植物嫁接砧木与接穗互作机制研究进展
2022-05-16孟景祥魏永成
张 捷,艾 迪,,孟景祥,魏永成,张 勇
(1 东北林业大学 园林学院,黑龙江 哈尔滨 150040;2 中国林业科学研究院热带林业研究所 热带林业研究实验室,广东 广州 510520)
嫁接技术在园艺作物,尤其在果蔬类作物上应用广泛[1]。与自根植物相比,通常嫁接植物所选砧木的根系更发达,可以改善植株对水分和营养物质的吸收利用,继而对接穗产生影响[2]。嫁接植物的砧木可以通过一些细胞和分子水平上的生理生化响应,对接穗生长、有机物积累、激素产生与运输、抵御非生物胁迫以及基因表达等产生显著影响[1,3]。近年来,砧穗互作一直是研究热点,并且随着分子生物学的发展,在分子水平上探究砧穗互作的关系逐步深入。因此,完善砧穗互作机制方面的研究,对选择性状优异的砧木、获得更加合理有效的砧穗组合及促进嫁接技术在园艺作物中高效应用具有重要意义。本研究从水分和矿质离子、植物激素、抗性以及嫁接亲和性、基因表达、RNA长距离运输和DNA甲基化等方面,综述了砧穗互作的生理及分子机制,以期为深入研究砧穗互作机制提供借鉴。
1 砧穗互作的生理机制
1.1 水分和矿质离子的吸收与利用
水分是植物生长所必需的物质,其吸收与转运会影响离子、激素以及一些有机物质的运输,与植株的生长和发育息息相关[3-4]。一般来说,水分运输能力与嫁接植株所处的生长发育阶段有关。Johkan等[5]研究发现,年老的甜椒(Capsicumfrutescens)植株因嫁接部位木质部老化,造成砧穗连接处发育不良,导致叶片气孔阻力和水势较幼株低。接穗和砧木的基因型也对植株的水分利用至关重要,尤其是砧木。张志焕等[6]发现,在缺水条件下,番茄(Lycopersiconesculentum)砧木‘606’嫁接苗的叶片水势和水分利用效率显著高于‘112’(砧木品种)和自根番茄,说明‘606’砧木的根系吸水能力更强,从而导致嫁接后植株对水分的利用效率更高。同样,在甜瓜(Cucumismelo)嫁接中,Agele等[7]发现砧木基因型对嫁接组合和接穗水分关系的影响更为显著。砧木和接穗的相互作用也会改变矿质营养元素的吸收效果,进而影响植株的生长。在番茄嫁接组合R/GUO(R为接穗品种,GUO为砧木品种)和R/ZA中,幼苗根和叶片K、Ca、Mg、Na含量显著高于自嫁接对照植株R/R[8]。嫁接在同一种砧木上,牛油果接穗品种‘Hass’(Perseaamericana)叶片的Ca、Mg、Na、P和K含量明显高于接穗品种‘Ettinger’和自嫁接苗;与嫁接到原产印度的砧木相比,‘Hass’嫁接到原产墨西哥的砧木品种后叶片Cl的含量较高,但Mg和Mn含量较低[9]。因此,植株中大量和微量营养元素含量受嫁接砧木和接穗互作的影响,且嫁接后一些性状优良的砧木可以消除或削弱环境条件对植株吸收养分元素的不利影响[10-12]。如Venema等[13]发现,自嫁接番茄植株的总氮含量随着根区温度的降低而升高,但不同接穗嫁接的番茄无明显差异。
