贵州麻黄洞空气CO2与上覆土壤空气CO2相关性研究
2022-04-28石亮星周忠发丁圣君范宝祥汤云涛
石亮星, 周忠发*, 丁圣君, 董 慧, 范宝祥, 安 丹, 汤云涛
贵州麻黄洞空气CO2与上覆土壤空气CO2相关性研究
石亮星1, 2, 周忠发1, 2*, 丁圣君1, 2, 董 慧1, 2, 范宝祥1, 2, 安 丹1, 2, 汤云涛1, 2
(1. 贵州师范大学 喀斯特研究院/地理与环境科学学院, 贵州 贵阳 550001; 2. 贵州省喀斯特山地生态环境国家重点实验室培育基地, 贵州 贵阳 550001)
CO2作为岩溶作用的驱动力, 在岩溶作用中起着关键作用。岩溶区特有的地上地下二元结构表明, 洞穴系统作为地下空间的窗口, 对其CO2及δ13CCO2研究是十分必要的。本研究对贵州绥阳麻黄洞上覆土壤空气CO2、洞穴内部和外部大气参数以及CO2浓度和δ13CCO2进行了为期12个月的监测, 监测结果表明: ①麻黄洞洞穴空气和上覆土壤空气CO2与δ13CCO2均呈现出明显的时空变化规律, 表现出雨季CO2浓度高、δ13CCO2偏轻, 旱季CO2浓度低、δ13CCO2偏重的特征。②土壤CO2是内源性CO2以及大气CO2经平流渗透综合作用的结果, 在温度、湿度双向调控下土壤微生物作用存在差异进而导致CO2浓度和δ13CCO2存在时空差异, 洞穴CO2浓度和δ13CCO2主要受上覆土壤CO2及其通风效应的影响。③麻黄洞CO2主要来源于上覆土壤CO2以及外部大气, 在通风干预下洞穴内部产生稀释效应或者集聚效应。探究土–气CO2和δ13CCO2变化特征, 探明洞穴CO2来源, 不仅有利于了解洞穴系统碳循环机制, 而且对全球碳循环中“遗漏碳汇”研究具有积极意义。
岩溶洞穴; CO2来源; 稳定碳同位素
0 引 言
近年来全球气候变化受到广泛关注, 碳作为气候变化的重要驱动力, 研究全球碳循环对于研究全球气候变暖的根源、预测极端天气具有积极意义(Serrano-Ortiz et al., 2010; 吴泽燕等, 2019; 曾广能等, 2019)。洞穴作为地下岩溶系统特有的窗口, 对岩溶碳循环过程产生重要的源汇效应(汪炎林等, 2020), 且CO2主导岩溶作用发生(Lachniet, 2009)、洞穴环境质量(童晓宁等, 2013; 张萍等, 2017; 梁明强等, 2019)以及洞穴次生沉积物再沉积和溶蚀过程(James et al., 2015)。作为岩溶碳循环的重要一环, 研究岩溶洞穴土–气响应不仅有利于研究洞穴系统碳迁移机制, 还有利于理解岩溶动力系统三相过程, 对探究“遗漏碳汇”具有积极意义。
作为地球表面最大的碳库, 土壤在岩溶碳循环中发挥了重要作用(田娜等, 2010)。由于植物根系呼吸作用以及微生物对有机质的分解作用, 土壤中含有大量的CO2。土壤中CO2主要以两种方式参与陆地生态系统碳循环: ①通过土壤呼吸向大气中排放CO2, 为大气CO2的源(任京辰等, 2006); ②土壤中CO2为岩溶作用提供重要动力, CO2以HCO3−的形式进入岩溶水中, 成为大气CO2的汇(赵瑞一等, 2015)。岩溶水携带上覆CO2下渗至洞穴内部, 成为洞穴CO2的源, 为洞穴系统碳循环的一部分。除了土壤作用, 通风效应也会影响洞穴内部CO2变化。与外部大气相比, 由于岩石的低导热性, 洞穴内部温度通常比较稳定, 因此会产生内外温度差, 从而引发通风, 促进洞内外空气交换过程(De Freitas et al., 1982; Sánchez-Cañete et al., 2013; Wang et al., 2019)。在通风效应驱动下, 洞内外发生气流交换, 使洞穴内部CO2发生积累或者稀释, 进而影响洞穴系统碳循环。
