百合叶烧病发病过程叶片细胞和差异表达基因分析
2022-04-28葛金涛王江英汤雪燕孙明伟腾年军朱朋波赵统利吴秋月邵小斌
葛金涛,王江英,汤雪燕,孙明伟,腾年军,朱朋波,赵统利,吴秋月,邵小斌
(1. 连云港市农业科学院,江苏 连云港 222000;2. 南京农业大学园艺学院,江苏 南京 210095;3. 连云港市海州区农业农村局,江苏 连云港 222006)
0 引 言
【研究意义】百合叶烧病是一种生理性病害,国内外百合种植过程中普遍发生,严重影响切花百合和盆栽百合生产质量。其发病症状表现为百合生长至30~40 cm,花蕾出现前,顶部6~10 片新叶离叶尖约2 cm 处出现灰白色病斑,后期逐渐向叶尖蔓延,最后变成焦枯状。百合叶烧病具有显著的品种特异性,但目前尚未发现与叶烧病存在紧密联系的品种特征[1],因此探索百合叶烧病发病机理,对百合新种质创制和栽培技术创新具有重要意义。【前人研究进展】前人研究发现,通过剪掉一半刚展开的下部叶片[2]、在发病之前喷施氯化钙和硝酸钙溶液[3]、通过手动展开尚未展开的新叶和降低栽培环境的湿度[4]等方法能够降低叶烧病的发生率,同时施用生根剂、微生物菌肥和使用合适的栽培基质均能有效地降低叶烧病的发生[5−7]。【本研究切入点】目前,百合叶烧病发病原因及防治方法等相关研究已较为深入,其中由缺钙导致叶烧病发生已被确认,但由于钙参与多项植物生理调节过程,如何确定叶烧病发生分子机理,以及缺钙导致哪些代谢途径受阻从而产生叶烧病,是今后选育抗叶烧病品种的重要研究基础。【拟解决的关键问题】通过扫描电镜和投射电镜观察东方百合Tarrango 正常叶片、轻度叶烧叶片和重度叶烧叶片超微结构的差异,并通过比较东方百合Tarrango 正常叶片、叶烧叶片、正常叶片喷钙和叶烧叶片喷钙4 种处理的转录组测序数据,以期探索与叶烧病发病相关的分子机理和关键调控基因。
1 材料与方法
1.1 试验材料
以荷兰进口的Tarrango 百合种球为试验材料,种植于连云港市农业科学院东辛农场实验基地日光温室,Tarrango 百合生长至现蕾期,对部分百合叶烧叶片和正常叶片喷施30 mmol·L−1硝酸钙叶面肥一次,其余百合喷施等量清水。24 h 后分别取正常叶片 (TarCK)、叶烧叶片(TNCK)、叶面喷钙正常叶片(TarCa)和叶面喷钙叶烧叶片(TNCa)4 个处理,干 冰冷藏送样检测,每个处理生物学重复3 次。
1 .2 转录组测序及分析试验方法
1.2.1 RNA 提取及处理 上述4 个处理中,取每个处理叶片混合样品1 g,参照TaKaRA 全RNA 提取试剂 操作说明进行叶片总RNA 提取。
1.2.2 文库构建及测序 百合叶片cDNA 文库构建及测 序均委托苏州金唯智生物科技有限公司完成。
1.2.3 转录组数据组装及基因功能注释 百合叶片转录组测序及对获得的数据库 Unigene 的全面分析和注 释,均委托苏州金唯智生物科技有限公司完成。
1.3 百合叶片扫面电镜及透射电镜观察
采集Tarrango 正常叶片、轻度发病叶烧叶片和重度发病叶烧叶片(图1),无菌水漂洗干净后,手术刀裁成3 mm×3 mm 大小,置于电镜固定液中固定2 h,再转移至4 ℃保存,4 ℃冰袋运输送样至武汉赛维尔生物科技有限公司。

图1 不同叶烧程度百合叶片Fig. 1 Leaves from different degrees of ULN-infection注:A,百合Tarrango 正常叶片;B,百合Tarrango 轻度叶烧叶片;C,百合Tarrango 重度叶烧叶片。图2 同。Note: A: Leaf from normal lily plant; B: Leaf from lily plant mildly infected by ULN; C: Leaf from lily plant severely infected by ULN. Same for Fig. 2.