1.2 植物激素的调控作用
植物激素是砧穗互作调控嫁接植株生长的机制之一[14]。研究发现,生长素(IAA)与细胞分裂素(CTK)、乙烯(ETH)、茉莉酸(JA)和赤霉素(GA)等激素在嫁接愈合过程中会积极参与愈伤组织的形成[4]。IAA与GA和CTK具有协同作用,能刺激细胞分化,促进维管束形成及木质部与韧皮部的重新连接,从而恢复生长素的对称性。而GA与IAA互作,会通过细胞分化刺激愈伤组织的出现[14-15]。龙伟等[16]发现,油茶(Camelliaoleifera)砧穗受到嫁接创伤后,‘18号’品种砧木创伤面上的反式玉米素核苷(tZR)、玉米素(Zeatin)和水杨酸(SA)含量总体高于‘53号’品种,这些激素变化使嫁接在‘18号’上的接穗愈伤组织发育良好。
激素会参与嫁接植物的生长发育[17]。Galvez等[18]发现,辣椒(Capsicumannuum)砧木‘Atlante’品种可以通过CTK和脱落酸(ABA)之间的有效平衡提高接穗的活力,而砧木‘Terrano’会限制GA在芽中的积累,导致株高降低。孟红志等[19]也发现,嫁接在苹果(Malusdomestica)中间砧‘SH40’上的(IAA+ZR+GA)/ABA在果树生长发育期显著高于自根砧处理,生长较为旺盛。此外,激素可能会影响嫁接组合对环境胁迫的抵抗能力[20-21],Etehadnia等[22]研究表明,使用耐盐接穗或具有高ABA水平的砧木嫁接,可以有效改善盐胁迫对植物的不利影响[23-24]。Albacete等[25]也证明,番茄栽培品种嫁接到耐盐野生番茄上后,植株通过调节矿质离子和激素的供应而使盐胁迫下番茄作物的生产力显著提高。
1.3 抗 性
盐碱、干旱、重金属、土壤贫瘠、高温、低温等非生物胁迫,以及病虫害等生物胁迫,均会造成植物体内离子毒害、渗透失调、代谢紊乱、质膜破坏等不利影响[26],但这些不良反应可以通过嫁接得以缓解。嫁接后,具有优良基因型的砧木或接穗能通过离子协调、质膜保护、抗氧化物质、激素转运、渗透调节物质(可溶性糖、可溶性蛋白等)缓解胁迫因子对植物体造成的伤害[1,3],提高其抗性,从而促进植物生长发育(图1)。有大量研究证实,砧穗间相互作用对增强植物对外界胁迫的抵抗能力有重要作用。
以抗盐响应为例,盐碱环境会严重影响植物生长发育,既有研究提供了多种有助于植物耐盐的方式,嫁接是其中一种[2]。嫁接使植物通过渗透物质调节来缓解盐损伤,渗透调节物质包括K+、Na+、Cl-等无机离子和脯氨酸、可溶性糖、可溶性蛋白等有机溶质[2,8]。Li等[21]发现,苹果品种‘G935’砧木根系可以更加有效地排除细胞中的Na+,阻止其向地上部分运输,降低离子毒性,从而提高接穗的耐盐性。