近年来, 岩溶洞穴系统土–气研究取得了丰硕的成果, 如Fairchild et al. (2006)基于水、岩、土、气等方面研究了垂直碳迁移路径, 得出洞穴内部主要受上覆土壤CO2影响, 认为由于上覆基岩的不可知性, 碳迁移路径存在不可知性; 殷超等 (2017)基于洞穴上覆、洞穴滴水以及洞内CO2变化研究得出上覆土壤CO2是洞穴CO2主要来源, 且存在1个月的“滞后效应”。随着CO2与水化学特征响应研究的不断深入, Cao et al. (2020)对雪玉洞不同时间尺度上覆土壤CO2对滴水化学响应关系进行了研究, 印证了洞穴上覆CO2会影响洞内CO2变化。关于洞穴CO2来源研究, 也逐渐经历了由定性向定量不断精确的过程。如Rutiligge et al. (2014)通过对洞穴滴水监测数据, 结合数理统计方法, 得出洞内CO2变化是上覆土壤环境与外界大气综合作用的结果; Mattey et al. (2016)基于稳定同位素的研究表明, 在直布罗陀地区, 土壤CO2并非洞穴CO2主要来源, 洞穴CO2主要是受包气带中“地面空气”的影响; 刘子琦和李开萍 (2018)基于洞穴对石漠化响应研究表明土壤CO2并非洞穴CO2的唯一来源; 汪炎林等 (2020)基于不同季节洞穴系统各层CO2变化特征研究得出土壤和表层岩溶带是洞穴CO2的两个来源。由于稳定碳同位素能够指示碳元素在岩溶动力系统中的迁移变化, 反映岩溶过程中碳的变化过程、CO2的具体来源以及定量分析, 稳定碳同位素技术在岩溶碳循环研究中发挥了重要作用, 对于岩溶CO2研究具有指示意义(Mandić et al., 2013; Krajnc et al., 2017)。
前人对岩溶洞穴CO2研究存在以下不足: ①对洞穴内部CO2影响因素量化不足, 缺乏论证; ②论证土–气响应关系时, 往往忽略通风效应对洞穴CO2变化的影响, 导致结果存在偏差。本文通过对麻黄洞内部及其上覆土壤CO2浓度和δ13CCO2进行为期一年的监测, 探讨洞穴空气、上覆土壤空气CO2的时空变化规律和响应关系, 从而探究喀斯特洞穴CO2的来源和迁移机制, 为岩溶洞穴系统碳循环研究提供理论依据。
1 研究区概况
研究区位于贵州省绥阳县温泉镇境内, 距遵义市区120 km, 属芙蓉江一级支流双河洞小流域。地质构造上, 洞系属于大娄山山脉的东北支, 由于受不同方向区域构造应力作用, 形成NE、NW及SN向褶皱断裂带。其中寒武系中上统娄山关组(Є2-3)地层中喀斯特发育, 地貌类型丰富, 为洞穴主要发育地层, 洞区岩性主要以白云岩和白云质灰岩为主(刘平, 2008)。研究区气候属中亚热带季风气候, 并伴有亚热带山地季风气候的特征, 雨热同期。土壤以黄壤、石灰土为主, 植被以亚热带常绿阔叶林和阔叶落叶混交林为主(李坡等, 2008)。
麻黄洞是双河洞洞穴系统的一级支洞, 总洞道海拔在678~720 m之间, 长约1100 m, 洞口海拔720 m,高约32 m, 宽约15 m, 洞道为南北走向, 平直且多洞厅, 洞穴中部发育地下河, 洞内多滴水点, 洞穴上覆植被类型为有林地、撂荒地以及旱地。
2 数据来源及研究方法
如图1所示, 结合麻黄洞洞道及其上覆地表覆被, 选取6个洞内监测点(MH1#~MH6#; 表1)、3个上覆土壤空气监测点(S1#~S3#; 表2)以及1个洞外监测点进行逐月监测。于2018年10月至2019年9月对麻黄洞洞内外空气环境及其上覆土壤空气CO2进行为期12个月的监测, 并采集气体样本。为了减少样本和实地监测误差, 采样和监测固定在每月中旬进行。洞内使用美国Telaire-7001型便携式红外CO2仪对CO2浓度(体积分数)进行逐点测量, 仪器分辨率为1×10−6, 测量范围为0~10000×10−6, 测量精度为±50×10−6。为了保证测量的精确性, 测量时仪器与人保持大于2.5 m的距离。洞穴内外空气中的风速、温度、相对湿度、气压、海拔则使用美国Kestrel-4500型便携式气象站进行实时监测, 仪器分辨率分别0.1 m/s、0.1 ℃、0.1%、0.1 kPa、1 m, 精度分别为±3%、±1.0 ℃、±3%、±0.15 kPa(25 ℃)、±15 m。