干旱胁迫及复水对M9T337苹果砧木苗生理特性的影响
2022-05-14赵文哲刘晓杜桂英邓文鹏宋新卫肖伟李玲
赵文哲,刘晓,杜桂英,邓文鹏,宋新卫,肖伟,李玲
(1.作物生物学国家重点实验室/山东果蔬优质高效生产协同创新中心/山东农业大学园艺科学与工程学院,山东 泰安 271018;2.五莲县农业农村局,山东 五莲 276800;3.肥城市新城街道办事处,山东 肥城 271600)
世界上有30%以上的土地处于干旱和半干旱地区,其他地区由于气候的不同也出现了季节性干旱[1]。我国北方自然降水少,农业用水不足,已成为限制农业生产的主要因素。果树生长发育和产量形成受水分影响很大,而我国北方落叶果树种植区往往无灌溉条件或者灌溉条件较差,严重影响果树产量和果实品质,干旱胁迫已成为影响北方果树生产的重要环境因子[2]。
在遭受干旱胁迫时,植物体会进行一系列生理生化调节来抵御水分胁迫对自身的伤害[3]。Redillas等研究发现,干旱胁迫下存在叶绿素含量先升高后下降的现象[4],这可能有助于提高胁迫初期植株的捕光能力,促进光合机构的能量连通性,在一定程度上防御PSⅡ发生光抑制[5]。光化学猝灭系数(qP)、最大光化学效率(Fv/Fm)、实际光化学效率(ΦPSⅡ)的变化可以反映光系统Ⅱ的反应活性及植物的光能利用效率,在干旱胁迫条件下三者均下降,胁迫程度越严重下降越显著[6]。渗透调节是植物抵御逆境胁迫的重要方式之一,其主要表现是细胞内渗透势的下降和渗透调节物质的积累[7]。干旱胁迫下,植物体内可以积累大量的游离脯氨酸以增强细胞的渗透调节能力,游离脯氨酸含量与植物的抗旱性呈正相关关系,可作为植物抗旱性评价的指标;另外,干旱胁迫强度直接影响植物可溶性蛋白含量的变化,胁迫强度越大,其积累越明显[8]。
苹果(Malus domestica)是蔷薇科苹果属多年生木本植物,为落叶乔木,主要通过嫁接进行繁殖,因此,砧木对逆境的适应性直接影响嫁接苹果品种的适应能力。Liu等研究发现,当‘嘎啦’嫁接到抗旱砧木上时,其水分利用率明显提高[9]。因此,在干旱、半干旱地区,选用较为抗旱的砧木嫁接苹果品种,可使其更好地适应缺水环境[10]。