Shahid等[27]发现,耐盐的柑橘(Citrusreticulata)砧木有助于渗透调节物质(脯氨酸和甘氨酸甜菜碱)积累,且耐盐砧木嫁接植物的超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、谷胱甘肽还原酶(GR)等抗氧化酶活性显著增强,说明耐盐砧木可通过提高植株的抗氧化酶活性来缓解盐伤害。黄瓜(Cucumissativus)、柠檬(Citruslimon)的嫁接组合在盐胁迫下也显示出抗氧化酶活性升高等响应机制[26,28-29]。此外,光合作用路径改变也会对植物耐盐性产生积极影响。桑树(Morusalba)嫁接苗与盐胁迫下自根幼苗相比,叶片的光化学活性和量子传递速率更高,有效地减少了由活性氧(ROS)介导的氧化反应[30]。孙士涛[31]研究发现,盐胁迫下,嫁接在南瓜(Cucurbitamoschata)砧木上的黄瓜接穗叶片气孔对ABA的敏感性较自嫁接植株提高,引起气孔快速关闭,蒸腾作用降低,水分散失减少,从而缓解植株萎蔫,显著增强了嫁接黄瓜的盐胁迫适应性[32]。因此,植物激素也可能是植株对抗盐胁迫的关键因素之一,但其分子机制还有待进一步充分证实[19]。

图1 嫁接后砧穗互作对植物抗性调节示意图Fig.1 Schematic diagram of regulation of plant resistance through rootstock-scion interaction after grafting
2 砧穗互作的分子机制
Kurotani等[33]发现,细胞壁重建基因的调控作用可以提高嫁接成功率。如果植物拥有这种基因或被培育成大量拥有这种基因的品种,嫁接的种间障碍就可以消除,从而可以更有效地利用远亲双方的优势。近年来,有关砧木接穗相互作用的分子机制研究也逐渐增多,基因表达和蛋白质功能是生理变化的基础[34-35],准确地认识基因和蛋白质对于水分、激素、酶等生理变化的影响,将有助于理解砧穗间相互作用的机理。砧穗互作的分子机制包括基因表达变化、蛋白质活性变化以及RNA分子长距离运输等[36]。图2显示,砧木和接穗间的蛋白质活性变化、RNA等物质跨越嫁接结合长距离运输都会影响基因表达,进而影响植物的表型;小RNA的遗传信息传输与DNA甲基化相关,且DNA甲基化的变化与基因表达相关,同样影响植物的表型[37]。
2.1 嫁接亲和性
在嫁接伤口愈合期间,砧木与接穗的相互作用涉及复杂的生理生化和分子机制。研究发现,蛋白质能够从接穗的芽细胞移动到砧木的根细胞,从而调节植物的重要生理过程[38]。Zhang等[39]发现,通过上调植物质膜内在蛋白(PIP1B),可以增加细胞含水量,促使细胞伸长,促进愈伤组织的形成。在山核桃(Caryaillinoinensis)嫁接相关miRNAs中,miRS26参与了愈伤组织的形成,miR164、miR156、miRS10和miR166与维管束的分化有关,这说明miRNA在山核桃嫁接成活中起着重要作用[40]。