2 结果与分析
2.1 百合叶片叶烧病扫描电镜分析
如图2 所示,近轴面表皮层均无气孔分布,其中Tarrango 正常叶片和Tarrango 轻微叶烧叶片的上表皮细胞横直径为500 μm。而Tarrango 重度叶烧叶片上表皮细胞发生严重皱缩,其上皮细胞的横直径只有312.5 μm。轻度叶烧时,百合叶片上表皮细胞出现轻微的收缩,重度叶烧时,百合叶片表皮细胞表面出现褶皱痕迹。

图2 百合叶片近轴面超微结构Fig. 2 Ultrastructure of lily leaves in adaxial view
如图3 所示,百合叶片远轴面表皮层分布了大量气孔,气孔数量最多的是Tarrango 重度叶烧叶片(图3-A),其次为Tarrango 轻度叶烧叶片(图3-B)和Tarrango 正常叶烧叶片(图3-C)叶片。Tarrango正常叶片、Tarrango 轻微叶烧叶片和Tarrango 重度叶烧叶片气孔直径大小基本一致,均在400 μm 左右。Tarrango 轻度叶烧叶片下表皮层便发生皱缩现象,重度叶烧叶片皱缩程度更加严重。

图3 百合叶片远轴面超微结构Fig. 3 Ultrastructure of lily leaves in abaxial view注:A,百合Tarrango 重度叶烧叶片;B,百合Tarrango 轻度叶烧叶片;C,百合Tarrango 正常叶片。Note: A, Leaf from lily plant severely infected by ULN; B, Leaf from lily plant mildly infected by ULN; C, Leaf from normal lily plant.
2.2 百合叶片透射电镜分析
正常叶片叶肉细胞(图4-A、B)和轻度叶烧叶肉细胞(图4-C、D)液泡较大,叶绿体分布在边缘。重度叶烧叶片的叶肉细胞由于液泡失水,叶绿体分布在整个细胞,细胞较为规则;重度叶烧细胞由于失水,整个细胞皱缩,形状不规则(图4-E、F)。在叶烧病发展过程中,叶绿体结构未发生太大变化,基粒垛叠层数较多且排列规律,3 个处理中叶绿体均含有数量不等的淀粉粒。但重度叶烧叶片叶肉细 胞中,线粒体数量明显较少。

图4 百合叶片透射电镜下超微结构Fig. 4 Ultrastructure of lily leaves shown by TEM注:A 和B:百合Tarrango 正常叶片;C 和D:百合Tarrango 轻度叶烧叶片;E 和F:百合Tarrango 重度叶烧叶片。Note: A and B: normal leaf of lily Tarrango; C and D: mildly ULN leaf of Lily Tarrango; E and F: severely Lily ULN leaf of Lily Tarrango.