土壤空气采用JA901泵吸式二氧化碳检测仪测定, 测量范围为1×10−6~50000×10−6, 分辨率为1×10−6, 检测精度≤300×10−6, 线性误差≤100×10−6, 响应时间≤20 s。现场采用DEV31-1型气体采样袋采集大气空气、土壤空气和洞穴空气, 用于室内稳定碳同位素测定, 室内实验分析在中国科学院地球化学研究所环境地球化学国家重点实验室测定完成。野外采集大气和洞穴气体δ13CCO2使用美国Finnigan MATG公司生产的MAT252气体同位素质谱仪测定, 每测4次样品测定一个平行样品, 测定结果用国际标准VPDB(Vienna Pee Dee Belemnite)校准, 测试精度≤0.01‰。气象数据来源于双河洞国家地质公园最近的桐梓县气象观测站, 指标包括气温、降水量等信息。
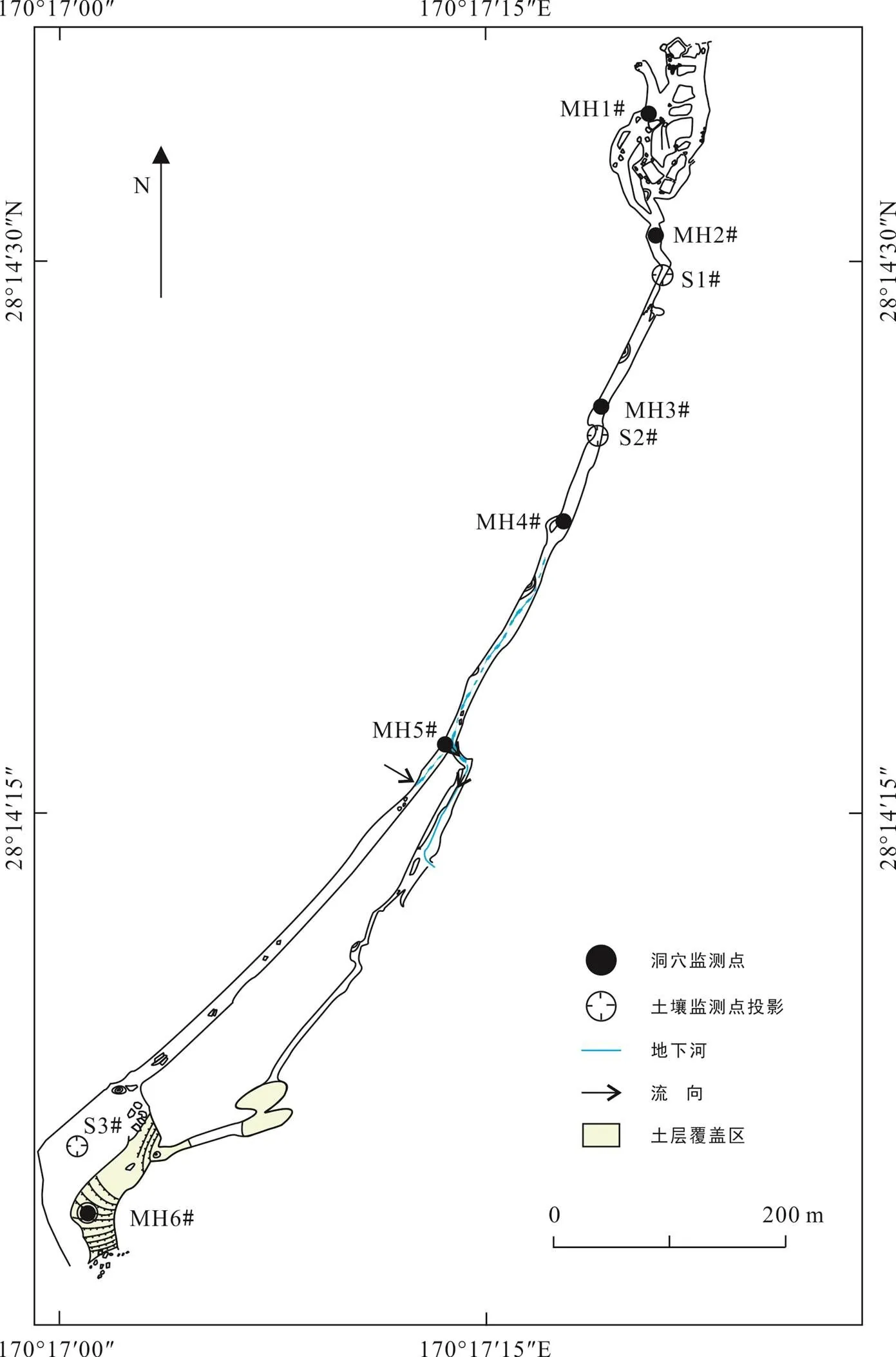
图1 麻黄洞洞道及监测点概况
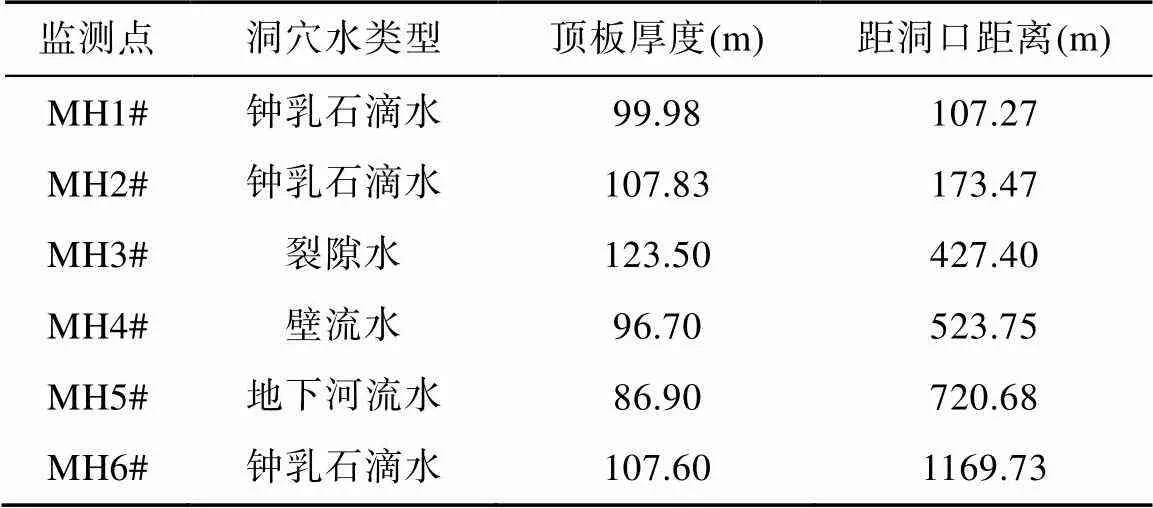
表1 麻黄洞洞穴各监测点基本参数统计表
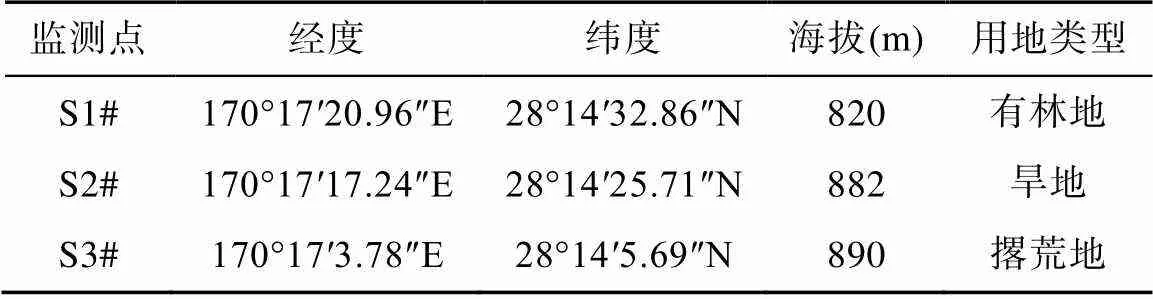
表2 麻黄洞上覆土壤监测点基本参数统计表
3 结 果
3.1 洞内外温度、降水的变化特征
麻黄洞洞内外环境呈现出明显的时空变化(图2)。受亚热带季风气候控制, 研究区温度、降水均呈现出明显的季节变化。研究区温度变化范围为4.4~26.1 ℃,平均温度为15.3 ℃; 降水总量为1289.7 mm, 集中在4~9月, 在此期间月均降水为204 mm, 降水量达到了全年的80%, 其余月份月均降水仅为38 mm。因此, 按照气候特征及降水分布状况, 本研究中将4~9月划分为雨季, 10月至次年3月划分为旱季。由于岩石的低导热性, 岩层较厚且洞穴较为封闭, 因此相比外部大气, 洞穴内部环境相对稳定, 内部温度变化较小。洞穴内温度变化范围为5.6~19.6 ℃, 平均值为13.5 ℃, 其变化趋势与外部大气保持一致。位于洞口的MH1#温度变化范围为6.4~18.3 ℃, 位于洞穴中部的MH3#范围则为9.4~16 ℃, 洞穴深处的MH6#范围为12.3~19.6 ℃, 总体呈现出随离洞口距离增加而逐渐趋于稳定的特征。

图2 洞内外气象参数变化特征
3.2 土壤、洞穴空气CO2浓度及δ13CCO2变化特征
如图3所示, 上覆各监测点土壤CO2浓度(体积分数)存在明显的时空特征。空间上, 各土壤监测点CO2浓度变化幅度较大, 变化范围在1494×10−6~ 40684×10−6之间, 各土壤监测点CO2浓度随着深度增加而增大。S1#、S2#、S3#土壤CO2浓度变化范围分别为4105×10−6~40684×10−6、1494×10−6~32083×10−6以及6480×10−6~31329×10−6, 平均值分别为17735×10−6、13857×10−6以及16255×10−6。时间上, 各监测点总体上呈现出雨季高、旱季低的特征。在雨季, 部分监测点表层CO2浓度先增加后降低。由于各监测点植被覆盖存在差异, 其CO2浓度也存在差异, 各土壤监测点平均CO2浓度大小依次为S1#>S3#>S2#。
洞穴CO2浓度总体变化范围在477×10−6~898×10−6之间, 平均值为639×10−6。最低值出现在2019年2月的MH6#, 最高值出现在2019年4月的MH3#, 但其总体趋势表现为随着洞口距离的增加变化趋于稳定(图2)。时间上, 与上覆土壤CO2浓度变化趋势一致, 麻黄洞内空气CO2浓度表现出明显的雨季高、旱季低的周期性特征。雨季, 各监测点CO2浓度变化范围为573×10−6~898×10−6, 平均值为821×10−6; 旱季变化范围为477×10−6~614×10−6, 平均值为534×10−6。
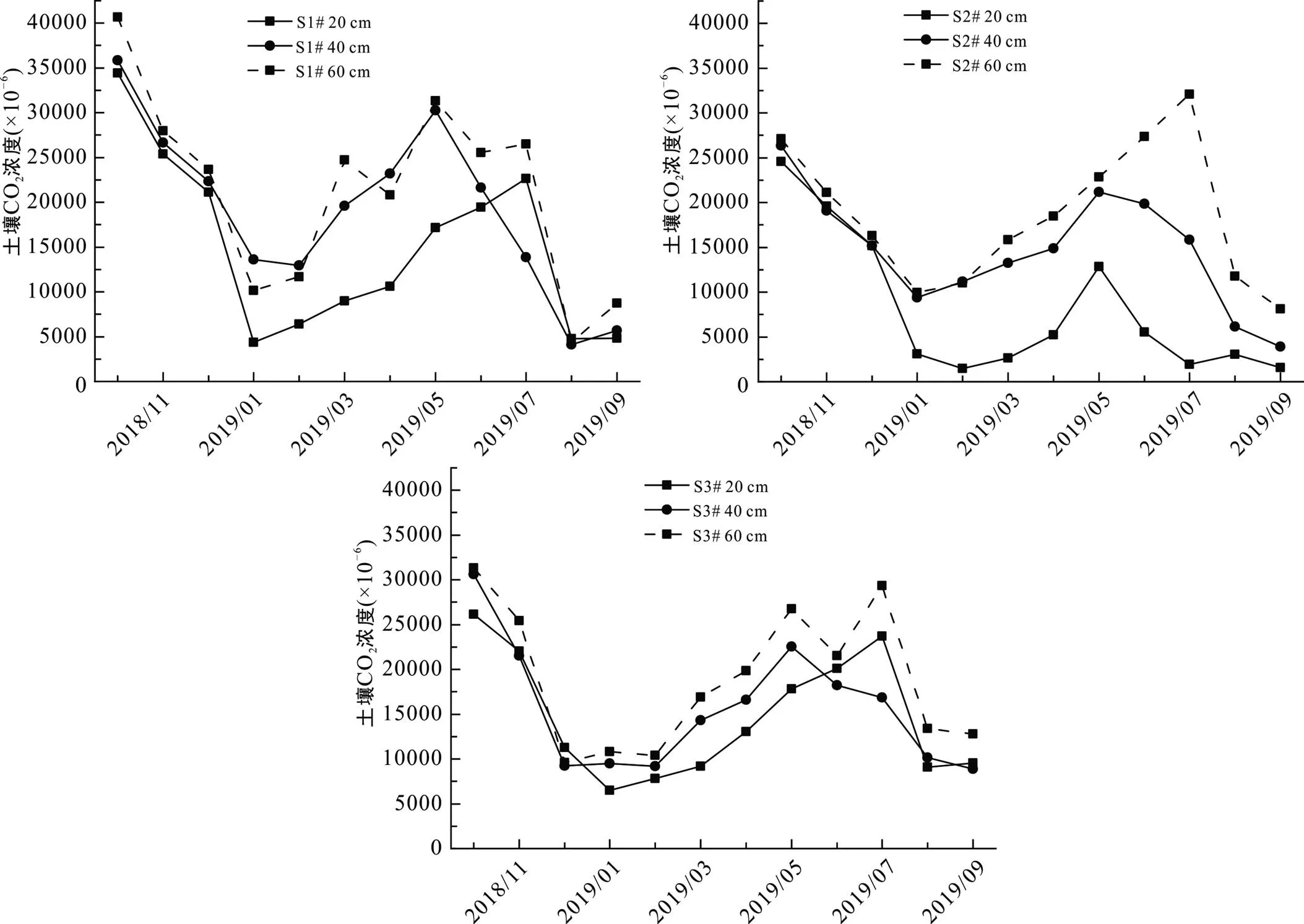
图3 土壤CO2浓度变化特征