有研究表明,西瓜(Citrulluslanatus)嫁接后大多数IAA转运蛋白调控基因的表达水平在砧穗连接形成时发生变化,并可有效促进伤口愈合和维管形成[38,40]。Zhai等[41]发现,与自根植物相比,嫁接在含有生长素合成基因(iaaM)砧木上的烟草(Nicotianatabacum)接穗中,内源性生长素水平升高,生长素应答性DR5::GUS表达量增加,这表明生长素从根茎向上转移到接穗组织,通过增加砧木和接穗组织中参与嫁接愈合的基因表达量来促进嫁接结合处形成。此外,愈伤组织的生长与特定代谢物的产生有关。Prodhomme等[42]在葡萄(Vitisvinifera)藤中进行代谢物分析发现,砧穗接合界面氨基酸的产量以及二苯乙烯化合物的积累有所增加。而Amri等[43]发现,桃子(品种‘SG’)/李子(品种‘Damas GF 1869’)的远缘嫁接中,砧木‘Damas GF 1869’诱导接合处的可溶性糖、淀粉、酚类化合物和抗氧化物含量显著增加,但这些物质不利于砧穗结合处愈伤组织的形成,可能是导致SG/Damas GF 1869嫁接不亲和的主要原因[44]。
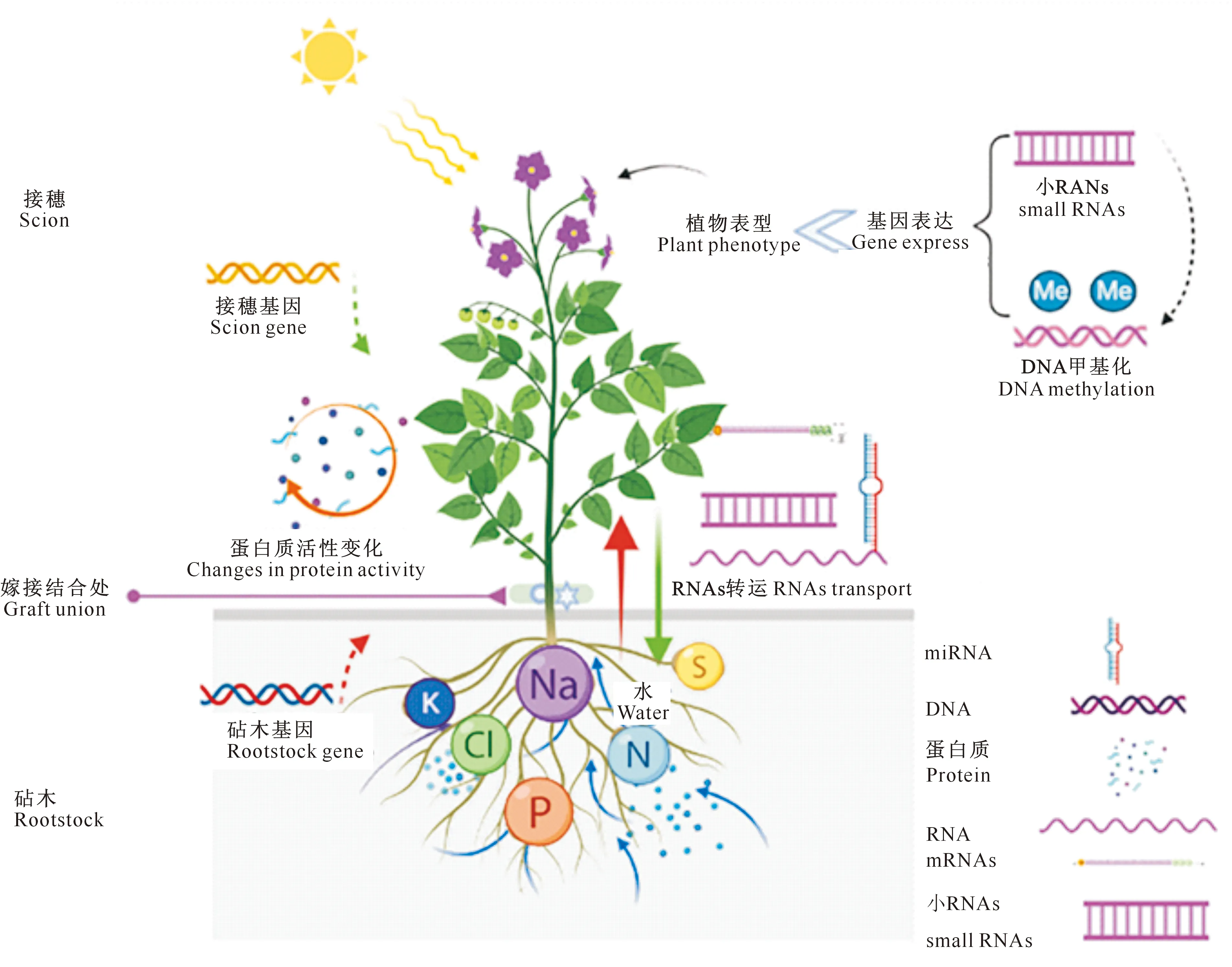
图2 嫁接植物中的分子作用机制示意图[37]Fig.2 Schematic diagram of molecular mechanism of action in grafted plants[37]
2.2 嫁接对基因表达的影响
植物嫁接中遗传基因或信息的表达对植物的生长发育有重要的调控作用[45]。Garcia-Lozano等[45]研究了嫁接在葫芦上的西瓜接穗的转录组变化,发现大量与成熟、品质以及胁迫反应相关的基因在2种嫁接组合的组织中均有不同表达。苹果品种‘M.9’砧木叶片和根部YUCCA1Oa和IPT3A基因的表达量较低,导致其上的嫁接组合矮化效果更显著[46-47]。在低K+环境下,棉花(Gossypiumspp.)嫁接组合中4个特定的K+转运蛋白基因(3个GhHAK5和1个GhKUP3)的表达水平均升高,促使植物能够应对K+缺乏症[48-49]。Zhao等[50]对嫁接黄瓜果皮的转录组进行了分析,结果发现,不同的南瓜砧木可以诱导参与糖和芳香化合物合成的基因转录组发生显著变化,从而影响果实品质。与自嫁接对照相比,马铃薯(Solanumtuberosum)砧木在番茄接穗上诱导的表型变化较小,对差异表达基因的影响也较小。然而,番茄接穗对马铃薯砧木有重大影响,导致数千个基因差异表达,其中包括一些参与激素信号和激素通路的基因[20]。在盐胁迫下,南瓜砧木可诱导嫁接黄瓜植株体内ABA生物合成的相关基因NCED2、ABCG22、PP2C和SnRK2.1的表达水平均上调,从而增强了接穗对根向地上部传递ABA的敏感性,降低了叶片的蒸腾速率和气孔导度,进而提高了黄瓜的耐盐性[51]。在柑橘嫁接组合TAL/RL中,与抗氧化酶相关的基因表达量上调,从而阻碍过多的ROS产生,赋予柑橘类植物更强的耐旱性[52]。此外,Bennici等[53]研究表明,柑橘砧木‘C35’对接穗‘Tarocco’中CiFT2基因过表达介导的成花诱导有显著影响。