2.3 转录组测序组装及unigene 注释分析
使用软件Cutadapt 对测序原始数据(Pass filter data)去除接头以及低质量序列,获得后续信息分析用的过滤数据(Clean data)。4 个处理12 个文库产生了541 393 662 条序列,约80.68 Gb 转录组数据,Unigene 平均长度在148.63 ~149.00 bp,GC 含量变化范围为49.17%~52.32%(表1)。由表2 可以看出,转录组数据经组装产生了19 585 575 个重叠群,349537 条unigenes,平均长度为513.25 bp。在所有unigenes 序列中,序列长度小于500 bp 的占75.92%(表3)。Unigene注释分析显示有124 405 unigenes 获得注释,占unigenes总数的35.59%,其中获得注释较多的数据库分别是Nr 注 释121 501(34.76%)、COG 注 释49 594 条(14.19%)、Swissport 注 释72 421 条(20.72%)和K EGG 注释17 051 条(4.88%)。

表 1 12 个cDNA 文库的过滤数据Table 1 Clean data of 12 cDNA library

表 2 转录组序列组装分析Table 2 Summary of transcriptome assembly

表3 Unigene 的长度及数量统计Table 3 Unigene length and quantity statistics
2.4 差异表达基因筛选及分析
对4 组处理转录组数据进行层次聚类(Hierarchical clustering)分析,喷钙2 个处理TarCa 与TNCa 基因表达模式相近,并且与喷清水2 个处理TarCK 与TNCK基因表达模式存在显著区别,通过基因表达量可以将差异表达基因划分成A、B 两个簇(图5-A),同时通过对转录组数据分组对比分析发现,TarCK/TarCa比较中差异表达基因最多,总数为4 816 个,其中上调表达基因2 419 个,下调表达基因2 397 个;其次TNCK/TNCa 比较中差异表达基因2 782 