡,从而激活UPRmt,并且对寿命增加有着极强的正反馈作用。如雷帕霉素抑制TOR 信号以改变nDNA 的翻译,以UBL-5依赖性的方式诱导UPRmt,并使线虫寿命延长了16%,这与ATP 水平增加,及nDNA/mtDNA 氧化磷酸化蛋白比率改变有关[43-44]。在表观遗传阅读器BAZ-2 和H3K9 甲基转移酶SET-6 基因突变的线虫中,也发现了nDNA/mtDNA 翻译水平的不均衡能够激发UPRmt并使线虫寿命延长[45]。此外,白藜芦醇作为具有延寿功能的添加药物,也可以诱导肝细胞中的nDNA编码蛋白失衡,并通过UBL-5 激发UPRmt,延长线虫寿命,并且增加呼吸作用,保持ATP水平和柠檬酸合成酶活性[43-44]。
另一方面,适当敲低mtDNA或干扰mtDNA的正常翻译也可以激活UPRmt并延长寿命。例如,当线粒体暴露在低剂量的溴化乙锭中时,会特异性地抑制mtDNA 的转录,使nDNA/mtDNA 翻译不均衡,诱发UPRmt,引起寿命的延长[46]。而特异性干扰mtDNA 编码的蛋白基因,如用RNAi 的方法降低 线 粒 体 小 核 糖 体 蛋 白MRPS-5[17]、 降 低OXPHOS复合物IV中的细胞色素氧化酶组装蛋白1(cytochrome oxidase assembly protein 1,CCO-1)[22]、或者降低富含亮氨酸的PPR 基序蛋白LRPPRC[47]表达水平,都可激活线虫的UPRmt,并延长寿命。
以上实验证明,通过多种方法影响线粒体内nDNA编码蛋白与mtDNA编码蛋白之间的不均衡,可以有效诱导UPRmt的发生,并且对寿命有着延长的效果。然而,UPRmt的激活并不是机体寿命延长的充分条件。例如,降低OXPHOS 复合物II 琥珀酸脱氢酶的线虫同源物MEV-1会激活UPRmt,但并不会延长线虫的寿命[22,48]。这可能是由于MEV-1的所有亚基是由nDNA单独编码,而不是由nDNA与mtDNA共同编码,降低MEV-1不会产生这种不同来源蛋白质复合物的配比失衡,因此并不会延长线虫寿命。此外,经过RNAi筛选实验也发现,并不是所有引发UPRmt的基因都可以延长寿命,且atfs-1功能获得突变(gain of function)中有UPRmt的组成性活性,但虫株并未显示出寿命延长的表型,其中的内在机制仍不清晰[49]。
除了激活UPRmt可以直接引起机体寿命延长之外,生物机体的长寿效果同时还需要线粒体其他多方面应激反应途径的激活。NAD+是调节代谢稳态的重要辅助因子,在老年线虫中表达水平大幅降低,增加NAD+的水平与过表达的人源sirtuin 同源物SIR-2.1 相互作用激活UPRmt;同时,增加的NAD+水平也激活了FOXO 调控细胞超氧化物歧化酶SOD-3通路,UPRmt和ROS防御的联合激活可以抑制线虫衰老相关的衰退,从而达到延长寿命的效果[50]。线虫神经细胞中的行为衰老调控基因rgba-1可以表达多种神经肽,它可与神经肽受体家族NPR-28 结合,并且通过调控SIR-2.1 激活的UPRmt通路调节线虫寿命[51]。线粒体产生的ROS也通过细胞程序化死亡机制共同增加寿命[52]。以上多种作用与UPRmt一起,共同维持线粒体稳态,调节寿命,从而保证生命体的健康存活。
6 总结与展望
近十年来,有关UPRmt的研究已经被证实远超出了其维持蛋白质稳态的基本功能。多种UPRmt信号直接参与蛋白质转录以及染色质重塑,也影响衰老相关的线粒体功能。这些研究表明,该领域将在调控模式和生理作用等方面得到进一步发展。然而,蛋白质未折叠反应也是一把双刃剑,虽然UPRmt的短期激活可能是有益的,并可能作为未来多种疾病的治疗靶点,但长期激活这种反应的影响尚不完全清晰,可能会影响线粒体的正常功能,这对生命体是否能健康存活也是一种挑战。由于UPRmt研究的主要机制已经较为清晰[53],近五年来,相关研究的重点主要放在表观遗传与UPRmt以及细胞间非自主性UPRmt的调控机制上。
本文系统性地总结了UPRmt细胞内与细胞间信号转导的相关机制,详细介绍了UPRmt激活的基础信号通路ATFS-1/DVE-1/UBL-5 等,并且也将近年来表观遗传对UPRmt的调控机制进行了总结,发现组蛋白的甲基化相关酶,如MET-2 和JMJD 家族,以及组蛋白的乙酰相关酶如包含有HDA-1的NuRD复合物都可以参与线粒体压力后的翻译,从而维持线粒体稳态,恢复机体的健康。另一方面,随着组织间细胞非自主性UPRmt激活机制研究的深入,已经发现多种“mitokine”参与组织间线粒体压力信号的传递,并且也发现多种组织,如神经细胞和生殖细胞,都参与到了这种细胞非自主性的信号传递中去。这很可能是生命体进化出的一种自我保护与修复的机制,若所有细胞都必须历经外界压力造成的损伤再进行“按部就班”地修复,必将对机体的健康与正常运作造成不可忽视的影响,而这种组织间的信号传递相当于一个“预警”机制,提前激活了下游压力正常组织的修复效应,从而更加迅速高效地应对外界损伤带来的挑战。
此外,本文也讨论了UPRmt对于寿命的影响。虽然,从压力回复等方面来说,UPRmt在很大程度上是有益的,通过激活相关基因的转录与蛋白质的翻译,维持蛋白质稳态,引起代谢重组,诱导一系列压力应激反应的激活,进而延长机体的寿命,但是这种激活对于延寿效果来说并不是充分且必要的条件。也许寿命延长的效果仅仅是UPRmt对于压力缓解带来的一种附加功效,或者说,轻微、短期、有限定条件的UPRmt可以延长寿命,而强烈、长期、普遍性的UPRmt激活可能与延寿效果并不相关。本文对UPRmt相关的机制以及最新研究做了详细且系统的介绍,为日后的进一步研究提供了参考。
最后,ATFS-1 和DVE-1 等相关转录因子如何独立激活UPRmt的机制尚未完全清晰,UPRmt与组蛋白修饰相关的机制也有疑点亟待发掘,细胞非自主性UPRmt的组织间传递的信号分子也并不完全明确,压力信号在胞质与细胞核内传导方式依旧有极大的发掘空间,UPRmt有关寿命的调控机制也仍需研究,这些问题还需要日后更多的实验使人们对UPRmt领域的认知更加完善。