M9T337是荷兰木本植物苗圃检测服务中心选育出来的脱毒M9矮化砧木优系,比M9砧木矮化程度高20%[11]。本试验即以苹果砧木M9T337为试材,从叶绿素含量、叶绿素荧光参数、根系活力、渗透调节物质含量等角度研究该砧木对干旱胁迫的生理响应,以期为砧木抗旱性评价体系的建立和高效抗旱砧木资源的筛选提供理论依据。
1 材料与方法
1.1 试验材料
试验于2018年6—10月在山东农业大学园艺实验站进行。选取生长状况良好且树势基本一致的一年生M9T337苹果砧木苗进行试验。
1.2 试验设计
2018年6 月中旬将所选一年生M9T337苹果砧木苗移栽入上口径34 cm、下底径19 cm、高23 cm的塑料盆中,盆栽土壤为山东农业大学苹果中心园土,每处理25盆,每盆4株,每处理共100株;8月中旬将试材移入避雨棚内进行持续干旱处理,以正常供水为对照;胁迫第18天,供试植株叶片出现萎蔫时,进行复水处理,浇透水。
分别于干旱处理的第0、6、12、18、24天测定新梢中部功能叶片荧光参数,并取样,每次随机取15株,迅速放入冰盒,带回实验室于-80℃冰箱保存备用;同时取土样进行土壤含水量测定。
1.3 测定项目及方法
1.3.1 叶绿素荧光参数 采用英国Hansatech公司的FMS-2型便携脉冲调制式荧光仪测定M9T337砧木叶片光适应下的稳态荧光(Fs)、最大荧光(Fm′)、最小荧光(Fo′),充分暗适应20 min后测量叶片的初始荧光(Fo)、最大荧光(Fm)和PSⅡ原初光能转化效率Fv/Fm。通过公式计算得到PSⅡ实际光化学量子产量(ΦPSⅡ)和光化学淬灭系数(qP)。qP=(Fm′-Fs)/(Fm′-Fo′),ФPSⅡ=(Fm′-Fs)/Fm′。
1.3.2 叶绿素含量 采用分光光度法测定。取新鲜叶片0.2 g于离心管中,加95%乙醇20 mL浸泡,用铝箔纸将离心管包住后置于黑暗条件下24~36 h至叶片发白,取出于663、646、470 nm下比色(以95%乙醇为对照)。重复3次。
1.3.3 根系活力 采用氯化三苯基四氮唑还原法(TTC法)进行测定[12],结果用单位鲜重根系还原量表示。
1.3.4 渗透调节物质含量 可溶性糖含量采用蒽酮比色法测定,淀粉含量采用酸水解法测定,可溶性蛋白含量采用考马斯亮蓝(G-250)法测定[13]。