个,其中上调表达基因1 090 个,下调表达基因1 692 个;TarCK/TNCK 比较中差异表达基因822 个,其中上调表达基因399 个,下调表达基因423 个;TarCa/TNCa 比较中差异表达基因最少,总数为113 个,其中上调表达基因56 个,下调表达基因57 个(图5-B)。
通过PCA(Principal Component Analysis)分析发现,TarCa 与TNCa 两个处理的重复组间的变异较小,并且两个处理间的差异性也较小,而TarCK 与TNCK两个处理间不但处理间的差异性较大,重复组间的变异也比较大(图5-C)。同时,在4 组对照分析中,发现了特异的和共同的差异表达基因,共发现7 185 个差异表达基因,包括5 860 个特异差异表达基因和1 325 个共同差异表达基因(图5-D)。其中TarCK/TarCa 与TNCK/TNCa 共差异表达基因最多,为991 个;其次为TarCK/TNCK 与TarCK/TarCa 共差异表达基因为230 个和TNCK/TNCa 与TarCK/TNCK共差异表达基因为96 个,TarCa/TNCa 与TarCK/TNCK共差异表达基因最少,只有1 个,通过上述数据表明,喷钙处理能显著降低叶烧叶片与正常叶片的差异 表达基因数量。
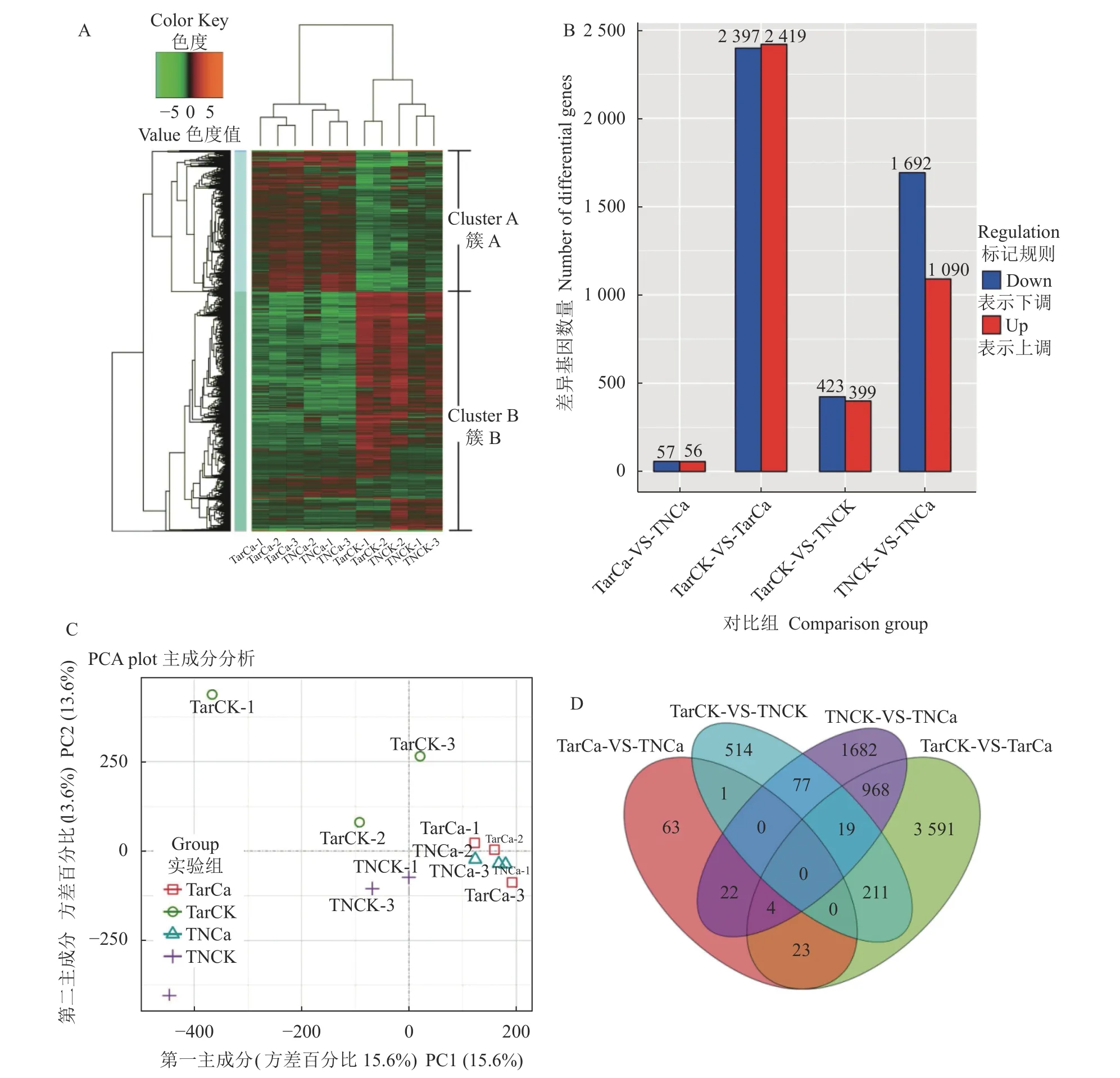
图5 差异表达基因的表达谱分析Fig. 5 Expression profiling of differentially expressed genes注:A,差异表达基因的聚类分析;B,差异表达基因的数量;C,差异表达基因的主成分分析;D,差异基因维恩图分析。Note: A: Cluster analysis on differentially expressed genes; B: Number of differentially expressed genes; C: Principal component analysis on differentially expressed genes; D: Venn diagram of differential genes.
