磷添加对橡胶林土壤N2O、CO2排放及净氮转化速率的影响
2022-05-07刘彦婷尹俊慧王进闯朱四喜王敬国曹文超
刘彦婷,尹俊慧,宋 贺,张 文,王进闯,朱四喜,王敬国,曹文超,5*
(1.贵州民族大学 生态环境工程学院,贵州 贵阳 550025;2.中国农业大学 资源与环境学院,北京 100193;3.安徽农业大学 农学院,安徽 合肥 230036;4.中国热带农业科学院 环境与植物保护研究所/海南省热带生态循环农业重点实验室,海南 海口 571101;5.潍坊科技学院,山东 寿光 262700)
【研究意义】氧化亚氮(N2O)和二氧化碳(CO2)均是重要的温室气体,在全球气候和生态环境的研究中备受关注。热带地区是陆地生态系统中N2O 排放量面积最大和排放速率增长最快的区域,表明该地区是亟需关注的重点区域[1]。同时,热带地区通过土壤微生物与根系呼吸作用对CO2的吸收和排放产生重要影响[2]。在高度风化的热带土壤中,CO2和N2O排放不仅受碳氮底物浓度的影响,也受其他养分有效性含量的限制,特别是磷。磷是构成微生物基因遗传载体、细胞成分合成与代谢及能量转化的必需元素[3]。有效磷含量在热带地区土壤中普遍较低[4],同时土壤Fe3+、Al3+矿物对磷的固定以及通过径流、生物吸收等途径,也会加剧热带酸性土壤的磷限制[5]。土壤磷含量缺乏可影响微生物的生理和活性,并最终影响生态系统包括CO2和N2O 排放在内的碳氮循环过程[6-7]。因此,研究热带地区磷含量如何影响土壤N2O 和CO2产生量的变化,对预测未来气候变化下的温室气体排放具有重要意义。【前人研究进展】磷素作为生态系统初级生产力的重要限制因子[8],其可调控植物生产力和微生物活性[5],改变土壤氮底物有效性并影响氮素转化过程。例如,“氮饱和”森林土壤,磷素亏缺将降低植物和土壤微生物对氮素的吸收[9],导致土壤无机氮浓度增加。磷有效性低会限制土壤微生物活性,并降低土壤氮的矿化[10]。同时,磷有效性低的土壤会造成高量N2O 排放[11],而在热带森林土壤中CO2排放受磷限制的影响程度超过氮,且在低磷土壤中更为明显[12]。因此,外源磷输入对土壤氮素动态变化和N2O 与CO2的排放具有重要影响。【本研究切入点】橡胶树(Hevea brasiliensis)作为热带地区广泛种植的经济树种,不仅保障了国家橡胶战略资源,也在一定程度上发展了偏远地区经济。海南是我国主要的橡胶林种植区,其中橡胶树种植面积占全省植被总面积的1/4[13]。然而,在海南各市县调查橡胶林土壤发现,其有效磷含量(0.49~2.47 mg/kg)均低于橡胶林土壤有效磷养分的正常标准(5~8 mg/kg)[14]。因此,外源磷的输入是保证橡胶树正常生长发育及维系土壤养分平衡的关键。可是,外源磷输入对橡胶林土壤氮素转化和N2O、CO2排放的影响缺乏基础性数据,亟待深入研究。【拟解决的关键问题】本研究利用连续在线监测的自动培养系统,阐明好氧条件下外源磷输入对橡胶林土壤N2O、CO2排放和氮素转化速率的影响,从而为探讨磷素对热带地区橡胶林土壤氮转化调控机制提供理论依据,以期为橡胶林土壤生态系统温室气体减排措施提供数据支撑。
