基于网络药理学和分子对接探讨肉苁蓉治疗帕金森病的作用机制
2022-04-26徐朝辉
徐朝辉,邹 苑,陈 瑛,梁 彪,赵 卿
(1.上海中医药大学,上海 201203;2.上海市养志康复医院(上海市阳光康复中心),同济大学附属养志康复医院,上海 201619)
帕金森病(Parkinson’s disease,PD)是一种常见的中老年神经系统退行性疾病,临床上以运动迟缓、静止性震颤、肌强直、姿势平衡障碍等运动症状以及睡眠障碍、嗅觉障碍、自主神经功能障碍、认知和精神障碍等非运动症状为显著特征[1]。PD主要病理特征为黑质中多巴胺能神经元的丢失和由α-突触核蛋白(α-Synuclein,α-Syn)组成的路易小体的存在,但发病机制尚未完全明确。目前我国65岁以上人群中PD发病率为1.7%,且随着老龄化程度加剧,据推测我国PD患病人数到2030年将达到500万人,约占到全球PD患病人数的一半[2]。目前PD的药物治疗多以左旋多巴制剂为主,虽然能够在一定程度上改善症状,但长期使用存在药效减退、剂末现象、异动症等运动并发症,且无法从根本上阻止疾病进展。
肉苁蓉,俗称大芸,始载于《神农本草经》,列属上品,味甘、咸,性温,归肾、大肠经,中医认为其具有补肾阳,益精血,润肠通便之效。研究表明,肉苁蓉具有抗凋亡、抗氧化、抗衰老、抗脑缺血再灌注损伤等神经保护作用[3],其化学成分有苯苷类、环烯醚萜及其苷类、木脂素及其苷类、多糖以及单萜苷类、生物碱等,其中又以苯乙醇苷类和多糖为主[4],药理学研究发现肉苁蓉在PD及阿尔兹海默病等神经退行性疾病中发挥的作用显著[5−6]。据统计,肉苁蓉在抗衰老延年类古方中的出现率仅次于人参[7]。目前,肉苁蓉常被应用于PD的临床治疗,但其发挥疗效的具体机制并不明确,本文主要从网络药理学和分子对接的角度,利用相关数据库筛选肉苁蓉的主要活性化合物,并将其作用靶点与PD靶点比对,得出关键作用靶点,探析其治疗PD的物质基础与作用机制,以期为深入研究提供指导。
1 资料与方法
1.1 有效化合物
本研究使用中药系统药理数据库(Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform TCMSP,http://lsp.nwu.edu.cn/tcmsp.php)检索肉苁蓉成分,口服生物利用度(oral bioavailability,OB)≥30%,类药性(drug-likeness,DL)≥0.18[8]为阈值,因血脑屏障功能障碍与PD病程有关[9],故设定血脑屏障通透性(blood brain barrier,BBB)≥−0.85,通过以上限定条件筛选肉苁蓉中的活性成分作为有效化合物。
1.2 查询PD相关疾病靶点
利用PubChem数据库(www.ncbi.nlm.nih.gov/pccompound/)、sea数据库(http://sea.bkslab.org/)获得肉苁蓉所有靶点基因名称。应用GeneCard数据库 ( https://www.genecards.org/) 以 “Parkinson’s disease”为检索词获取PD靶点基因名称。过比对中药和疾病靶点,利用VENNY2.1软件作交集饼图得到PD与肉苁蓉共有靶点基因信息。