2.5 差异基因GO 富集分析
通过GO 富集分析对差异表达基因进行生物学功能研究,在错误发现率FDR<0.01 时,25 个GO 条目富集在TarCa/TNCa,25 个GO 条目富集在TarCK/TNCK,31 个GO 条目富集在TarCK/TarCa,31 个GO条目富集在TNCK/TNCa。有18 个GO 条目在4 组对比分析中均有富集,其中,“催化活性” “结合”和“代谢过程”是富集基因最多的GO 条目。在4 组对比分析中有5 个特异性GO 条目,其中“多细胞生物过程”和“繁殖”GO 条目仅在TarCK/TarCa 中富集,“突触部分”GO 条目仅在TNCK/TNCa 中富集,“细胞外基质”和“细胞外基质成分”GO 条目仅在TarCa/TNCa 中富集 ,TarCK/TNCK 无特异性GO 条目(图6)。

注:1,催化活性;2,结合;3,转运活性;4,结构分子活性;5,电子载体;6,核酸结合转录因子活性;7,酶调节活性;8,抗氧化活性;9,分子传感器活性;10,细胞组分;11,细胞器;12,膜部分;13,细胞器部分;14,细胞膜;15,高分子复合物;16,胞外区;17,类核;18,细胞连接;19,代谢过程;20,细胞过程;21,单一生物过程;22,生物调节;23,应激反应;24,定位;25,发育过程;26,多生物过程;27,免疫系统过程;28,组织细胞组成或生物起源;29,生殖过程;30,多细胞生物过程;31,繁殖;32,细胞外区域部分;33,细胞外基质;34,细胞外基质成分;35,运动;36,生长;37,突触部分。Note: 1: catalytic activity; 2: binding; 3: transporter activity; 4: structural molecule activity; 5: electron carrier activity; 6: nucleic acid binding transcription factor activity; 7: enzyme regulator activity; 8: antioxidant activity; 9: molecular transducer activity; 10: cell part; 11: organelle; 12:membrane part;13: organelle part; 14: membrane; 15: macromolecular complex; 16: extracellular region; 17: nucleoid; 18: cell junction; 19: metabolic process; 20: cellular process; 21: single-organism process; 22: biological regulation; 23: response to stimulus; 24: localization; 25: developmental process;26: multi-organism process; 27: immune system process; 28: cellular component organization or biogenesis; 29: reproductive process; 30: multicellular organismal process; 31: reproduction; 32: extracellular region part; 33: extracellular matrix; 34: extracellular matrix component; 35: locomotion; 36:growth; 37: synapse part.图 6 差异表达基因的GO 富集分析Fig. 6 GO enrichment analysis on differentially expressed genes
2.6 差异基因KEGG 富集分析
在植物体内,Ca2+参与的代谢调控十分复杂,通过KEGG 代谢通路分析,在本次转录组测序中“剪接体” “代谢途径” “丙酮酸代谢” “次生代谢产物的生物合成”和“光合作用生物的碳固定作用”代谢途径在4 组试验处理对比分析中都有基因富集,经过差异基因COG 分析,4 组试验处理对比分析共有25 个COG分组,其中4 组试验处理共有COG 功能分类22 个,包括“细胞外结构” “信号转导机制” “未知功能基因”“细胞内运输、分泌和囊泡运输” “翻译、核糖体结构和生物发生” “细胞壁/膜/信封生源论” “一般功能预测基因” “复制、重组和修复” “碳水化合物运输和代谢” “无机离子的转运和代谢” “细胞骨架” “氨基酸的运输和代谢” “脂质运输与代谢” “细胞周期控制,细胞分裂,染色体分裂” “能源生产与转换” “染色质结构和动力学” “翻译后修饰,蛋白质转换,伴侣” “辅酶运输和代谢” “RNA 加工和修饰” “次级代谢产物生物合成、运输和分解代谢”。