1 材料与方法
1.1 试验材料
供试土壤采自海南省五指山市毛阳镇毛旦村(18°55′N,109°32′E),该地区属热带山区气候,冬暖夏凉,年平均气温22.4 ℃左右,年平均降雨量为1 690 mm。
采样时间为2020 年9 月中旬,选择林龄相同的6 龄橡胶树林,每个样地在15 m×15 m 样方内5 点取样。为了尽量减少橡胶林土壤的空间异质性,设置4次重复。采集0~20 cm土层土壤,充分混合后放入冰盒运回实验室,土壤过2 mm筛,充分混匀后置于4 ℃下保存备用。土壤样品粉粒、砂粒、粘粒的比例分别55.2%、36.8%和7.97%,pH为5.5,NO3-2.47 mg N/kg,NH4+12.1 mg N/kg,全氮1.21 g/kg,有效磷3.41 mg/kg,速效钾80.0 mg/kg,有机质28.95 g/kg。
1.2 试验设计
试验共设置6 个处理:1)CK,空白对照(不添加任何肥料);2)P50,仅添加磷酸二氢钾50 mg P/kg;3)P100,仅添加磷酸二氢钾100 mg P/kg;4)N60,仅添加硫酸铵60 mg N/kg;5)N60P50,添加硫酸铵60 mg N/kg与磷酸二氢钾50 mg P/kg;6)N60P100,添加硫酸铵60 mg N/kg 与磷酸二氢钾100 mg P/kg。每个处理3次重复。称取相当于30 g 烘干土,分别置于120 mL 血清瓶中,根据试验设置处理,用注射器分别向血清瓶内土壤表面喷洒一定浓度的硫酸铵、磷酸二氢钾溶液,以保证其相应氮磷添加量。用去离子水调节土壤质量含水量为25%。利用橡胶塞和铝盖对血清瓶进行密封,采用抽真空-充气系统(北京帅恩科技有限公司)反复用含21%氧气的氦氧混合气冲洗血清瓶5次,最后充入氦氧混合气。将血清瓶置于20 ℃恒温水浴槽中培养,利用注射器平衡血清瓶气压,在Robot自动培养系统上每间隔8 h在线监测血清瓶内N2O和CO2的浓度变化。
Robot自动培养系统主要包括气体分析模块和自动进样模块。其中,气体分析模块主要由气相色谱(Agilent 7890A)组成。气相色谱中包含热导检测器(TCD)、电子捕获检测器(ECD)和火焰离子化检测器(FID)3 个检测器,能够监测N2O(ECD、TCD)、CO(2TCD)和O(2TCD)气体浓度的变化。自动进样模块包括双向旋转的蠕动泵(Gilson Model 222,Gilson,法国)和顶空自动采样器(CTC GC-Pal)。相关Robot自动培养系统的运行模块细节可参见文献Molstad等[15]和McMillan等[16]研究。
1.3 测定项目与方法