1.3 靶点蛋白互作(protein-protein interaction,PPI)网络的构建
将1.1及1.2获得的中药、疾病共有基因输入STRING(https://string-db.org/),选择“Homo sapiens”,将最小相互作用阈值设为中等“Highest confidence>0.9”,其余设置为默认值[10],进行PPI网络构建。
1.4 基因本体(gene ontology,GO)富集和基因组百科全书(Kyoto encyclopedia of genes and geomes,KEGG)通路富集
筛选关键靶基因(大于2倍degree),将筛选出的靶基因导入DAVID6.8数据库(https://david.ncifcrf.gov/summary.jsp),限定物种为“Homo Sapiens”,以P<0.05为阈值,选择其中BP、CC、MF 3对关键靶基因进行GO功能富集分析,筛选具有显著差异的生物过程及可靠的靶点通路,按照P值从小到大排序,前20个作为重要通路用Omicshare (http://www.omicshare.com/) 制作成可视化气泡图。通过KEGG通路富集分析靶点信号通路,以P<0.05为阈值,依据count值排序,选取前20条信号通路选用Omicshare ( http://www.omicshare.com/) 制作成可视化气泡图。
1.5 构建“肉苁蓉-化合物-靶点”网络图
选取前20条KEGG信号通路上的基因作为治疗靶基因,将筛选的治疗靶基因对应的化合物和中药导入到cytoscape软件中,构建“肉苁蓉-化合物-靶点”网络图。
1.6 “活性成分-靶点”分子对接
选取网络药理学筛选出的肉苁蓉有效化合物与帕金森病的关键靶点,利用AutoDock Vina工具进行分子对接,了解靶点蛋白质与活性成分的结合方式。
2 结果与分析
2.1 肉苁蓉有效化合物及靶点基因筛选
使用TCMSP网站筛选出肉苁蓉中药共6个有效化合物,分别 为beta-sitosterol(β-谷甾醇)、arachidonate(花生四烯酸)、suchilactone(脱水内酯)、Yangambin(丁香树脂二甲醚)、quercetin(槲皮素)、Marckine(枫马京)。见表1。根据有效成分筛选出435个靶点基因名称。
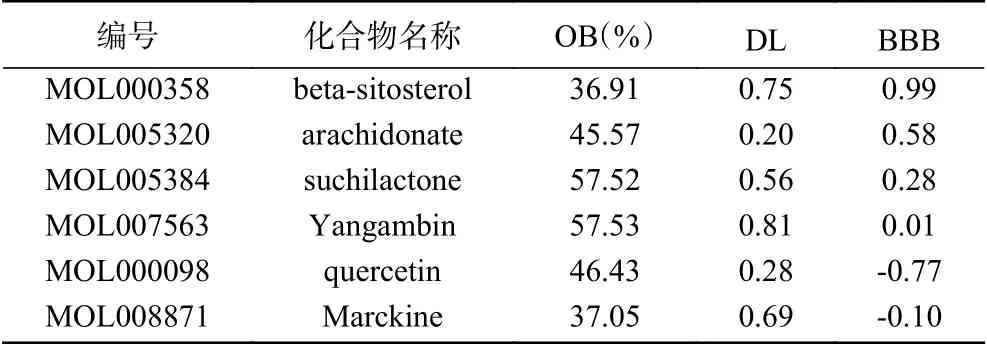
表1 肉苁蓉有效成分Table 1 Effective components of Cistanches Herba
2.2 帕金森病靶点基因筛选
检索GeneCard数据库(https://www.genecards.org/)得到PD相关基因,共计7846个,如LRRK2、SNCA、PRKN、PARK7、GBA、PINK1、MAPT等。
2.3 肉苁蓉活性成分治疗帕金森病预测靶点的筛选
通过TCMSP收集肉苁蓉6个活性化合物对应靶点,去重得到435个蛋白靶标,与GeneCard数据库去重所得7846个蛋白靶标进行比对取交集,运用VENNY2.1软件对疾 病靶点与药物靶点整理后得到308个交集靶点,见图1。蓝色代表PD靶点,黄色代表肉苁蓉靶点,重叠部分为共同靶点。