1.3.5 相对电导率 使用电导率仪测定相对电导率。鲜样用自来水冲洗后取0.5 g,加入20 mL蒸馏水,抽气3次,每次20 min,取出摇动,室温保持3~4 h,测电导率S1;封口沸水浴10 min,冷却,平衡10 min后测S2,同时测定蒸馏水S0,根据公式计算相对电导率:相对电导率(%)=100×(S1-S0)/(S2-S0)。
1.3.6 半胱氨酸蛋白酶(Casp)活性 参照文献[14]的方法进行测定。称鲜叶样约1 g在液氮中充分研磨,加入1×PBS(pH 7.2~7.4)9 mL匀浆,然后2500 r/min离心30 min,取上清,用Plant Casp ELISA Kit试剂盒进行半胱氨酸蛋白酶活性测定,在450 nm波长处测定各孔的OD值,重复3次。
1.4 数据处理与统计分析
采用Microsoft Office Excel 2010进行数据处理,采用GraphPad Prism 6作图,采用IBM SPSS Statistics 21对数据进行方差分析和显著性检验。
2 结果与分析
2.1 干旱及复水处理对M9T337苹果砧木叶片叶绿素含量的影响
如图1所示,干旱胁迫能够促进M9T337砧木叶片中叶绿素的积累,干旱胁迫第12天时,叶绿素含量最高,显著高于对照31.5%,此时土壤含水量为8%左右;干旱胁迫第18天的叶绿素含量次之,也显著高于对照;其余时间干旱处理的叶绿素含量与对照差异不显著。复水第6天的叶绿素含量较干旱胁迫第18天低,但仍高于对照,差异未达显著水平。
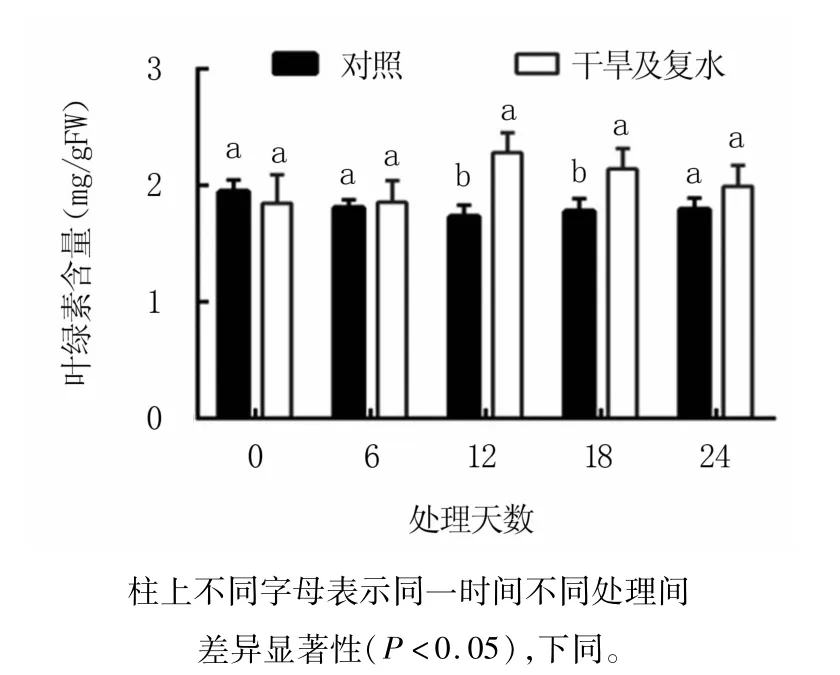
图1 干旱及复水过程中M9T337苹果砧木叶片的叶绿素含量
2.2 干旱及复水处理对M9T337苹果砧木叶片荧光参数的影响
自然干旱过程中M9T337苹果叶片光适应下的最大荧光Fm′逐渐下降,胁迫第18天时最低,比对照低41.0%;复水后Fm′快速上升,至复水第6天时已基本恢复到未进行干旱胁迫时的值(图2A)。干旱胁迫6天后,光化学猝灭系数(qP)和PSⅡ实际光化学效率(ΦPSⅡ)快速下降,第18天时值最低,与对照相比分别降低了53.1%和48.8%;复水后逐渐恢复,至复水第6天时已接近胁迫第6天的值(图2 B、C)。干旱胁迫与复水过程中PSⅡ最大光化学效率除在干旱12天时略有降低外,其余时间变化很小,与对照相比差异不大(图2D)。