1.3.1 土壤理化参数 土壤样品中pH值测定的水土比为2.5∶1(w/v),用pH计测定。全氮含量采用凯氏定氮仪测定。有机碳含量采用重铬酸钾-硫酸溶液氧化滴定法测定[17]。有效磷含量采用0.5 mol/L NaHCO3浸提-钼锑抗吸光光度法测定。速效钾含量采用1.0 mol/L NH4OAc 浸提-火焰光度法测定。土壤水分含量在105 ℃下烘干24 h进行测定。土壤培养之前和之后的NH4+和NO3-含量用1 mol/L KCl溶液浸提后用连续流动分析仪(AA3,Bran Luebbe,Germany)测定。
1.3.2 有关指标的计算方法 顶空体积气体数据计算依据在线监测的气体数值和已知标准气体浓度,计算N2O 和CO2气体每克干土的产生量,计算过程可参见Molstad 等[15]。同时,根据培养前后NO3-和NH4+含量的差值分别计算净氮矿化速率(Nmin)、净氨化速率(Namm)、净硝化速率(Nnit)和硝化率。计算公式如下:
试验数据采用Sigmaplot12.5作图,利用SPSS 20.0分析软件进行因素方差分析。
2 结果与分析
2.1 橡胶林土壤N2O产生速率动态变化及其累积产生量
橡胶林土壤N2O 产生速率动态变化如图1a 所示。整体而言,不同处理土壤N2O 产生速率开始均呈持续上升趋势并于培养32 h后趋近稳定。与其他处理相比,单独添加氮肥N60处理土壤的N2O平均产生速率最大,为0.000 29 nmol N(/g·h)。与N60处理N2O平均产生速率相比,P50与P100处理均显著降低了N2O平均产生速率(P<0.05)。在氮添加基础上,对比N60P50与N60P100来看,增加磷添加量并未显著降低N2O 平均产生速率。从N2O 累积产生量来看(图1b),橡胶林土壤各处理N2O 累积产生量平均范围在0.036~0.047 nmol N/g。单独添加磷的两个处理P50 与P100 土壤N2O 累积产生量并无显著差异,同时二者均显著低于单独氮添加(N60)和氮磷复合添加(N60P50和N60P100)处理(P<0.05)。与N60处理相比,氮磷复合添加并没有显著降低N2O累积产生量。
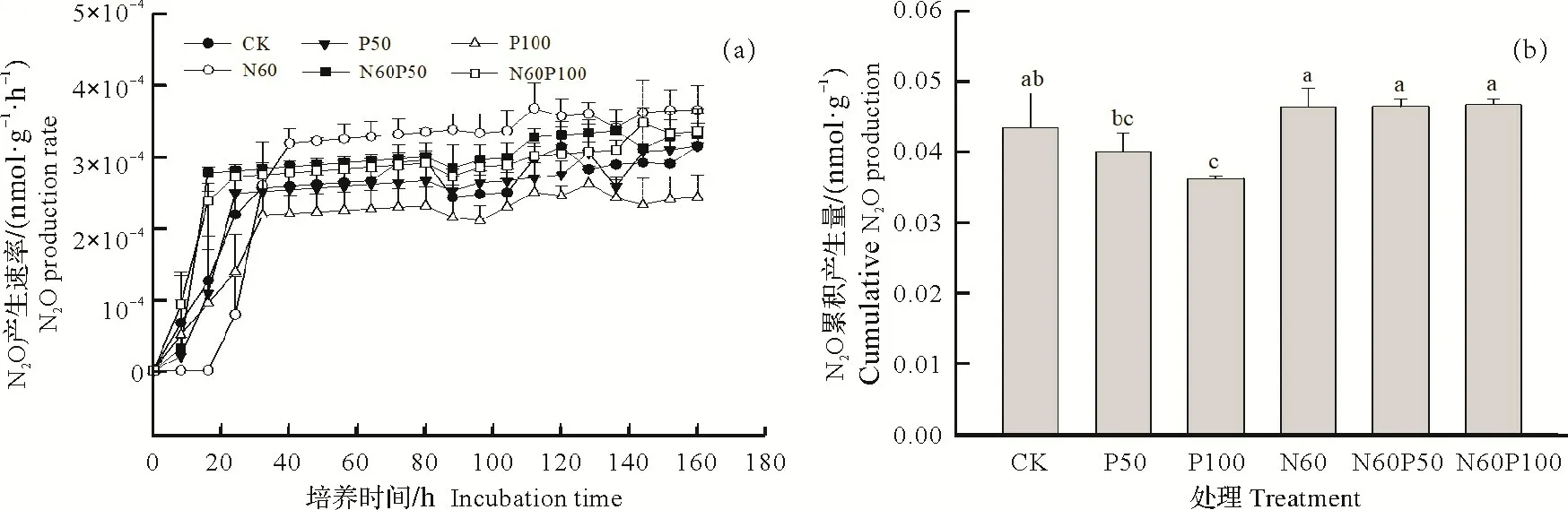
图1 土壤N2O产生速率变化及其累积产生量Fig.1 Dynamic changes of N2O production rate during the incubation period and its cumulative production
2.2 橡胶林土壤CO2产生速率动态变化及其累积产生量
