稻纵卷叶螟对性信息素和花香的嗅觉行为反应及在其种群监测中的应用
2022-04-19郭前爽卓富彦朱景全林宇丰张政兵黄德超张素丽杜永均
郭前爽, 卓富彦, 朱景全, 林宇丰, 张政兵,黄德超, 张素丽, 杜永均,*
(1. 浙江大学农药与环境毒理研究所, 杭州 310058; 2. 全国农业技术推广服务中心, 北京 100125;3. 湖南省植保植检站, 长沙 410000; 4. 广东省农业有害生物预警防控中心, 广州 510500)
稻纵卷叶螟Cnaphalocrocismedinalis是重要的迁飞性害虫,卷叶危害,大发生时显著影响水稻产量。水稻卷叶虫的Cnaphalocrocis属有多种种类,其成虫外形类似(冯波等, 2017b),非分类专业难于分辨和鉴定,在测报灯下破损的外形更难以正确鉴定,且稻纵卷叶螟对灯趋光性并不强。各地普遍采用清晨赶蛾的方式进行测报,但赶蛾测报方法工作量大、误差也大(姚海峰等, 2018)。因此,如何准确监测田间稻纵卷叶螟的种群动态一直是生产上迫切需要解决的难题。为此,植保部门一直在比较、评价不同的监测方法,包括赶蛾、性诱,甚至提出了花香诱等多种方法。但对于植保推广部门,这些技术仅仅是监测到了诱蛾的数量,无法深入了解田间成虫的生理状态,如年龄、迁飞、交配和产卵的状态,从而难以实现精准测报和施药。
性信息素诱捕的靶标害虫是性成熟的、正在求偶的雄蛾。多年、多地的田间试验证实了性诱监测稻纵卷叶螟的有效性(Kawazuetal., 2004; Wuetal., 2013),其专一性对于测报的自动化、数字化和智能化更具意义。稻纵卷叶螟雌成虫释放的性信息素最早从菲律宾和斯里兰卡种群中鉴定为Z13-18∶Ac和Z11-16∶Ac,其合适比例为10∶500, 而印度种群的则为500∶50,其后在印度种群中又鉴定出Z13-18∶OH (Raoetal., 1993, 1995; Kawazuetal., 2000)。Kawazu等(2000, 2001)鉴定出日本种群的性信息素为Z11-18∶Ald,Z13-18∶Ald,Z11-18∶OH和Z13-18∶OH(11∶100∶24∶36),两醛类化合物是必需的,醇类化合物起协同增效作用。我国不同地区对稻纵卷叶螟的性信息素经过重新鉴定核对和配比优化,基本接近日本报道的结果(Kawazuetal., 2002; Wuetal., 2013)。但在实际田间诱捕中,性诱稻纵卷叶螟的不稳定性是性诱测报的一个显著弱点,迁飞性、季节性、卵巢和精巢发育等与稻纵卷叶螟性诱密切相关(冯波等, 2017a)。由此人们希望采用寄主植物气味或花的气味来引诱两性害虫,相关报道很多,如从玉米Zeamays、油菜Brassicanapus、向日葵Helianthusannuus、棉花Gossypiumhirsutum、扁豆Lablabpurpureus、鹰嘴豆Cicerarietinum、苦苣菜Sonchusoleraceus、亚麻Linumusitatissimum、苹果Malusdomestica、桉树类Eucalyptus等植物鉴定出来的(Knight and Light, 2001; Xiaoetal., 2002; Del Socorroetal., 2010; Greggetal., 2010),或从茉莉花Jasminumofficinale、金盞花Calendulaofficinalis、灰菜Hirschfeldiaincana、飞蛾草Araujiahortorum、山桃草Gauralindheimeri、菊苣Chicoriumintybus、牛膝菊Galinsogaparviflora等花气味中发现的(Heathetal., 1992; Landolt and Smithhisler, 2003)。