如表3所示, 麻黄洞上覆监测点土壤空气δ13CCO2值存在时空差异。上覆土壤空气δ13CCO2变化范围为−26.7‰ ~ −20.0‰。其中, S1#的δ13CCO2变化范围为−26.7‰ ~ −22.2‰, 平均值为−24.6‰; S2#的δ13CCO2变化范围为−26.1‰ ~ −20.0‰, 平均值为−24.7‰; S3#的δ13CCO2变化范围为−26.4‰ ~ −22.1‰, 平均值为−24.7‰。时间上, 各土壤监测点δ13CCO2值呈现出雨季偏轻, 旱季偏重的特征。
洞穴空气CO2主要由两部分组成: 碳同位素偏轻的生物源CO2以及碳同位素偏重的大气CO2, 前人研究表明其来源主要是通风效应(Mandić et al., 2013)、滴水脱气作用(Rutlidge et al., 2014)以及包气带传输扩散(Cao et al., 2020)。麻黄洞两个空气点δ13CCO2值变化范围为−20.1‰ ~ −8.0‰, 平均值为−13.4‰。MH2#、MH3#的δ13CCO2范围分别为: −16.7‰ ~ −8.3‰和−20.1‰ ~ −8.0‰。时间上, 与上覆土壤空气一致, 洞穴空气δ13CCO2同样呈现出雨季偏轻, 旱季偏重的季节特征。
相比洞穴空气与土壤空气, 大气δ13CCO2总体偏重, 变化范围为−12.0‰ ~ −9.5‰, 平均值为−11.1‰, 与大气CO2浓度一致, 全年较稳定。
4 讨 论
4.1 洞穴上覆土壤空气CO2浓度和δ13CCO2变化的原因
如表2所示, S1#为有林地, 植物根系发达, 腐殖质层较厚, 蓄水能力强, 土壤微生物作用强, CO2浓度高, 平均值为17735×10−6; S2#为旱地植物根系以及微生物相对较少, CO2浓度相对较低, 平均值为13857×10−6; S3#为撂荒地, 根系发育, 生物作用较强, CO2浓度较高, 平均值为16255×10−6。土地利用类型不同, 其生物作用、理化性质存在差异使得各监测点CO2浓度呈现出空间差异。研究区属亚热带季风气候, 雨热同期, 雨季在温度与降水的双重调控下, 植物根系呼吸作用以及微生物作用达到一年间顶峰, 各监测点土壤CO2浓度均达到全年的最高值; 旱季, 由于降水较少, 温度较低, 生物作用受到抑制, 土壤CO2浓度出现低值。因而土壤CO2呈现出雨季高, 旱季低的特征。

表3 监测点δ13CCO2值(‰)
综合而言, 土壤CO2来源主要涉及三个方面: ①植物光合作用固定大气的碳, 残体以有机碳的形式进入土壤中, 与其他有机残体一起被微生物分解, 产生CO2(Breecker et al., 2012); ②植物呼吸作用(Buczko et al., 2015); ③大气CO2(Breecker et al., 2012)。CO2来源和植被类型不同, δ13C值会存在差异。通常将生物作用产生的CO2称为内源性CO2, 麻黄洞上覆植被以C3植物为主, 其δ13C值为−28‰ ~ −26‰。除CO2来源影响δ13CCO2之外, 土壤δ13CCO2变化还受两个过程影响: ①植物根系呼吸作用产生的动力学分馏, 通常会优先分出偏轻的12C; ②土壤空气与大气的平流渗透, 土壤中CO2是大气与表层岩溶带等深部岩溶中CO2扩散传输与分馏的最终结果, 与内源性CO2存在差异。在土壤孔隙度、土壤湿度、温度等物理参数的驱动下, 旱季雨季混合程度存在差异。
监测数据表明, 麻黄洞上覆土壤空气δ13CCO2呈现出雨季偏轻、旱季偏重的特征。雨季, 生物作用强, 土壤孔隙度低, 土壤空气中主要为内源性CO2, 其δ13C值偏轻。另外, 由于基岩有较好的溶蚀速率, 在降水的淋溶作用下, 生成的HCO3−随土壤流失, 使偏重的13C向下转移, 形成了较轻的δ13CCO2, 范围为−22.2‰ ~ −26.4‰。旱季, 土壤湿度和温度均较低, 微生物作用受到抑制, 土壤孔隙度大, 从而使土壤空气和大气之间的交换频率增加, 使两者充分混合, 使得δ13CCO2偏重, 范围为−20‰ ~ −25.1‰。此外, 由于8~9月在高温驱动下, 土壤含水率降低、土壤孔隙增大, 同样加剧了土–气交换, 尽管降水丰沛, 其土壤δ13CCO2呈现出季节性低值。