土壤CO2产生速率动态变化如图2a 所示。整体而言,不同处理土壤CO2产生速率在培养第16 时均达到最大值,平均范围为25.5~31.1 nmol C(/g·h),随后呈下降趋势。与CK 相比,单独添加氮肥(N60)显著增加橡胶林土壤CO2平均产生速率(P<0.05)。与N60处理相比,单独添加磷肥(P50、P100)和氮磷复合添加(N60P50和N60P100)处理均显著增加CO2平均产生速率(P<0.05)。与单独添加低磷P50处理相比,增加磷添加量(P100)显著增加土壤CO2平均产生速率(P<0.05)。类似地,在氮添加基础上,增加磷添加量后CO2平均产生速率显著增加(P<0.05)。从CO2累积产生量来看,与未施肥的对照处理相比,单独氮磷添加或氮磷复合添加均显著增加CO2累积产生量(P<0.05,图2b)。与CO2平均产生速率类似,单独磷添加(P50与P100)和氮磷复合(N60P50、N60P100)添加处理橡胶林土壤CO2累积产生量均显著高于单独氮添加N60处理(P<0.05)。与单独磷添加P50处理相比,P100处理橡胶林土壤CO2累积产生量显著增加6.31%。在氮添加基础上,N60P100处理的CO2累积产生量显著高于N60P50处理,为3 849 nmol C/g。
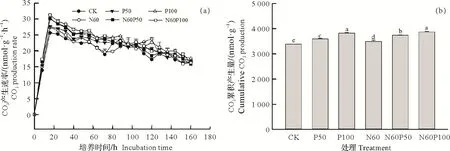
图2 土壤CO2产生速率变化及其累积产生量Fig.2 Dynamic changes of CO2 production rate during the incubation period and its cumulative production
2.3 橡胶林土壤培养前后铵态氮与硝态氮含量变化
与橡胶林土壤初始铵态氮含量(12.1 mg N/kg)相比,各处理土壤铵态氮含量在培养后均显著增加(P<0.05)。单独氮添加N60处理土壤铵态氮含量为51.6 mg N/kg,分别是CK、P50、P100、N60P50、N60P100处理的2.77、3.14、4.00、1.23 和1.38 倍。与对照处理相比,单独磷添加P50 和P100 处理土壤铵态氮含量均显著降低,且二者差异显著(P<0.05)。在氮添加基础上,氮磷复合添加N60P50 与N60P100 处理间铵态氮含量分别为41.8 mg N/kg 和37.5 mg N/kg,磷添加量增加并未显著降低铵态氮含量(P>0.05),但均显著高于对照和单独磷添加处理(P<0.05)。
对于硝态氮含量而言,与初始硝态氮含量(2.47 mg N/kg)相比,各处理的橡胶林土壤硝态氮含量在培养结束后均显著增加(P<0.05),范围为4.87~5.46 mg N/kg。由图3b可知,培养结束后各处理间硝态氮含量并无显著差异。热带地区橡胶林土壤N2O 平均产生速率与铵态氮含量呈极显著正相关关系(P<0.001,图3c),而与土壤硝态氮含量无显著相关性(P>0.05,图3d)。
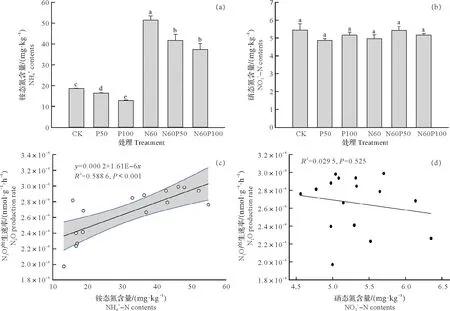
图3 培养后土壤铵态氮、硝态氮含量及其与N2O产生速率的关系Fig.3 The contents of soil ammonium and nitrate and its relationship with N2O production rate at the end of incubation