图1 帕金森病-肉苁蓉靶点交集韦恩图Fig.1 Wayne diagram of target intersection of Parkinson’s disease-Cistanches Herba
2.4 PPI网络的构建及关键靶点筛选
将上述筛选出的靶基因导入STRING(https://string-db.org/)数据库,物种选“Homo sapiens”进行PPI分析,得到肉苁蓉作用于PD的PPI网络图,见图2。共得到283个节点,3509条边。

图2 靶点蛋白互作网络图Fig.2 Protein-protein interaction network
2.5 GO功能富集分析
筛选关键靶基因(大于2倍degree),将筛选出的靶基因导入DAVID6.8数据库,限定物种为“Homo Sapiens”,对38个靶基因进行GO富集分析。结果包括生物过程(biologica process,BP)305条、细胞成分(cellular component,CC)25条、分子功能(molecular function,MF) 69条,按照P值从小到大排序,前20个重要通路用Omicshare作气泡图进行可视化分析,如图3。图中气泡越大富集基因个数越多,气泡颜色越红,P值越小,显著性越高。图3A中BP包括RNA聚合酶II启动子的转录负调控、以DNA作为模板转录、细胞增殖、凋亡过程的正调控、细胞内信号转导等;图3B可见CC分析中包括膜筏、神经元突触、细胞膜、细胞核膜、蛋白质复合物、线粒体等;图3C可见MF富集结果主要为腺嘌呤核苷三磷酸结合、蛋白异源二聚体活性、蛋白质复合物结合、蛋白质酪氨酸激酶活性等。

图3 GO功能富集分析Fig.3 Gene ontology function enrichment analysis
2.6 KEGG通路分析及构建“肉苁蓉-化合物-靶点“网络图
通过KEGG通路富集分析靶点信号通路,以P<0.05为阈值,依据count值排序,选取前20条信号通路,其中主要包括PI3K/AKT信号通路、miRNAs调控、HIF-1、TNF、FOXO信号通路等,依据结果绘制气泡图,见图4。图中气泡越大富集基因个数越多,气泡颜色越红,P值越小,显著性越高。选取前20条KEGG信号通路上的基因作为治疗靶基因,将91个治疗靶基因对应的化合物和中药导入到cytoscape软件中,构建中药-化合物-靶点网络图,见图5。网络中的节点表示靶标或化合物,而边则表示活性成分与PD靶标之间的相互作用,颜色越深,degree值越高。

图4 KEGG信号通路富集分析Fig.4 Enrichment analysis of Kyoto encyclopedia of genes and geomes signal pathway

图5 “肉苁蓉-化合物-靶点”网络图Fig.5 Network diagram of “Cistanches Herba-compound-target”
2.7 “活性成分-靶点”分子对接