综上所述, 土壤空气δ13CCO2主要受植物根系呼吸作用、植物根系对土壤有机物分解以及微生物分解有机质影响, 在温度与湿度驱动下呈现出的季节性特征。同时, 土壤通过与表层岩溶带和大气间的对流运动和扩散运动进行气体交换, 外部环境变化对对流运动产生影响, 进而影响土壤空气δ13CCO2变化。
4.2 洞穴空气CO2浓度和δ13CCO2变化的原因
麻黄洞洞内CO2浓度呈现出雨季高旱季低的特征, 雨季CO2浓度为573×10−6~898×10−6, 平均值为821×10−6; 旱季变化范围为477×10−6~614×10−6, 平均值为534×10−6。与CO2浓度相反, δ13CCO2呈现出雨季偏轻旱季偏重的特征, 变化范围为−20.1‰ ~ −8.0‰。
雨季, 在温度与降水的双向调控下, 上覆土壤中生物作用较强, 土壤中形成较高浓度生物源CO2, 经过裂隙孔隙、滴水脱气等作用进入洞内。此时降水量和下渗量均大, 渗流携带更多的土壤CO2进入洞穴; 大气降水下渗时会形成“活塞效应”, 将更多的土壤空气压缩至洞内, 洞内形成相对较高CO2浓度。此外, 雨水在快速下渗作用下进入洞穴发生脱气, 在脱气过程中也会优先脱出较轻的12C。在较轻的土壤δ13CCO2的影响下, 洞内偏轻δ13CCO2积累, δ13CCO2呈现出低值。旱季, 温度较低, 降水较少, 上覆土壤生物作用减弱, 干旱改变土壤理化性质, 土气交换量增加, 土壤中CO2浓度较低使得土壤δ13CCO2偏重。此时, 土壤CO2输送较少且洞穴下渗作用弱, 随滴水进入洞穴CO2较少, 洞内CO2浓度降低。降水在基岩部分停留时间增加, 促进基岩溶蚀作用, 渗流携带更多基岩中的老碳, 使得滴水δ13CCO2偏重, 进入洞穴后发生脱气使洞穴空气δ13CCO2偏重。
通风效应对洞穴CO2浓度和δ13CCO2也会产生重要影响。受通风驱动, 洞穴环境存在周期性变化。通风状态由地貌决定, 由天气变化调节, 因此每个洞穴的通风状态都存在差异。通风模式主要受洞内外密度差驱动。由于麻黄洞CO2浓度低且不存在人为干扰, 可以用洞穴内外部温差来估算洞穴空气密度, 判断洞穴的空气浮力, 进而表明通风模式。计算公式为(Vieten et al., 2016):

其中: 当Δ时, 浮力为正, 表明洞外空气密度高于洞内, 气流从洞外流向洞内; 当Δ时, 浮力为负, 表明洞内空气密度高于洞外, 意味着气流从洞内流向洞外, 洞内外气流交换受到抑制。
计算结果如图4所示。结果表明: 雨季, 洞内虚拟温度低于洞外, 洞穴空气密度高于外部空气, 外部大气自洞口进入洞内, 逐渐冷却下沉, 洞内外气流交换受限, 上覆高浓度CO2在洞内发生积累效应, 形成较高浓度CO2, 且δ13CCO2偏轻; 旱季, 洞内温度高于洞外, 洞外空气自动口向内长驱直入, 外部低浓度CO2与洞内空气发生混合, CO2浓度降低, 洞内CO2与大气偏重CO2发生混合, 使得δ13CCO2偏重。前人研究表明, 距离洞口越近通风越强(James et al., 2015; Treble et al., 2015), 使洞穴CO2浓度出现空间差异。基于同位素平衡理论, 由于MH2#更接近洞口, 通风更强, 与大气交换更频繁, 在偏重大气δ13CCO2的参与下, MH2#与MH3#洞穴空气δ13CCO2存在差异。
4.3 麻黄洞洞内CO2来源
洞穴CO2来源主要为: 上覆CO2通过裂隙和岩溶管道进入洞穴(Fichez, 1991; 任坤等, 2016)、土壤CO2被下渗流或地下水携带进入洞穴发生脱气效应释放CO2(Baldini et al., 2006; 张结等, 2018)以及通风效应带来的外部CO2(Shindoh et al., 2017), 涉及大气、地上、地下三个端元之间的对流、扩散以及脱气作用。