2.4 橡胶林土壤净氮矿化和净氨化速率
培养期间不同处理土壤净氮矿化速率如图4a 所示。未施肥的对照处理和磷添加(P50、P100)处理橡胶林土壤净氮矿化速率均为正值,说明氮矿化量大于氮固定量,对照CK 和P50 处理土壤净氮矿化速率分别为1.40 和1.03 mg N/(kg·d),二者没有显著差异(P>0.05),但都显著高于P100 处理(0.48 mg N(/kg·d))。橡胶林土壤氮添加(N60)与氮磷添加(N60P50、N60P100)净氮矿化速率分别为-2.57,-3.90,-4.56 mg N/(kg·d),表现为氮的净固定。与单独氮添加N60 处理相比,氮磷复合添加显著增加橡胶林土壤氮的净固定(P<0.05);在氮添加基础上,增加磷添加量并未显著提高橡胶林土壤氮的净固定(P>0.05)。
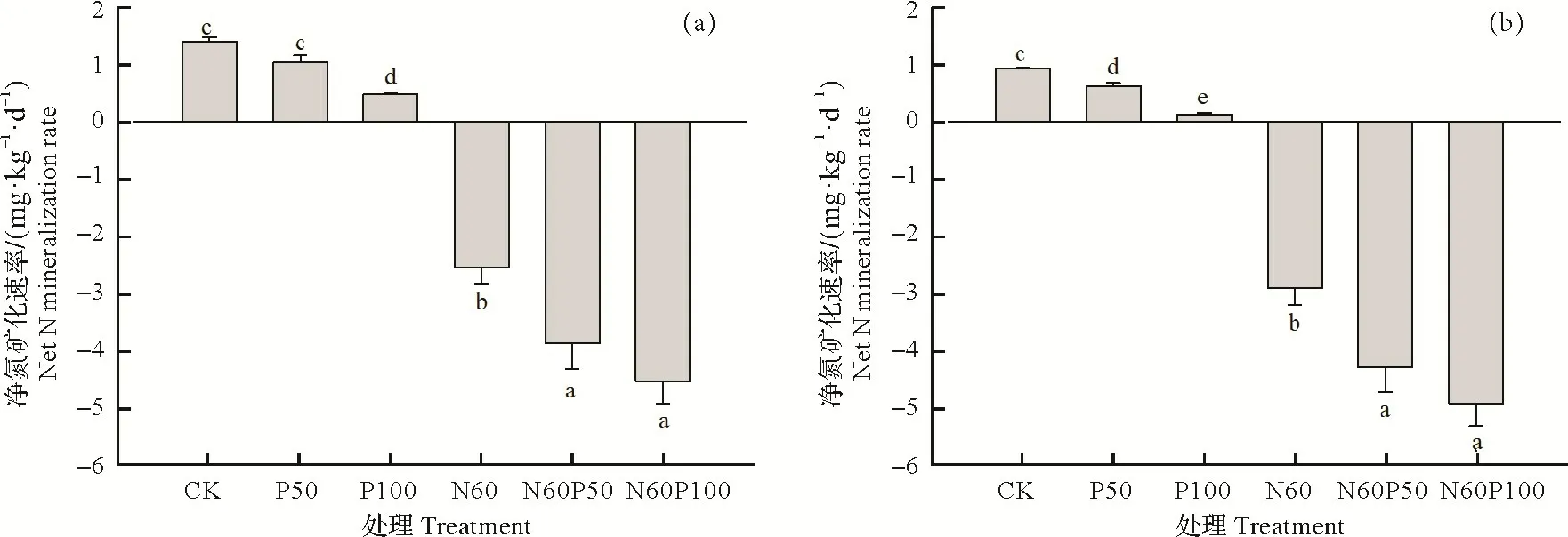
图4 培养结束后土壤净氮矿化速率和净氨化速率Fig.4 The rates of net nitrogen mineralization and net ammonification at the end of incubation
与净氮矿化速率类似,高磷(P100)处理的净氨化速率显著低于对照CK 与P50 处理(P<0.05),同时,P50处理与对照CK差异显著(P<0.05)。对于添加氮肥的所有处理而言,净氨化速率均为负值,表明铵态氮含量在培养之后下降。单独氮添加(N60)处理土壤净氨化速率为-2.93 mg N(/kg·d),显著高于氮磷复合添加处理(P<0.05),其净氨化速率在N60P50和N60P100 中分别显著降低了47.7%和69.0%。此外,在氮添加基础上,增加磷添加量并未显著降低橡胶林土壤的净氨化速率。
2.5 橡胶林土壤净硝化速率与硝化率
不同处理的土壤净硝化速率见图5a。对照CK 处理土壤的净硝化速率为0.38 mg N(/kg·d),分别是P50、P100、N60、N60P50 和N60P100 处理的1.06、1.10、0.98、0.83 和0.98 倍。与CK 处理相比,单独磷添加(P50、P100)与单独氮添加(N60)处理均未显著提高橡胶林土壤净硝化速率。在氮添加基础上,N60P50处理的净硝化速率显著高于N60P100处理(P<0.05)。
不同处理橡胶林土壤的硝化率如图5b 所示。与CK 处理硝化率(23.7%)相比,单独磷添加P50 与P100 处理土壤的硝化率分别为24.5%和26.9%,且与P100 处理差异显著(P<0.05)。整体来看,与CK、P50 和P100 相比,单独氮添加N60 处理和氮磷复合添加N60P50、N60P100 处理土壤硝化率均显著降低,分别为8.82%、11.5%和12.3%。同时。在氮添加基础上,N60P50 与N60P100 处理间硝化率并无显著差异(P>0.05)。