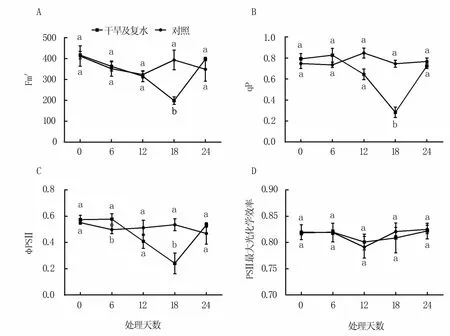
图2 干旱及复水处理对M9T337砧木叶片叶绿素荧光参数的影响
2.3 干旱及复水处理对M9T337苹果砧木叶片中渗透调节物质含量的影响
2.3.1 对可溶性糖含量的影响 由图3可以看出,干旱胁迫12天后M9T337砧木叶片中可溶性糖含量上升,第12、18天分别比对照高12.51%和49.93%,第18天时显著高于对照。复水后其含量明显下降,复水第6天的可溶性糖含量显著低于对照,降幅为38.37%。
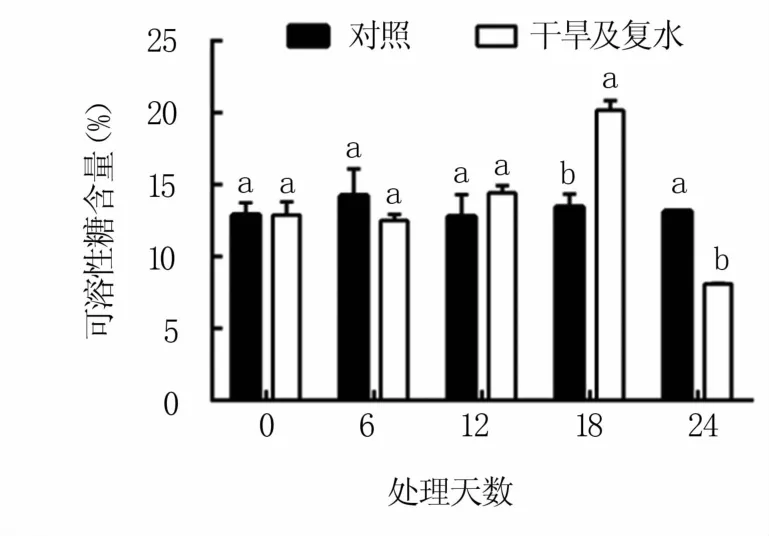
图3 干旱及复水处理对M9T337苹果砧木叶片可溶性糖含量的影响
2.3.2 对淀粉含量的影响 如图4所示,干旱胁迫显著降低M9T337叶片中的淀粉含量,尤其胁迫第12、18天时大幅下降,分别比对照降低85.4%和92.5%。复水后淀粉含量上升很快,第6天时淀粉含量仅比对照低6.8%,差异不显著。
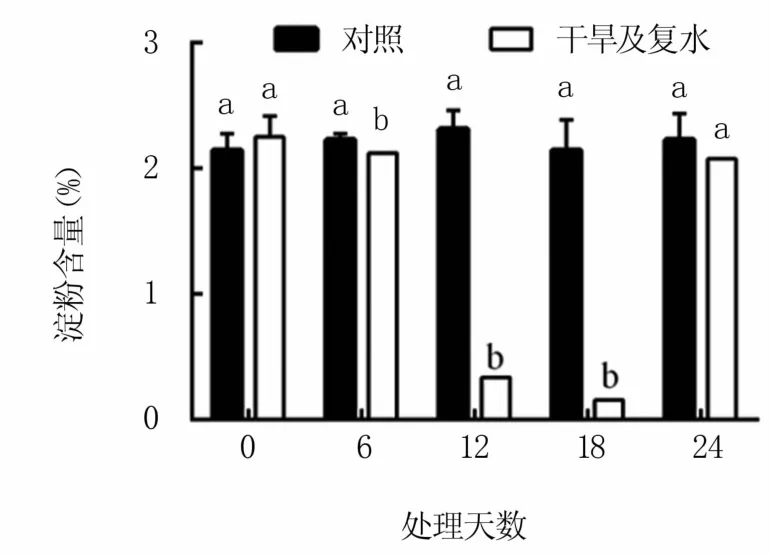
图4 干旱及复水处理对M9T337苹果砧木叶片淀粉含量的影响
2.3.3 对可溶性蛋白含量的影响 由图5可见,干旱胁迫显著提高M9T337叶片中的可溶性蛋白含量,且随胁迫时间的延长逐渐升高,第18天时可溶性蛋白含量达到最高值,是对照的3.7倍。复水后又快速下降,至复水第6天时与对照相当。
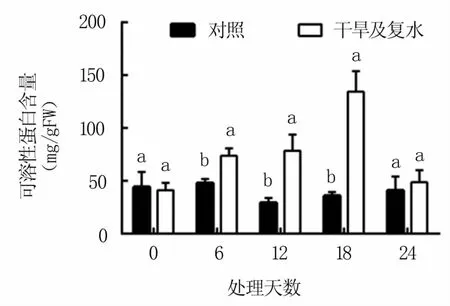
图5 干旱及复水处理对M9T337苹果砧木叶片可溶性蛋白含量的影响
2.4 干旱及复水处理对M9T337苹果砧木叶片相对电导率的影响
由图6可知,干旱胁迫12天内,M9T337的相对电导率变化较小,与对照差异不显著,但之后迅速上升,第18天时达到对照的2.53倍。复水后相对电导率又明显下降,复水第6天时与对照差异不大。