为了量化各因子对于洞穴CO2的贡献率, 基于主成分分析法定量分析土壤CO2浓度以及其他参数对洞穴CO2的影响。所选样本显著性检验的值小于0.05, 表明被解释变量与解释变量间的线性相关性显著, 建立的模型是合理的, 结果如表4所示。
由表4可知, 结果提取了土壤CO2浓度和洞穴温度两个主成分, 二者为影响洞穴CO2浓度变化的主要因素。而温度的变化往往直接影响洞穴通风, 由此可知洞穴CO2浓度主要受土壤CO2浓度以及通风效应的控制。
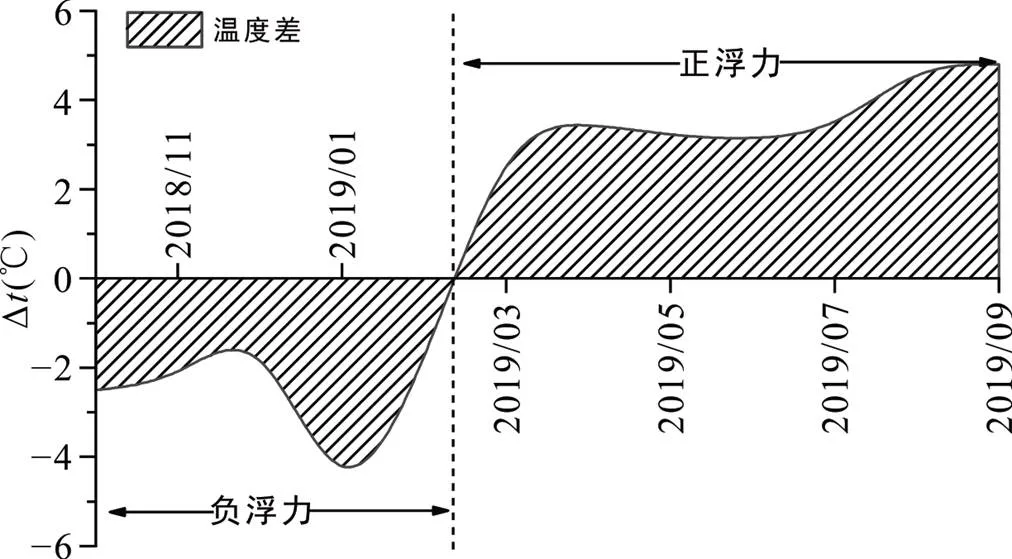
图4 洞内外温度差变化趋势
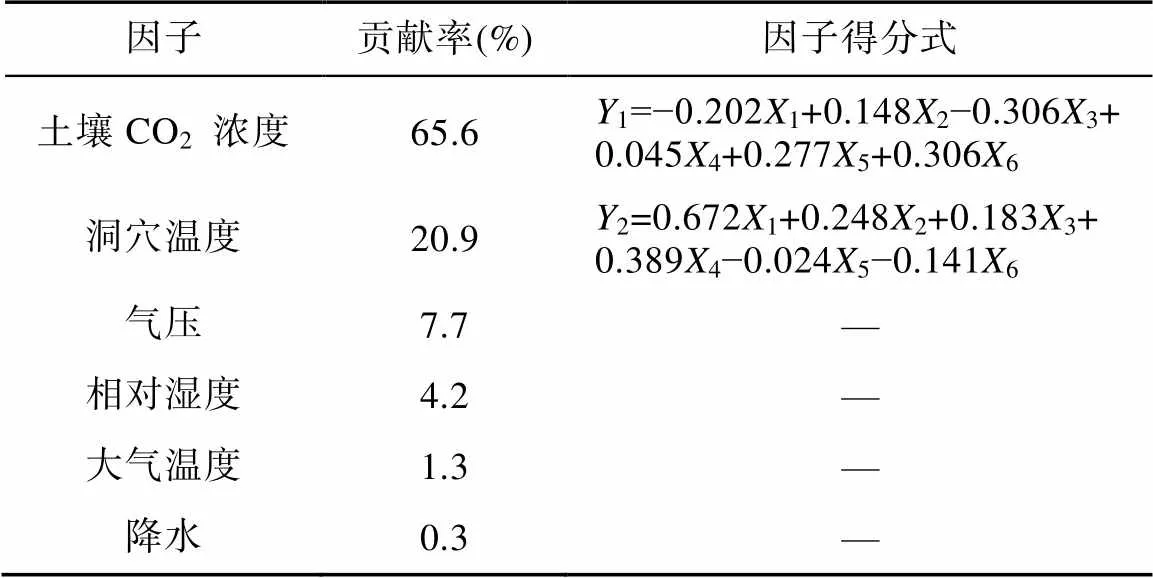
表4 基于主成分分析各因子对洞穴CO2贡献率
麻黄洞上覆土壤空气CO2浓度与洞穴CO2浓度存在旱季低雨季高的变化趋势, 基于主成分分析表明, 土壤CO2浓度是洞穴CO2浓度变化的主要影响因子, 印证了上覆土壤是洞内CO2的主要来源之一, 且存在明显的季节特征。麻黄洞空气δ13CCO2偏重且远重于上覆土壤δ13CCO2, 介于上覆土壤空气δ13CCO2以及大气δ13CCO2之间, 且随大气和土壤空气δ13CCO2变化而变化, 间接印证了麻黄洞空气CO2与上覆土壤空气CO2和大气CO2混合存在响应关系。
为了进一步探究洞穴CO2来源, 利用Keeling plot模型(Nickerson and Risk, 2006)建立洞穴1/(CO2)与δ13CCO2的线性关系, 结果显示二者间存在较好的线性关系(图5), 说明洞穴CO2存在稳定的补给来源, 其截距值与上覆土壤δ13CCO2基本接近, 说明土壤CO2是洞穴CO2的主要补给, 但基于主成分分析可知, 洞穴CO2会受到通风效应的干预。因此, 洞穴CO2主要存在外部大气和上覆土壤两个端元补给。
相较于旱季, 雨季洞穴空气δ13CCO2总体偏轻, 表明4~9月洞穴上覆土壤CO2是洞穴内部CO2变化的主要贡献因子。雨季, 由于温度升高和降水增加, 土壤微生物作用强烈, 产生大量的高浓度的土壤CO2并溶于水中, 沿着水流路径(岩溶裂隙和管道)下渗进入洞穴。然而, 在旱季, 因温度减低且降水减少, 土壤微生物作用减弱, 由生物作用产生的土壤CO2相应减少, 导致土壤CO2同位素偏重。此外, 降水减少导致岩溶裂隙和管道中的水处于非饱和状态, 水流在岩溶裂隙和管道中的滞留时间增加, 水岩相互作用显著, 使碳酸盐岩不断溶蚀, 在进入洞穴时通过瞬时脱气作用而贡献CO2。此时的洞穴空气δ13CCO2可能继承了基岩属性, 使得洞穴δ13CCO2值增加。由于麻黄洞是单洞道洞穴, 在旱季, 洞外气温较低, 洞内外气流交换作用显著, 洞外冷重的空气进入洞穴不断稀释或交换洞穴空气CO2, 导致洞穴δ13CCO2偏重。

图5 洞穴1/φ(CO2)与δ13CCO2的相关图
综上, 麻黄洞CO2主要受土壤CO2与外部大气综合作用, 且主要受土壤控制。雨季, 上覆生物作用强, 产生大量偏轻生物源CO2, 随降水下渗作用进入洞穴内部, 经脱气作用, 脱出较轻CO2进入洞穴, 由于洞穴处于限制性通风阶段, 洞内外空气交换受限, 此时洞穴内部CO2浓度较高, δ13CCO2出现低值。旱季, 上覆生物作用较弱, 土壤δ13CCO2偏重, 下渗水在基岩停留时间较长, 形成偏重δ13CCO2, 脱气进入洞穴后, 此时洞穴为积极性通风模式, 洞内外气流交换, 在大气作用下, δ13CCO2在混合作用调控下出现高值。
5 结 论
(1) 麻黄洞空气CO2与上覆土壤空气CO2存在明显的季节性特征。总体表现为CO2浓度雨季>旱季, δ13CCO2雨季<旱季。
(2) 麻黄洞土壤空气CO2主要为生物作用来源, 是内源性CO2、深层CO2与大气混合作用的结果, 其季节变化特征主要受温度和湿度干预。基于主成分分析可知, 洞穴CO2变化主要受土壤CO2和通风效应控制。