图5 培养结束后土壤净硝化速率和硝化率Fig.5 The net nitrification rates and nitrification ratios in different treatments at the end of incubation
3 讨论
本研究发现,热带海南地区橡胶林土壤N2O 平均产生速率范围为0.000 22~0.000 29 nmol N(/g·h),这与Ishizuka等[18]的研究类似,二者均显著低于西双版纳地区橡胶林土壤的N2O排放[19-20]。原因可能在于橡胶林土壤通常较为贫瘠,再者,低pH 酸性土壤环境导致硝酸盐产生量很低,并不利于硝化与反硝化作用的进行。此外,土壤N2O 排放也受有效磷含量与pH 的影响,这些性质的下降会改变细菌群落并降低细菌物种丰富度[21],进而调控土壤N2O的产生过程及排放。
与单独添加氮肥、氮磷复合添加处理相比,单独磷添加显著降低了热带地区橡胶林土壤N2O 产生速率和累积产生量(P<0.05,图1),这与前人的研究结果类似[22-24]。例如,在我国西南部亚热带地区,磷添加降低次生混交林土壤N2O排放[28],原因可能与磷添加影响植物根系对土壤矿质氮的吸收和微生物对氮素的同化作用有关。与本研究结果不同,Mehnaz 等[25]研究发现磷添加增加土壤N2O 累积排放量,这可能是因为磷添加增加了土壤微生物生物量[26]和反硝化活性[27,31]。由图3c 可见,热带地区橡胶林土壤N2O 平均产生速率与其NH4+含量呈显著正相关关系,而与NO3-含量无显著相关性,意味着该土壤N2O 产生的主要来源可能是硝化作用而非反硝化作用,这与Zhou等[19]在西双版纳橡胶林土壤中的结果相一致。
与对照和单独氮添加处理相比,单独磷添加或氮磷复合添加均显著增加热带地区橡胶林土壤CO2排放,表明橡胶林CO2排放易受土壤有效磷含量的影响。本研究发现,增加有效磷含量能显著促进橡胶林土壤CO2排放。磷被认为是影响热带林地土壤CO2排放的关键限制因子。Cleveland 等[28]研究结果显示,在富含磷的热带森林土壤中磷添加并未提高土壤呼吸速率,而在磷匮乏土壤中显著增加了CO2排放。全球大样本分析表明,热带森林生态系统中磷添加显著增加土壤呼吸速率[29]。一方面,磷添加能够增加根系生物量和细根量[9],进而增加自养呼吸;同时,磷添加增加热带森林土壤中的微生物生物量碳含量[30],促进土壤有机物的分解[31]和异养呼吸作用。此外,磷添加可能通过诱导微生物对氮的需求或刺激SOM降解菌群的生长来加快SOM分解[27]。本研究中,橡胶林土壤CO2排放在各处理中均呈先增加后下降的趋势,这与热带森林土壤中的研究结果相一致[12],意味着土壤呼吸还受碳底物含量的限制。