AM和EcM菌根特征及其对环境变化的响应*
2022-04-20闫国永邱露瑶黄梦娣邢亚娟王庆贵
闫国永, 卢 洁, 邱露瑶, 黄梦娣, 邢亚娟, 王庆贵
(曲阜师范大学生命科学学院,273165,山东省曲阜市)
0 引 言
植物与土壤真菌之间存在多种相互作用,其中常见及最重要的是真菌—根共生体. 在菌根共生体中,植物和菌根真菌二者互惠互利:菌根真菌改善其宿主植物的营养状况、影响生长发育、增强矿物质营养和水分的吸收以及抗病能力,而宿主植物为共生真菌提供生长和繁殖所必需的养分[1],二者达到一种互利互助、互通有无的高度统一. 据估计,约85%的陆地植物物种与菌根真菌形成共生关系[2]. 与植物共生的2种主要菌根类型是丛枝菌根(AM)和外生菌根(EcM),它们在环境偏好、形态和功能上有很大差异[2]. 因此,EcM和AM菌根对主要环境变化因子(全球变暖、大气氮沉降、大气CO2浓度增加和干旱)的响应方式不同.
1 AM和EcM菌根简介
在AM菌根中,真菌首先形成附着结构穿过外根细胞层,形成松散的细胞内线圈和分枝菌丝,最后到达内皮层细胞,在那里产生树状的丛枝,菌丝被丛枝周膜包裹. 在EcM菌根中,菌丝不伸入根部细胞,在表皮细胞之间发育,菌丝体紧密地包围植物幼嫩的根,形成菌套,有的向周围土壤伸出菌丝,代替根毛的作用(表1).

表1 AM和EcM菌根结构特征
2 AM和EcM菌根的生态功能
生物的形态结构决定其在生态系统中的功能. 基于上述AM和EcM菌根的形态特征的差异,它们的生态功能也存在较大区别.
2.1 丛枝菌根(AM)生态功能
AM菌根能够对生物和非生物胁迫有更高耐受性,例如表现出的抗旱性、抗水性、耐盐性以及抗病性等. 通过菌丝吸附在内的多种机制增加土壤聚集性和团聚体的水稳定性[3],从生态水平上改善土壤质量;优化磷的获取,AM菌根宿主植物吸收的所有磷几乎都来自AM真菌;增加宿主植物生物量,主要原因是外部菌丝扩大了根系吸收养分的空间,AM真菌可能激活了土壤中不溶性无机磷酸盐和有机磷酸盐与磷细菌之间的相互作用[3];提高作物、饲料、水果的品质和产量(如谷物的强化),并且在作物收获后可增强食物储存性能[4];与宿主植物紧密联系的优势克服了真菌因丧失腐养能力而产生的风险[5].
2.2 外生菌根(EcM)生态功能
EcM菌根能够参与土壤有机质中氮的酶解,主要是EcM真菌产生木质纤维素分解酶降解土壤有机质并将降解土壤有机质中的氮传递给宿主植物[5]. 其次,EcM菌根在森林管理中也具有巨大潜力,菌根与宿主之间的互助关系帮助林火后森林资源的更新和矿山的生态恢复[2];最后,EcM真菌因为借助与宿主植物的共生关系帮助宿主植物吸收营养物质,所以可以将EcM真菌接种用于针叶林和阔叶林的育苗,帮助幼苗生长[2].
3 AM和EcM菌根氮的吸收
AM(丛枝菌根)真菌和EcM(外生菌根)真菌是自然界中最常见的两种共生真菌,它们对氮的吸收利用存在明显的不同:AM真菌主要利用土壤的无机氮;EcM真菌主要通过分泌多种降解酶来降解土壤有机质中的氮,EcM真菌也可以少量利用小分子无机氮(图1).