(3) 碳同位素研究表明麻黄洞洞内CO2来源主要为上覆生物源CO2以及外部大气CO2。雨季, 通风作用弱, 上覆生物作用强, 洞穴CO2主要受洞穴滴水脱气以及外部大气控制, 此时洞穴发生积累效应; 旱季, 通风作用主导洞穴环境, 洞穴CO2主要受外部大气稀释作用影响。
致谢:感谢中国地质科学院岩溶地质研究所蒋忠诚研究员与另一位匿名审稿专家对本文提出的宝贵修改意见。
李坡, 贺卫, 钱治, Bottazzi J. 2008. 双河洞地质公园研究. 贵阳: 贵州人民出版社: 58–101.
梁明强, 李俊云, 周菁俐, 张健, 陈朝君. 2019. 重庆市芙蓉洞空气环境变化特征与影响因素分析. 长江流域资源与环境, 28(4): 226–234.
刘平. 2008. 贵州绥阳双河洞国家地质公园洞穴基本特征及成因探讨. 贵州地质, 25(4): 302–305.
刘子琦, 李开萍. 2018. 贵州石漠化地区降雨期间洞穴 CO2变化特征与其影响因素——以石将军洞为例. 贵州师范大学学报(自然科学版), 36(4): 13–17.
任京辰, 张平究, 潘根兴, 宋林华. 2006. 岩溶土壤的生态地球化学特征及其指示意义. 地球科学进展, 21(5): 504–512.
任坤, 沈立成, 袁道先, 王晓晓, 徐尚全. 2016. 2012–2013年重庆雪玉洞洞穴系统碳循环特征. 地球科学, 41(8): 1424–1434.
田娜, 王义祥, 翁伯琦. 2010. 土壤碳储量估算研究进展. 亚热带农业研究, 6(3): 193–198.
童晓宁, 周厚云, 黄颖, 贺海波, 朱礼妍. 2013. 广东英德宝晶宫CO2浓度的时空变化特征. 热带地理, 33(4): 439–443.
汪炎林, 周忠发, 薛冰清, 李坡, 张结, 田衷珲, 汤云涛. 2020. 喀斯特关键带水–土–气CO2分压垂直转化特征及影响因素. 地理学报, 75(5): 1008–1021.
吴泽燕, 章程, 蒋忠诚, 罗为群, 曾发明. 2019. 岩溶关键带及其碳循环研究进展. 地球科学进展, 34(5): 488–498.
殷超, 周忠发, 田衷珲, 曹明达, 张结, 潘艳喜, 汪炎林. 2017. 土壤CO2与喀斯特洞穴CO2季节变化响应分析. 水土保持学报, 31(4): 304–310.
曾广能, 罗维均, 王彦伟, 李勇, 王世杰. 2019. 喀斯特洞穴甲烷研究进展. 地球与环境, 47(2): 227–234.
张结, 周忠发, 汪炎林, 潘艳喜, 张昊天, 田衷珲. 2018. 短时间高强度旅游活动下洞穴CO2的变化特征及对滴水水文地球化学的响应. 地理学报, 73(9): 79–93.
张萍, 杨琰, 孙喆, 梁莎, 张娜, 田宁, 李建仓, 凌新有, 张志钦. 2017. 河南鸡冠洞CO2季节和昼夜变化特征及影响因子比较. 环境科学, 38(1): 60–69.
赵瑞一, 吕现福, 蒋建建, 段逸凡. 2015. 土壤CO2及岩溶碳循环影响因素综述. 生态学报, 35(13): 4257–4264.
Baldini J U L, Baldini L M, McDermott F, Clipson N. 2006. Carbon dioxide sources, sinks, and spatial variability in shallow temperate zone caves: Evidence from Ballynamintra Cave, Ireland., 68(1): 4–11.
Breecker D O, Payne A E, Quade J, Banner J L, Ball C E, Meyer K W, Cowan B D. 2012. The sources and sinks of CO2in caves under mixed woodland and grassland vegetation., 96: 230–246.