从中药-化合物-靶点网络中将基因靶点以degree值高低进行排序,选择前5个关键靶点,分别为丝裂原活化蛋白激酶8(mitogen-activated protein kinase 8,MAPK8)、雌激素受体1(estrogen receptor alpha 1,ESR1)、类固醇受体辅助活化因子(steroid receptor coactivator,SRC)、蛋白酪氨酸激酶2(janus kinase signal transducers 2,JAK2)、丝氨酸/苏氨酸激酶1(serine/threonine kinases 1,AKT1)。利用Auto-Dock Vina[11]分别与肉苁蓉的6个活性成分(β-谷甾醇、花生四烯酸、脱水内酯、丁香树脂二甲醚、槲皮素和枫马京)进行分子对接。结合能<−4.25 kcal/mol提示配体小分子与受体蛋白之间有一定的结合活性;结合能<−5.0 kcal/mol表明二者之间有较好的结合活性;结合能<−7.0 kcal/mol表示配体与受体具有强烈的结合活性[12]。分子对接结果见表2,β-谷甾醇、花生四烯酸、脱水内酯、丁香树脂二甲醚、槲皮素和枫马京6种化合物与关键作用靶点MAPK8、ESR1、SRC、JAK2和AKT1的结合能均小于−5 kcal/mol,说明药物与靶点的结合活性较好[10]。其中β-谷甾醇、脱水内酯和丁香树脂二甲醚与JAK2、MAPK8和SRC,以及槲皮素和枫马京与ESR1、JAK2、MAPK8和SRC的结合能均小于−7 kcal/mol,说明药物与靶点之间具有较强的结合活性。随后利用PyMol(www.pymol.org)对6个肉苁蓉有效成分与其结合活性最高靶点的分子对接结果进行可视化分析,如图6所示。其中β-谷甾醇与活性位点附近的ASN-397形成氢键结合到SRC,花生四烯酸与活性位点附近的ARG-394形成氢键结合到ESR1,脱水内酯与活性位点附近的SER-936和ARG-980形成氢键结合到JAK2,丁香树脂二甲醚与活性位点附近的ARG-980形成氢键结合到JAK2,槲皮素与活性位点附近的THR-338、GLU-339、MET-341和ASP-404形成氢键结合到SRC,枫马京与活性位点附近的PRO-361、GLN-362、GLU517、PHE520和SER-522形成氢键结合到SRC。

表2 肉苁蓉有效活性成分与对应核心靶点的分子对接Table 2 Molecular docking of effective active components of Cistanches Herba with corresponding core targets
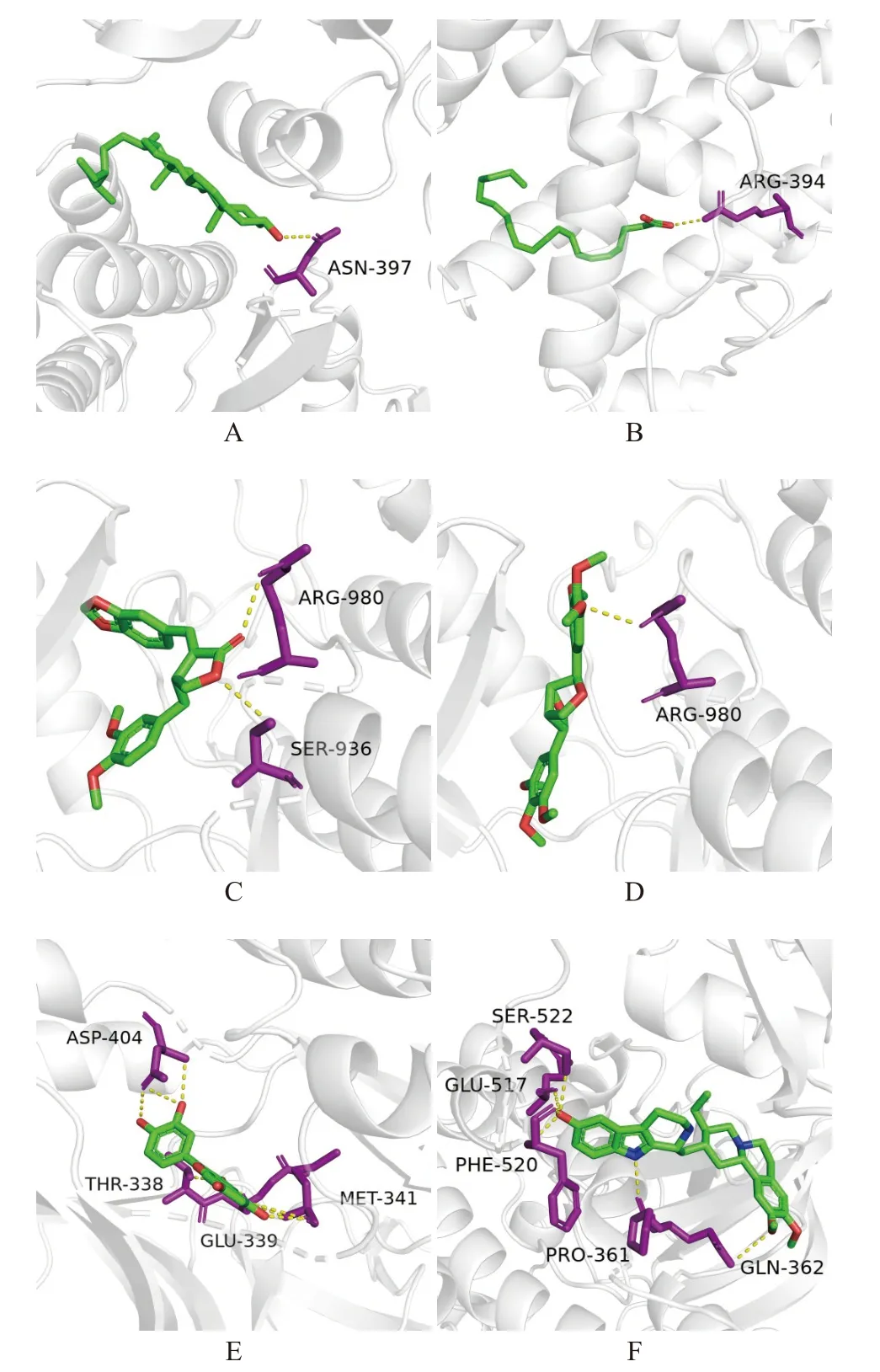
图6 分子对接模式图Fig.6 Molecular docking mode
3 讨论
研究表明肉苁蓉主要成分苯乙醇苷可通过抑制Caspase-3的活性,对抗1-甲基-4-苯基吡啶离子 (1-methyl-4- phylpyridiniumion,MPP+) 导致的多巴胺能神经细胞凋亡[13]。研究也发现松果菊苷作为肉苁蓉活性成分之一,能抑制PD小鼠黑质多巴胺能神经元的减少,且通过GFRa1/AKT信号通路上调胶质细胞源性神经营养因子(glial cell-derived neurotrophic factor,GDNF)表达[14],通过ROS/ATF3/CHOP通路抑制神经细胞凋亡[15],增加纹状体多巴胺神经递质含量,改善PD小鼠的行为学异常,表现出良好的神经保护和修复作用[16]。但关于肉苁蓉其他有效成分在治疗PD中的作用机制仍未探究。