2.3 RNA长距离运输以及DNA甲基化
目前已证实,miRNAs会影响植物开花、器官的形态及对各种激素信号的响应[53-54]。此外,miRNAs也会影响砧木和接穗之间的相互作用。Ahsan等[55]发现,砧木通过影响牛油果接穗中miRNA和mRNA的丰富度来促进嫁接结合处的形成,而miR172、miR156和miR156的靶基因SPL4与牛油果接穗和砧木成熟度直接相关。另外,大量研究表明,嫁接会影响植株的DNA甲基化,而在嫁接植物中移动的siRNA会调控DNA甲基化,从而诱导可遗传的调控转录基因沉默。同样,DNA甲基化也会改变植物的基因表达[56]。Xanthopoulou等[57]发现,葫芦砧木‘Munchkin’改变了嫁接组合的整体DNA甲基化模式和特定miRNAs的表达,导致‘Big Moose’接穗的果实表型发生显著变化。
RNA砧穗间运输对嫁接植株抗性也会产生影响。Sidorova等[58]对小RNA(sRNA)库的高通量测序表明,转基因李子砧木中构建的特异性干扰小RNA(siRNA)的积累达到2%以上,当嫁接转基因组织时, siRNA水平升高,植株对李痘病毒产生抗性,并阻止病毒通过砧木进入接穗。同样,Spanò等[59]发现,将易感病毒的番茄嫁接到抗性番茄品种砧木上,接穗积累的病毒RNA较少。面对干旱胁迫时,葡萄砧木‘1103 Paulsen’通过调节miRNA的表达增强了接穗‘ENTAV 115’的抗旱性[60],南瓜嫁接中也有类似报道[37]。但miRNAs表达量增加与胁迫适应的因果关系,以及不同砧木和接穗是否直接参与植株的抗性变化尚待探究[56,60]。
3 展 望
优良砧木具有发达的根系,能促进接穗的营养生长,提高果实品质,接穗生长又可以为根系提供更多的营养,从而达到砧穗间相互促进的效果。砧木接穗的相互作用对植物生长、开花、坐果率、产量和果实品质的影响复杂,涉及的机理广泛。水分、营养、植物激素等生理机制较为明确[2],但这些调控因子的相互影响增加了砧穗互作的复杂性,尤其是植物激素,其种类复杂多样,且具有相关性,在砧穗互作的机制及与基因关联性方面有待深入研究。近几年分子生物学技术的发展和应用为深入理解砧木与接穗的关系提供了可能,但仅局限于瓜果类。因此,砧穗间相互作用对不同多年生植物(包括观赏类植物)的甲基化模式、表观遗传标记及其维持是未来研究的重要领域。另外,植物嫁接是否会引起可遗传的表观遗传变化,从而对基因表达变异性产生显著影响,进而影响嫁接植株的表型,也可能会成为未来的研究热点。