例如,热带森林土壤在高碳浓度下(1 mg C/g)添加磷会增加CO2排放,而在低碳浓度下(0.1 mg C/g)无显著差别[32]。与对照处理相比,单独氮添加后橡胶林土壤CO2排放显著增加,表明氮添加促进土壤CO2的排放。这与Tian 等[33]研究的结果不同,其发现氮添加并未影响表层土壤CO2的排放。结果的不一致可能与土壤碳底物有效性的差别有关。同时,氮添加能够提高土壤轻组有机碳和新鲜凋落物的降解速率,而对重组有机碳和腐殖化有机质的分解有抑制作用[34]。
与对照处理相比,单独磷添加显著降低了橡胶林土壤净氮矿化与净氨化速率(图4),表明磷添加可能潜在增加橡胶林土壤氮的固定。同时,与单独氮添加处理相比,氮磷复合添加也显著增加土壤氮的固定,这与Cheng 等[7]的研究结果相一致。这可能是因为磷添加后增加土壤微生物活性,导致微生物从土壤中获取氮含量增加,进而增加NH4+的固定速率[35]。类似地,Mehnaz等[25]研究发现,磷添加并未提高土壤总氮矿化速率,原因在于磷添加后更多的土壤氮被微生物同化,而低分子量有机氮(如氨基酸)也被固定,阻止了其进一步矿化为NH4+的能力。此外,磷添加对土壤氮的转化可能存在不确定性。例如,热带森林土壤中磷添加会增加微生物生物量并改变其群落组成,但在4 年施肥后这种影响便消失[36],意味着热带地区土壤磷添加对氮转化的影响可能存在短期效应。从植物根系与土壤微生物竞争养分的角度来看,土壤细菌活性主要受碳限制[37],而植物生长常受氮、磷养分有效性的限制[38]。对于缺磷植物而言,磷的吸收速率也会限制它吸收非限制养分的能力[39],因而,在磷缺乏而氮饱和的生态系统中,土壤微生物对氮素的同化固定要高于植物对氮素的吸收,而外源磷添加可能促进氮饱和土壤氮矿化。可见,土壤氮转化对磷添加的响应较为复杂,这与土壤类型、氮磷含量、施磷时间以及其他因素有关。
4 结论
综上,与单独氮添加相比,热带地区橡胶林土壤单独磷添加后显著降低N2O 排放并增加土壤呼吸速率。单独磷添加对N2O 和CO2两种温室气体排放的影响存在某种程度上的“此消彼长”效应。在相同氮添加基础上,增加磷输入并未显著增加N2O 的产生,而显著增加了CO2产生量。磷添加显著降低土壤NH4+含量,而对NO3-含量无显著影响。橡胶林土壤N2O 平均产生速率与NH4+含量呈极显著正相关关系而与含量无关。与对照相比,单独磷添加显著降低橡胶林土壤净氨化速率。在氮添加基础上,磷的输入显著增加橡胶林土壤氮的固定。与单独氮添加相比,单独磷添加显著增加橡胶林土壤硝化率而对净硝化速率无影响。考虑到热带海南地区橡胶林土壤中普遍缺磷,结合本研究结果,表明在橡胶林土壤中合理施用磷肥可能是实现温室气体N2O 减排的重要措施,而在此过程中,磷肥施用增加温室气体CO2排放需要引起重视。
致谢:中国热带农业科学院基本科研业务费专项(1630042020004)同时对本研究给予了资助,谨致谢意!