本研究通过筛选发现肉苁蓉的有效活性成分一共6种,分别是beta-sitosterol(β-谷甾醇)、arachidonate(花生四烯酸)、suchilactone(脱水内酯)、Yangambin(丁香树脂二甲醚)、quercetin(槲皮素)、Marckine(枫马京)。研究发现β-谷甾醇具有抗氧化、抑制细胞凋亡等作用[17],而氧化应激损伤是PD发病的关键环节之一[18],推测β-谷甾醇可能通过抗氧化应激治疗PD。花生四烯酸可以通过激活过氧化物酶增殖体激活受体(peroxisome proliferatorsactivated receptors,PPAR)及促进新蛋白合成来增强神经组织内部的抗氧化系统,发挥抗氧化应激的作用,保护神经元细胞[19]。有研究发现槲皮素能够通过抑制AGEs/RAGE/NF-κB炎症通路,从而降低脑组织炎性因子的生成[20]。而另一项研究发现中药提取物可通过抑制晚期糖基化终末产物(advanced glycation end products,AGEs),阻断晚期糖基化终产物受体(receptor of advanced glycation endproducts,RAGE)表达,从而抑制人神经母细胞瘤细胞(human neuroblastoma cells,SH-SY5Y)中 甲 基 乙 二 醛(methylglyoxal,MGO)触发的核因子κB(nuclear factor kappa-B,NF-κB)激活和易位,而已知MGO是一种二羰基代谢物,与神经退行性疾病的发展有关,提示可通过调节AGEs/RAGE/NF-κB通路减轻反应性二羰基化合物的神经毒性[21]。此外,另一项研究发现具有高氧化还原活性的铁可催化大脑氧化反应,还参与产生高毒性自由基(Fenton反应),从而对细胞造成氧化应激损伤,最终导致多巴胺能神经元死亡,而槲皮素可充当强有力的铁螯合剂发挥抗氧化作用,表现出一定的神经保护作用[22]。
GO功能富集分析表明肉苁蓉具有促进ATP合成、保护线粒体呼吸链的功能完整的作用,而线粒体功能障碍是引起多巴胺能神经元变性、死亡机制之一[23]。肉苁蓉还可以调节蛋白质的合成、折叠、降解等活动,维持“蛋白质内稳态”。而研究表明蛋白质构象异常或过度沉积可抑制突触功能,干扰细胞内信号传导,导致由泛素蛋白酶体系统介导的蛋白质降解功能紊乱,最终导致多巴胺能神经细胞死亡[24]。
通过KEGG富集得到活性成分与PD靶标之间相互作用的主要信号通路,包括PI3K/AKT信号通路、miRNAs调控、HIF-1、TNF及FOXO信号通路等,表明肉苁蓉通过多条通路作用于PD发病的调控过程。AKT,又名蛋白激酶B,是一种促进细胞存活的激酶,其存在于磷脂酰肌醇-3-激酶(phosphatidylin-ositol-3-kinase,PI3K)信号途径中。PI3K激活后的产物与AKT的PH结构域相结合,使其Ser473和Thr308位点磷酸化,从而激活AKT。