2.7 显著差异基因分析
结合扫描电镜和透射电镜分析,本研究重点关注叶烧病发病过程和喷施钙处理中,细胞膜通透性相关基因的表达变化。如表4 所示,在百合叶烧病发病过程中即TarCK/TNCK 对比分析中,显著下调基因有参与脱落酸信号的负调控的FOLK 基因(Farnesol kinase 法呢醇激酶)[8]、影响脂类物质生物合成和细胞膜稳定性的PLD1_2 基因(phospholipase D1/2 磷 脂 酶D1/2)[9]、ATPeF1B 基 因(F-type H+-transporting ATPase subunit bet 膜上ATP 合酶)和KCS基 因(3-ketoacyl-CoA synthase 3-酮 脂 酰 辅 酶A 合酶)[10]。同时CALM 基因(Calmodulin 钙调蛋白)[11]、ENO 基因(enolase 烯醇酶)[12]和pel 基因(Pectate lyase 果胶酸裂解酶)[13]等响应钙离子信号和影响木质素和果胶生物合成的基因下调。在百合叶烧病发病过程中表达量上调的基因有促进生成脱落酸的AAO基因(abscisic-aldehyde oxidase 脱落醛氧化酶)[14]。

表4 显著差异表达基因Table 4 Significantly differentially expressed genes
而在叶烧叶片喷钙后即TNCK/TNCa 对比分析中,表达量上调的基因有CALM 基因、CPK 基因(Calcium-dependent protein kinase 钙依赖性蛋白激酶)、EIN2 基因 (ethylene-insensitive protein 2 乙烯不敏感蛋白2)[15]、AUX1 基因(Auxin influx carrier 生长素流入载体)[16]、PLD1_2 基因和果糖和甘露糖的代谢途径的SORD 基因(L-iditol 2-dehydrogenase L-艾杜糖醇-2-脱氢酶)[17]。与此同时下调表达的基因有ABF基 因(ABA responsive element binding factor ABA 响应元件结合因子)[18]和参与脂肪酸降解途径的MFP2基因 (Enoyl-CoA hydratase/3-hydroxyacyl-CoA dehydrogenase 烯酰辅酶A 水合酶/3-羟酰辅酶A 脱氢酶)[19]。
3 讨论与结论
通过百合叶片扫描电镜分析,叶片近轴面表皮细胞大小随叶烧程度的加深而减小,重度叶烧时,表皮细胞发生皱缩,表明此时表皮细胞失水。叶片远轴面表皮细胞在轻度叶烧时发生皱缩,而近轴面表皮细胞在重度叶烧时才发生皱缩,说明叶烧症状先出现在远轴面一侧。通过透射电镜分析,百合叶片在发生叶烧的过程中,液泡失水是导致表皮细胞体积缩小的原因。因此,推测细胞膜和液泡膜的通透性发生改变是导致百合叶片叶烧病症状的原因。
差异表达基因的PCA 分析发现,正常叶片3 个重复处理和叶烧叶片3 个重复处理之间的差异性都很大,而正常叶片喷钙后3 各处理和叶烧叶片喷钙后3 个处理之间差异性较小。同时在对比分析中也发现,TarCa/TNCa 比较组差异表达基因显著少于TarCK 与TNCK,都说明喷钙处理能降低正常叶片和叶烧叶片之间的差异表达基因数量,甚至喷钙处理能够降低正常叶片和叶烧叶片之间差异表达基因数量,说明喷钙处理能缓解叶烧病的发病过程,这与白菜干烧心[20−21]、莴苣[22]等缺钙生理性病害的一致。
在分析单个基因表达中发现,在叶烧病发病过程中,响应钙离子信号CALM 基因显著下调,而喷钙处理后CALM 和CPK 基因显著上调,说明CALM基因是叶烧病发病过程中的主要信号传导基因。同时,叶烧病发病过程中促进生成脱落酸的AAO 基因显著上调,叶烧叶片喷钙后乙烯信号调控基因EIN2和生长素输入载体基因AUX1 表达量上调,表明叶烧病发病可能受脱落酸、乙烯和生长素的调控。同时,叶烧病发病过程中,在细胞膜和液泡膜稳定相关基因phospholipase D1/2、ATPeF1B 、KCS 等表达均显著下调。而喷钙后phospholipase D1/2 显著上调,这与拟南芥中PLD 及其产物磷脂酸(PA)调控ABA响应的负调控因子[23−24]结果一致。
钙在植物生理活动中,既起着结构成分的作用,也具有酶的辅助因素功能,它能维持细胞壁、细胞膜及膜结合蛋白的稳定性,参与细胞内各种生长发育的调控作用。因此目前虽然基本证实百合叶烧病是由缺钙导致,但其具体的致病分子机理仍不能十分明晰,今后仍需开展相关基因分析验证工作,为百合分子育种提供理论支撑。