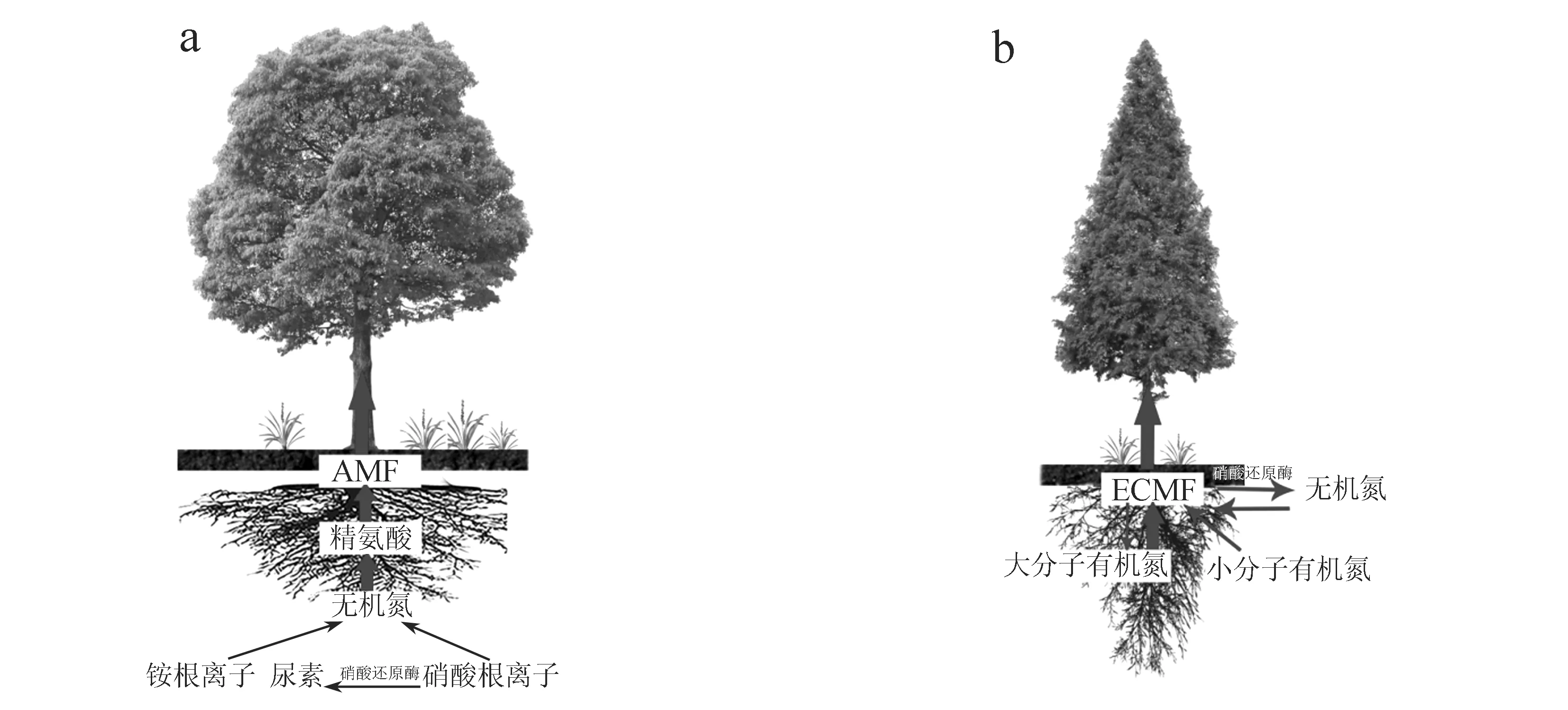
图1 (a)AM菌根对氮的吸收利用方式 (b)EcM菌根对氮的吸收利用方式
铵根离子和尿素是AM菌根吸收氮的两种主要形式,通过尿素循环途径以及高效的无机氮同化,合成有机态的精氨酸,有效组织铵根离子进入菌根组织,从而供给宿主植物需要的氮[6];AM菌根也可以利用无机硝酸根离子,但只有借助硝酸还原酶的还原作用将硝酸根离子转化为铵根离子,重新进行上述铵根离子和尿素的利用方式合成精氨酸才能被AM真菌和宿主植物利用[7-8]. EcM真菌分泌降解酶使土壤有机质中的氮降解传递给宿主植物,这种对氮的吸收利用途径是EcM菌根吸收氮最主要的方式;小部分的EcM真菌从土壤中吸收小分子的有机氮类如氨基酸和氨基糖[1,9],降解蛋白质并将降解后得到的氮转移到它们的宿主植物供宿主植物利用[10];此外极少部分的EcM真菌还可以分泌硝酸还原酶利用硝酸根离子进行无机氮的吸收利用.