研究发现PD患者黑质致密部(substantia nigra compacta,SNc)中总AKT水平及活性磷酸化AKT水平均显著降低[25]。林瑶等[26]发现肉苁蓉可使PD大鼠黑质纹状体中PI3K、ATK的表达升高及其下游凋亡相关蛋白Bcl-2表达升高、Bax表达降低,提示肉苁蓉通过激活PI3K/AKT通路,从而升高Bcl-2/Bax比值,减少神经细胞凋亡来发挥神经保护作用。糖原合成酶激酶-3β(glycogen synthase kinase-3β,GSK-3β)活性增高,诱导多巴胺能神经元凋亡,而研究发现载有碱性成纤维细胞生长因子(basic fibroblast growth factor,bFGF)的脂质体通过激活PI3K/AKT通路增加GSK-3β的磷酸化,从而抑制GSK-3β活性保护神经元[27]。此外,哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)作为PI3K/AKT下游信号分子,是自噬过程的主要调节因子,而α-Syn的聚集是PD发病机制的核心之一,激活PI3K/ATK/mTOR通路诱发自噬途径使聚集的α-Syn降解,从而发挥治疗PD的作用[28]。由此可见肉苁蓉有效成分可能通过激活PI3K/AKT信号通路促进神经细胞存活发挥治疗PD的作用。同样涉及α-Syn聚集导致PD的发病机制,Pino等[29]发现叉形头转录因子O亚型(forkhead box O,FOXO)家族重要的自噬调控因子FOXO3有效地减少可溶性α-Syn含量,且FOXO3控制神经元细胞中的自噬流量。由此可以推测,肉苁蓉有效成分可能通过FOXO信号通路减少α-Syn的积累保护多巴胺神经元。
miRNAs作为一簇微小的内源性非编码小RNA分子,能够通过序列特异性的方式来调节靶基因表达,参与调控多巴胺神经元增殖、凋亡及自噬等生物过程[30]。研究发现α-Syn(其编码基因为SNCA)的异常表达会诱导多巴胺神经元凋亡,而miRNA-7及miRNA-153可降低PD模型中SNCA的表达水平,改善PD症状[31];同时miR-425可以恢复功能失调的多巴胺能神经院并改善PD小鼠行为缺陷[32]。这些也验证了肉苁蓉通过调控miRNA影响PD发病过程,但具体机制尚需进一步研究。
缺氧诱导因子-1(hypoxia inducible factor-1,HIF-1)是一种对氧敏感的转录激活因子,由组成型表达的β亚基和低氧诱导型的α亚基构成,参与血管生成、铁代谢、葡萄糖能量代谢和细胞增殖及存活的基因转录诱导[33]。有研究发现鱼藤酮或MPP+处理的PD模型SH-SY5Y细胞中HIF-1α的蛋白质水平降低,铁铁螯合剂去铁胺可引HIF-1α的积累,伴随着SH-SY5Y细胞中自噬的增强;当HIF-1α基因被抑制时,去铁胺诱导的自噬也相应受到抑制,表明去铁胺在鱼藤酮诱导的PD细胞凋亡中的保护作用可能是通过HIF-1α积累以及HIF-1α介导的自噬诱导[34]。食欲素A(Orexin-A)是下丘脑神经元分泌的一种神经肽,可在神经病变中发挥保护作用。有研究发现Orexin-A作用于经MPP+处理SH-SY5Y细胞后,不仅诱导HIF-1α的产生,同时激活HIF-1下游靶标如血管内皮生长因子,减轻细胞损伤,起到神经保护作用[35]。可见,肉苁蓉有效成分可能通过增加HIF-1靶基因的表达激活诱导受损细胞自噬,减轻细胞损伤。