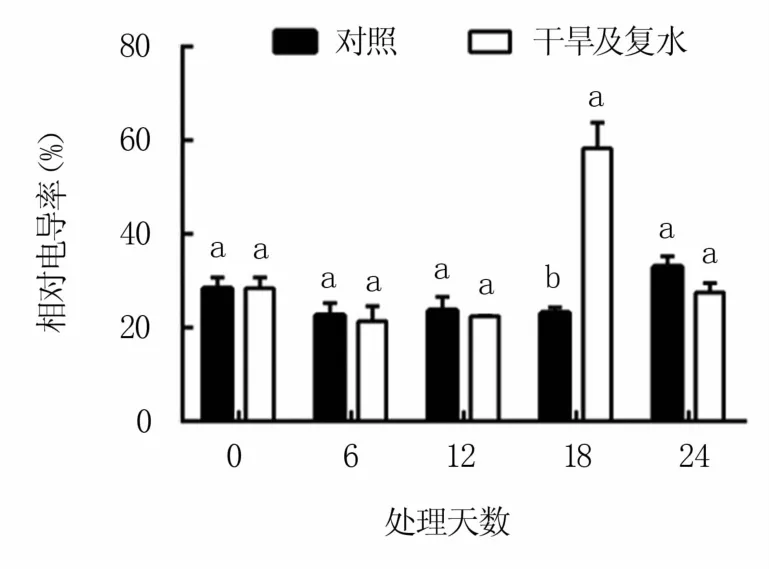
图6 干旱及复水处理对M9T337苹果砧木叶片相对电导率的影响
2.5 干旱及复水处理对M9T337苹果砧木叶片半胱氨酸蛋白酶活性的影响
如图7所示,M9T337苹果砧木叶片中的半胱氨酸蛋白酶活性在干旱及复水过程中变化较小,干旱18天时略有降低,复水后又逐渐恢复,与对照差异不显著。
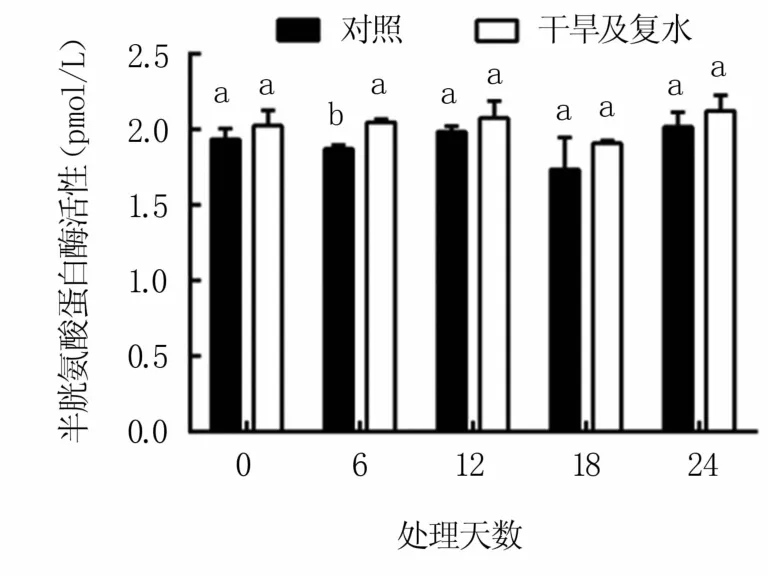
图7 干旱及复水处理对M9T337苹果砧木 叶片半胱氨酸蛋白酶活性的影响
2.6 干旱及复水处理对M9T337苹果砧木根系生长的影响
2.6.1 对根系形态的影响 如图8所示,干旱胁迫6 d内,植株细根较多且大多鲜嫩富有活力;胁迫第12天时,植株根部出现木质化;胁迫第18天时根系木质化严重,褐化,细根少;复水6 d时的根系形态变化不明显。

图8 干旱及复水过程中M9T337苹果砧木苗根系形态
2.6.2 对根系活力的影响 如图9所示,随着干旱胁迫时间的延长,M9T337砧木苗根系活力呈下降趋势,且在干旱6天后快速下降,干旱第12天的根系活力较对照下降67.5%,第18天接近于零。复水后M9T337砧木苗根系活力有所回升,但第6天时仍显著低于对照,约是对照的1/2。
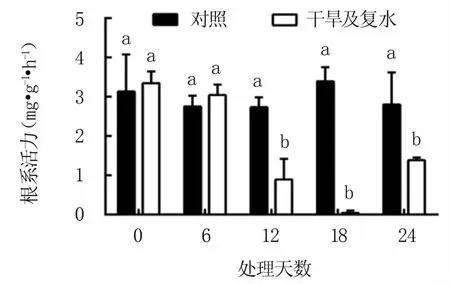
图9 干旱及复水过程中M9T337苹果砧木苗的根系活力
3 讨论与结论
3.1 干旱及复水过程中M9T337苹果砧木叶片叶绿素含量及叶绿素荧光参数的变化
王斌等研究表明光合和叶绿素荧光参数可作为评价木本植物抗旱性的有效指标[15]。叶绿素含量高低可以反映植物的光合作用能力强弱,而叶绿素荧光变化可以在一定程度上反映环境因子对植物光合作用的影响[16]。逆境条件下,叶绿体类囊体膜结构受到影响,会降低光系统(尤其PSⅡ)的活性,导致光系统对能量吸收、捕获和转换的能力降低,非光化学耗散比率增加[17]。本研究结果表明,干旱胁迫能增加M9T337叶片的叶绿素含量,至胁迫第12天时显著高于对照,达到最高值,第18天时稍有降低,仍显著高于对照;复水后的叶绿素含量高于对照,但与对照差异不显著。干旱胁迫能够降低植物的光化学猝灭系数(qP)、光适应下最大荧光(Fm′)和PSⅡ实际光化学效率(ΦPSⅡ),说明干旱胁迫使PSⅡ反应中心活性及光化学反应能力下降;但PSⅡ最大光化学效率(Fv/Fm)与对照差异较小,说明干旱胁迫对M9T337苹果砧木苗的最大光能转换效率影响较小。