4 AM和EcM菌根对磷吸收
磷作为植物生长的必需营养元素,参与细胞分裂,物质及能量合成,在植物生长发育过程中发挥重要的作用. 自然界中磷存在形式有两种:有机和无机形式,通常自然界中存在的无机态磷比有机态磷多,无机态磷占全磷的50%~90%,而许多有机态磷不能直接被植物根系获取[11].
大气CO2浓度升高等环境因素的作用会触发宿主植物枝条中依赖于过氧化氢的生长素的合成,随后生长素通过极性运输到达根部,诱导根中独脚金内酯(strigolactone,SL)的合成. AM菌根中存在着一种独特的信号级联,被定义为氧化还原—生长素—SL系统信号级联. 在土壤中缺乏AM真菌时,生长素和SL共同参与调节根系结构,这种信号级联可能会改变根系结构(RSA)来增强宿主植物对磷的吸收. 在土壤中不缺乏AM真菌时,SL作用下真菌菌丝吸收有机磷之后再传递给宿主植物,进而增加土壤对有机磷的吸收[12].
EcM真菌含有特殊的高亲和力无机磷酸盐(Pi)转运体,能够获得土壤中低浓度的Pi[13]. EcM菌根对磷的利用主要有2种形式:(1)EcM真菌能够溶解各种无机矿物来源的磷(如磷灰石等),为自身和宿主植物提供Pi的来源[14](图2). (2)EcM真菌产生磷酸酶,使有机来源的磷(如磷脂、核酸和蛋白质等)被释放出来,随后被高亲和力Pi转运体摄取[15]. 一旦Pi被EcM菌丝吸收,小部分被继续转移到宿主植物中供宿主植物利用,而大部分主要以聚磷酸盐 (poly Pi) 的形式储存在真菌菌丝中[16],并不能传递给宿主植物. 在针叶树中,Pi从EcM菌丝转移到宿主树需要Pi穿过真菌质膜进入宿主外质体空间,然后穿过根表皮的质膜,在较小程度上穿过皮层细胞,EcM菌根的Pi转移主要发生在Hartig net内[16].
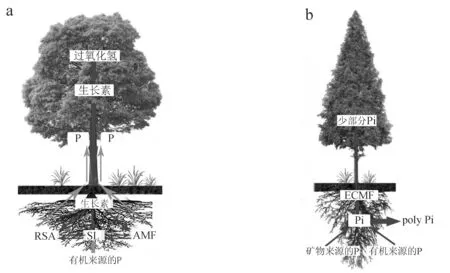
图2 (a)AM菌根对土壤中磷的吸收 (b)EcM菌根对土壤中磷的吸收
5 AM和EcM菌根对于4种环境因子的响应
由于AM和EcM菌根在结构和功能上的差异,导致AM和EcM菌根对氮沉降、温度升高、CO2浓度升高和干旱的响应并不完全一致.
5.1 氮沉降对AM和EcM菌根的影响
基于先前的研究发现,氮沉降对AM菌根的影响方向并不一致[9,17],氮沉降对AM菌根会产生正效应[18]、负效应[19-20]或无影响[17]. Wolf等(2003)研究中的数据表明添加氮的实验组对AM真菌的孢子群落没有显著影响,这主要验证氮沉降对AM菌根无影响. 氮沉降对AM菌根影响方向很大程度取决于研究地点土壤养分背景值的差异. 在磷受限制的土壤中,氮沉降可以增加AM真菌生物量促进其生长,表现出正效应;相反在磷充足的土壤中氮沉降会减少AM真菌生物量抑制其生长[21],表现出负效应. 土壤氮含量也调节AM菌根对氮沉降的响应,在氮素匮乏的土壤,一定程度的氮沉降会改善土壤的养分机制,可以增大对菌丝生物量的合成[22],表现出正效应. AM菌根对不同植物功能类群的响应也不同,具体来说,C3和C4光合途径植物的菌根对氮富集的响应会因它们在真菌营养方面的差异而不同[23].