越来越多证据支持神经炎症反应参与PD的发病机制[36],而肿瘤坏死因子(tumor necrosis factor,TNF)通路作为重要的炎症反应通路,介导激活炎性小体,进而激活小胶质细胞,活化的小胶质细胞通过增殖和分泌炎症因子可损伤多巴胺能神经元[37]。Liang等[38]用肉苁蓉提取物松果菊苷处理亚急性PD小鼠模型后发现,松果菊苷可减少小胶质细胞和星形胶质细胞的活化,降低炎症因子TNF-α水平,上调GDNF表达,改善PD小鼠行为学症状。
通过构建中药-化合物-靶点网络图,根据degree值高低进行排序,选择前5个关键靶点,分别为MAPK8、ESR1、SRC、JAK2、AKT1。Liu等[39]发现中药提取物可以通过激活丝裂原活化蛋白激酶8(mitogen-activated protein kinase 8,MAPK8),从而促进抗凋亡蛋白Bcl-2 磷酸化,进而清除受损线粒体,间接抑制细胞凋亡,最终通过调节自噬、凋亡再平衡而发挥神经保护作用,可能也是肉苁蓉的作用机制之一。Chakrabarti等[40]发现雌激素受体(estrogen receptor alpha,ESR)激动剂可通过抑制小胶质细胞活性减轻炎症反应发挥神经保护作用;而另一项研究发现雌激素作用于ESR抑制NADPH氧化酶(reduced nicotinamide adenine dinucleotide phosphate oxidase)活性,减少神经炎症标志物,从而减轻神经损伤[41]。由此可以得出肉苁蓉有效成分可能作用于ESR1靶点减轻神经炎症反应治疗PD。与ESR相反,类固醇受体辅助活化因子(steroid receptor coactivator,SRC)作为促炎因子会造成PD小鼠多巴胺能神经元损伤[42]。丝氨酸/苏氨酸激酶1(AKT1)属于AKT家族,是参与细胞存活和凋亡调控信号转导的中心分子,磷酸化时完全激活[43]。有研究显示在PD中,维持AKT1中半胱氨酸296和310的硫醇状态对于AKT1激酶活性和防止其被蛋白酶体降解至关重要[44]。可见,上述四个分子均可能是肉苁蓉保护效应的靶点。再利用AutoDock Vina将以上5个关键靶点与肉苁蓉的6个活性成分(β-谷甾醇、花生四烯酸、脱水内酯、丁香树脂二甲醚、槲皮素和枫马京)分别进行分子对接,均成功对接,结合能均小于−5 kcal/mol,提示药物与靶点的结合活性较好。其中槲皮素和SRC具有强烈的结合活性,结合能为−9.2 kcal/mol,可从抗神经炎症、抗凋亡角度进一步深入研究肉苁蓉的作用机制。
综上,本研究通过网络药理学及分子对接探讨了临床常用于治疗PD的有效药物肉苁蓉作用靶点,发现肉苁蓉通过多组分、多通路、多靶点的协同作用发挥其在PD中的疗效,为进一步研究肉苁蓉神经保护作用机理和深入探讨PD发病机制提供了依据。另外,本研究的筛选标准与中药各组分的口服吸收度及生物利用度均有一定关系,所以并非对所有有效成分都能进行充分分析,这对我们天然药物的开发也有一定启示作用,即应结合中药成分的吸收、代谢等特点,同时考虑不同给药途径及不同个体或组织器官的分布情况进行深入研究。