Buczko U, Bachmann S, Gropp M, Jurasinski G, Glatzel S. 2015. Spatial variability at different scales and sampling requirements forsoil CO2efflux measurements on an arable soil., 131: 46–55.
Cao M, Jiang Y, Chen Y, Fa J X, He Q F. 2020. Variations of soil CO2concentration andCO2in a cave stream on different time scales in subtropical climatic regime., 185, 104280.
De Freitas C R, Littlbjohn R N, Clarkson T S, Kristament I S. 1982. Cave climate: Assessment of airflow and ventilation., 2(4): 383–397.
Fairchild I J, Smith C L, Baker A, Fuller L, Spötl C, Mattey D, McDermott F, E I M F. 2006. Modification and preservation of environmental signals in speleothems., 75(1–4): 105–153.
Fichez R. 1991. Benthic oxygen uptake and carbon cycling under aphotic and resource-limiting conditions in a submarine cave., 110(1): 137–143.
James E W, Banner J L, Hardt B. 2015. A global model for cave ventilation and seasonal bias in speleothem paleoclimate records.,,, 16(4): 1044–1051
Krajnc B, Ferlan M, Ogrinc N. 2017. Soil CO2sources above asubterranean cave — Pisani rov (Postojna Cave, Slovenia).17(7): 1883–1892.
Lachnie M S. 2009. Climatic and environmental controls on speleothem oxygen-isotope values., 28(5–6), 412–432.
Mandić M, Mihevc A, Leis A, Bronić I K. 2013. Concentration and stable carbon isotopic composition of CO2in cave air of Postojnskajama, Slovenia., 42(3): 279–287.
Mattey D P, Atkinson T C, Barker J A, Fisher R, Latin J P, Durell R, Ainsworth M. 2016. Carbon dioxide, ground air and carbon cycling in Gibraltar karst., 184: 88–113.
Nickerson N, Risk D. 2006. Keeling plots are non-linear in non-steady state diffusive environments.,20(3): 3029–3083.
Rutlidge H, Baker A, Marjo C E, Andersen M S, Graham P W, Cuthbert M O, Rau G C, Roshan H, Markowska M, Mariethoz G, Jex C N. 2014. Dripwater organic matter and trace element geochemistry in a semi-arid karst environment: Implications for speleothem paleoclimatology., 135: 217–230.
Sánchez-Cañete E P, Serrano-Ortiz P, Domingo F, Kowalski A S. 2013. Cave ventilation is influenced by variations in the CO2-dependent virtual temperature., 42(1): 1–8.
Serrano-Ortiz P, Roland M, Sanchez-Moral S, Janssens I A, Domingo F, Goddéris Y, Kowalski A S. 2010. Hidden, abiotic CO2flows and gaseous reservoirs in the terrestrial carbon cycle: Review and perspectives., 150(3): 321–329.
Shindoh T, Mishima T. Watanabe Y, Ohsawa S, Tagami T. 2017. Seasonal cave air ventilation controlling variation in cave airCO2and drip water geochemistry at Inazumi Cave, Oita, northeastern Kyushu, Japan., 79(2): 100–112.
Treble P C, Fairchild I J, Griffiths A, Baker A, Meredith K T, Wood A, McGuire E. 2015. Impacts of cave air ventilationand in-cave prior calcite precipitation on Golgotha Cave dripwater chemistry, southwest Australia., 127: 61–72.
Vieten R, Winter A, Warken S F, Schröder-Ritzrau A, Miller T E, Scholz D. 2016. Seasonal temperature variations controlling cave ventilation processes in Cueva Larga, Puerto Rico., 45(3): 259–273.
Wang Y W, Luo W J, Zeng G N, Wang Y, Yang H L, Wang M F, Zhang L, Cai X L, Chen J, Chen A Y, Wang S J. 2019. High222Rn concentrations and dynamics in Shawan Cave, southwest China., 199: 16–24.
Study on the response relationship between soil and cave CO2concentrations in Mahuang Cave, Guizhou province
SHI Liangxing1, 2, ZHOU Zhongfa1, 2*, DING Shengjun1, 2, DONG Hui1, 2, FAN Baoxiang1, 2, AN Dan1, 2, TANG Yuntao1, 2
(1. College of Karst Science/School of Geography and Environmental Science, Guizhou Normal University, Guiyang 550001, Guizhou, China; 2. National Key Laboratory Breeding Base of Karst Mountain Ecological Environment in Guizhou Province, Guiyang 550001, Guizhou, China)
CO2plays a vital role as a driving force of karstification. The unique structure of the ground and underground in the karst region underscores the necessity to study the CO2and δ13CCO2of the cave system, which is the window of the underground system. In our study, we monitored the concentration of CO2and δ13CCO2in Mahuang cave and other parameters related to soil, air, cave, and atmosphere for 12 months. The results show that: (1) CO2and δ13CCO2of soil and cave demonstrated evident spatio-temporal variation, with high CO2concentration and lighter δ13CCO2in the rainy season and low CO2concentration and heavier δ13CCO2in the dry season. (2) Soil CO2mixes with endogenous and atmospheric CO2through advective infiltration under different temperatures and humidity, controlling soil bioactivity and leading to the spatio-temporal difference in soil CO2and δ13CCO2. Cave CO2and δ13CCO2are mainly affected by the soil CO2and ventilation effect. (3) The CO2of Mahuang Cave mainly comes from the soil and atmospheric CO2. With the intervention of the ventilation effect, the cave generates a dilution or an agglomeration effect. Exploring the variation characteristics of soil-cave CO2and δ13CCO2and the source of cave CO2will be beneficial to the study of the carbon cycle mechanism of the cave system as well as be of positive significance to the study of the “missing carbon sink” in the global carbon cycle.
karst cave; the source of CO2; carbon stable isotope
X142
A
0379-1726(2022)02-0223-10
10.19700/j.0379-1726.2022.02.006
2020-08-27;
2020-12-23
国家自然科学基金项目(41361081)、贵州师范大学资助博士科研项目(GZNUD[2017]号)和贵州省高层次创新型人才培养计划——“百”层次人才(黔科合平台人才[2016]5674)联合资助。
石亮星(1994–), 男, 硕士研究生, 自然地理学专业。E-mail: Shiliangxing999@163.com
周忠发(1969–), 男, 教授, 主要从事喀斯特生态环境与GIS研究。E-mail: fa6897@163.com