已有大量研究显示氮沉降对EcM菌根既表现出正效应又表现出负效应,其中对负效应的描述较多,例如减少菌丝数量抑制菌丝体生长,同时使真菌子实体生产力和丰富度降低,降低真菌生物量,抑制腐生微生物的降解过程[24-25]. 但是也有研究发现氮沉降可能增加真菌生物量[26-27]. EcM菌根对氮沉降表现出不同响应可能是由于初始土壤氮含量不同. 在土壤氮充足时,EcM菌根会下调氮的固定,使根瘤生物量减小,但对于一些非氮固定的幼苗仍然有平等获取氮的机会,从而会降低它们相对于非氮固定幼苗的优势[28],因此表现出真菌生物量降低的负效应. 但在北极苔原等营养非常缺乏的系统中,氮沉降实际上可以增加EcM真菌的生物量,原因是可以减轻真菌自身的氮限制[29],表现出正效应.
5.2 变暖对AM和EcM菌根的影响
截至目前最新的AM和EcM菌根对温度升高的响应数据中,63%的研究表明,温度升高增加了菌根的丰度,17%的研究发现温度升高时菌根的丰度有所下降,20%的研究发现温度升高时菌根的丰度没有变化;相比之下,温度升高时菌根的活性普遍下降(71%的研究),其余研究报告菌根活性在上升(14%)或没有变化(14%)[30]. 在此数据的基础上,进一步展开AM和EcM菌根对温度升高这一环境因子的响应机制的研究.
AM对升温的积极响应表现为AM真菌的孢子萌发率升高,增强碳分配,增加对磷的获取,也能增加宿主植物抗旱能力和植物生物量[31-32],表现的负效应为AM真菌定植量和多样性的下降,降低侵染率和减少泡囊数量[33-34]. 针对AM菌根对增温表现出不同的效应来分析原因:(1)AM在湿润的条件下对宿主植物的生长具有促进作用,而温度升高会伴随着土壤湿度的下降,从而抑制AM真菌生长[32,34]. (2)随着温度的升高,会使土壤的碳、氮储量下降,AM真菌必须要和增强优化碳的分配来应对土壤元素储量的下降[34].
EcM真菌随着温度升高表现出的正效应主要是定植量的增加,提高宿主植物的酶活性使其光合速率增加[35],但温度升高也可能导致EcM真菌相对丰富度显著减少[36]等负效应. 我们根据已有的实验研究来分析原因可能是:(1)环境异质性可以影响植物基因型和真菌基因型交互作用的结果和强度,导致“选择镶嵌”[37]. 温度升高带来的是干旱条件的增强,宿主植物会将资源提供给可以抵抗干旱的EcM真菌,导致EcM真菌丰富度会呈现明显的下降趋势[38]. (2)地理纬度的特殊性对EcM真菌响应升温带来显著差异. 世界冻土带地区植物共生EcM在面对增温时,通常会表现出更强的生长和繁殖能力,缓解因气温过低对植物和真菌的抑制性作用;但在较低纬度的地区,气温增加导致EcM真菌的相对丰富度显著减少[39].
5.3 CO2浓度升高对AM和EcM真菌的影响
大气CO2浓度升高,AM真菌的菌丝长度和定植会下降,其生物活性会降低,或无影响等[31,40]. 造成大气CO2浓度升高对AM菌根产生影响不同的原因可能为:(1)在较高的大气CO2浓度下,植物生长、根系定植、孢子和自由基外菌丝的产生会因AM真菌的宿主植物种类的不同而增强或减少. (2)AM真菌生长环境的不同导致其对CO2浓度变化的响应存在差异. 例如,在低氮条件下,AM真菌的宿主植物对CO2浓度在一定范围的变化没有响应,而在高氮条件下,AM真菌的宿主植物的生长会受益于CO2的增加[40].
对于EcM菌根,大气CO2浓度升高,会表现出定植、侵染率、菌根丰度增加的正效应. 同时,大气CO2浓度升高也会对EcM菌根产生菌根数量、定植、丰度等生长状况的减弱和生物活性的降低等负效应,或不产生影响[41]. 与AM菌根不同的是EcM菌根对大气CO2浓度变化的响应受宿主植物类型和生长环境条件的影响并不显著[31]. CO2浓度升高对EcM内部不同种真菌影响不同,会导致真菌群落结构发生变化[42]. 不同种类真菌对CO2的适应程度与响应不同,使得在CO2浓度升高时,不同种真菌的生存优势不同,对真菌群落的丰富度产生影响. Walker等(2006)[43]研究发现大气CO2浓度升高对EcM菌根的影响是阶段性的,随着生长年龄的增加,其侵染率逐渐增高. 除此之外,大气CO2浓度升高的情况下,绝大多数EcM菌根表现出生长速率加快的正效应,但这可能导致菌根真菌的寿命降低,对其产生不利影响[41].