3.2 干旱及复水处理对M9T337苹果砧木叶片细胞渗透调节的影响
渗透调节在植物对逆境的适应过程中发挥着重要作用[18]。渗透调节物质如可溶性糖、可溶性蛋白等的积累能降低渗透势,减轻逆境胁迫对植物细胞的伤害,维持细胞内代谢的正常进行[19,20]。淀粉是葡萄糖分子的高聚体,是植物体内主要的碳素贮藏物质,占有较高的比例。
本研究发现,干旱胁迫能增加M9T337砧木叶片中的可溶性蛋白和可溶性糖含量,降低淀粉含量,尤其胁迫6天后变化更为剧烈,但复水均能扭转这种趋势,使三者含量趋于正常。干旱胁迫下叶片中淀粉含量普遍降低,原因可能是一部分被淀粉酶降解,另一部分则水解转化为可溶性糖,从而增加可溶性糖的累积。
蛋白质在细胞生理代谢过程中有重要的催化功能,其含量在一定程度上能够反映植物内部代谢的活跃程度[21,22]。半胱氨酸蛋白酶作为蛋白水解酶,能够降解逆境条件下的受损蛋白,为新蛋白的合成提供肽段或游离的氨基酸,而且存在于叶绿体中的半胱氨酸蛋白酶可以降解光合和光呼吸中的重要酶RuBisCO[14]。在本试验条件下,干旱和复水对M9T337叶片中的半胱氨酸蛋白酶影响较小,仅在干旱胁迫第18天时略有下降。通过分析相对电导率的变化,仅在处理第18时相对电导率大幅升高,其余时间基本在正常水平。表明,在本试验的自然干旱处理时段内,M9T337受伤害的程度较轻,及时复水可有效帮助植株恢复正常生理代谢。
3.3 干旱及复水过程中M9T337苹果砧木根系的变化
本研究发现,短时干旱胁迫未对M9T337根系形态产生明显影响,但胁迫12天后,根系木质化逐渐加重,细根明显减少,根系活力显著降低。表明长时间干旱可明显影响根系的吸收能力,进而影响植株生理代谢。
综合本研究结果,停止浇水18天能对M9T337苹果砧木苗的根系活力和叶片生理代谢产生一定影响,时间越长影响越明显,但及时复水可在一定程度上缓解干旱对其的伤害。