5.4 干旱对AM和EcM菌根的影响
对于AM菌根而言,干旱情况对其菌根生长侵染和优势度改变影响较小,原因在于AM真菌多存在于干旱、温暖的地带,研究表明在亚热带沙漠和草原地区多以AM树为主,故AM真菌受干旱条件的制约较小,影响不大[34]. 干旱条件对于AM真菌产生的负效应多是出现在极端环境下,如极度或长期干旱的地区,AM真菌会表现出一系列的负效应,生长会受到抑制、菌丝变短、分支减少、对水和养分的吸收受到抑制,长期干旱的土壤中真菌过度缺水,其正常新陈代谢受到阻碍、产孢子率会下降,从而导致侵染率降低. 干旱对AM菌根的正效应出现在大多数环境适中的土壤条件下,短暂的干旱会激活AM真菌的抗旱机制,包括:(1)菌丝对水分的直接吸收作用和间接调节作用;(2)AM真菌可以提高与抗旱性相关的酶(SOD,POD,CAT)活性,改善水分状况;(3)增强非酶促反应系统(如可溶性蛋白)的活性;(4)AM真菌分泌的球囊霉素糖蛋白能够促进水分稳定的土壤团聚体的形成;(5)通过改善植物矿质营养状况及内源激素平衡状况来间接影响宿主植物的水分代谢[44]. 因此,在干旱条件下,AM真菌可以提高植物水分利用效率,增强抗氧化保护酶活性,降低丙二醛含量,能显著提高植物抗旱性,缓解干旱胁迫对植物造成的伤害.
对于EcM菌根而言,干旱对其产生的正效应主要包括:(1)形成子实体;(2)EcM真菌外延菌丝伸长,改善宿主营养状况(尤其增加对磷的吸收)[45];(3)外延菌丝和Hartig net的形成对根—土界面液流阻力的降低是至关重要的[45],植物在形成外生菌根共生体后,植物的根系上形成的菌套、菌索、外延菌丝和Hartig net等结构,扩大植物的土壤有效利用空间和根系的吸收面积,此外,菌套的形成对干旱条件下营养根的水分外渗起到了阻隔作用;(4)植物自身的活性氧代谢过程的4种关键酶:SOD(超氧化物歧化酶)、CAT(过氧化氢酶)、APX(抗坏血酸过氧化物酶)、GR(谷胱甘肽还原酶),活性都迅速上升[45]. 干旱对EcM真菌群落的负效应主要是,EcM真菌生长受阻,由于宿主植物缺水使有机物供应减少,加之自身水分吸收下降,EcM真菌生长所需的营养不足,对其生长繁殖造成威胁[45],降低群落结构中EcM真菌的多样性[45],EcM真菌子实体的形成受到抑制[45]. 由于真菌菌根种类不同,其抗旱能力存在一定差异,其对干旱这一胁迫条件产生的响应也有所不同.
6 总结与展望
本文从菌根共生的生态效应出发,结合生态环境发展状况,将近年来有关菌根共生体的生态学作用以及其对不同环境因子变化的响应有关的研究进行归纳汇总,更加系统直观地呈现了菌根共生体在生态系统中的具体作用与价值. 首先对AM和EcM菌根所属纲门、宿主植物类型、侵染部位及各自的结构特征进行简要介绍区分,它们各自侵入宿主植物根细胞的方式也存在不同. 其次选择了土壤中的两大限制因子氮和磷,发现AM和EcM对这2种因子吸收利用存在着显著差异. 如今全球气候变化对许多生态方面产生巨大影响,选择从氮沉降、变暖、CO2浓度升高以及干旱这几方面入手,总结出AM菌根和EcM菌根系统在形态和生理方面的响应,这对后续菌根共生体响应气候变化和应对气候变化的科学进展会起着一定的导向作用. 已知菌根真菌种类繁多,菌根共生体亦然,然而目前的研究较多是针对AM菌根和EcM菌根,单独研究介绍其功能和影响因素,缺乏对多种菌根共同形成的共同菌丝网络的研究和发现,未来可以考虑从此着手,发散思维,着眼于共同菌丝网络,探索其与生态环境间更深层的联系.