因为稻纵卷叶螟、东方粘虫Mythimnaseparata、草地螟Loxostegesticticalis等害虫是迁飞性昆虫,通常认为羽化后需要补充营养,也因此从花香气味的引诱去思考,以期找到理想的引诱剂。花香诱的试验报道很多,主要针对鳞翅目和双翅目昆虫(Meagher, 2002; Meagher and Landolt, 2010)。但因为是基于花香气味的活性化合物并不具有性信息素那样的专一性,因此,对非靶标害虫、传粉益虫、甚至自然天敌的引诱反应也十分强烈(Meagher and Mitchell, 1999; Rohrigetal., 2008)。基于在生态系统中存在这些负面的作用,花香诱捕逐步以应用于生物多样性调查为目的的居多(Landoltetal., 2007),是否可以应用于害虫的监测和防控有待更多的田间试验评价。
因为稻纵卷叶螟的迁飞状态、幼虫虫源地和生境的差异,成虫的营养、交配和卵巢发育等生理状态比较复杂,而且成虫还可以多次交配,迁飞与交配可以同时进行,因此常常出现同时同地的稻纵卷叶螟具有不同的寿命、年龄、性发育水平、交配状态等差异。张孝羲等(1981)依据卵巢发育为Ⅰ级的雌蛾百分比、交配率以及种群增长和下降3个指标来判断迁飞状态,即基本迁入型、部分迁入型、本地繁殖型和大部迁出型。本研究基于张孝羲等(1981)的工作基础以及我们多年田间性诱监测实践,在广西、湖南、浙江和四川5个试验点,通过田间网捕及花香气味和性信息素诱捕稻纵卷叶螟成虫,以及调查田间各龄虫量及其动态,结合解剖卵巢和精巢,探索迁飞性稻纵卷叶螟生理状态和交配、产卵行为的时间和空间规律,建立性诱、花香诱与田间幼虫发生动态之间的关系,为田间稻纵卷叶螟的精准施药提供技术依据。
1 材料与方法
1.1 性诱和花香诱捕及网捕
1.1.1性信息素诱芯:稻纵卷叶螟性信息素组成和剂量参照Wu等(2013),由顺13-十八碳烯醛(500 μg)、顺11-十八碳烯醛(50 μg)、顺13-十八碳烯醇(120 μg)和顺11-十八碳烯醇(90 μg)组成,诱芯材料采用聚氯乙烯(PVC)毛细管灌液结构诱芯,由宁波纽康生物技术有限公司提供。
1.1.2花香诱芯:花香诱芯化合物组成包括苯甲醛、苯乙醛、苯乙醇、水杨酸甲酯、里那醇等,为非专一性,通过黑色通气改性聚乙烯(PE)塑料袋(长86 mm,宽72 mm,厚0.15 mm)控制释放速率,诱芯中化合物总量20 g,测定日释放量平均0.13~0.15 g,由宁波纽康生物技术有限公司配制和提供。
1.1.3诱捕器:试验用塑料材质的、倒漏斗新型飞蛾诱捕器由宁波纽康生物技术有限公司提供。性信息素诱捕:每个诱捕器安装一枚性信息素诱芯,水稻秧苗时,诱捕器底口离地约0.8 m,水稻长高后,底口低于水稻叶面10~20 cm。花香诱捕:每个诱捕器安装一枚花香气味诱芯,诱捕器底口离地约1 m,水稻长高后,底口高于水稻叶面10~20 cm。试验点诱捕器完全随机设计,挂放按直线排列,诱捕器之间间隔为30 m。每种诱捕器设置6个重复。诱捕器排列顺序在各试验重复点中是按先后次序换位的,以减少位置和风向的影响。在各地选取长势相近,管理一致的田块进行实验,将性诱诱芯或者花香诱芯装入组装好的诱捕器组装的飞蛾诱捕器中,标记后插入稻田中。每天早上将诱捕器内的稻纵卷叶螟收集起来并计数记录,一个月更换一次诱芯。
1.1.4田间成蛾的网捕方法:每日清晨07∶00-09∶00时用捕虫网轻轻拨扫水稻或田边杂草的中上部,将虫惊起后,用捕虫网快速将飞动的稻纵卷叶螟收集起来,转移至网笼中,带回实验室解剖并记录其数量。需及时清理出其中的蜘蛛等捕食性昆虫,以免在解剖前稻纵卷叶螟被天敌捕食。
1.2 诱捕及网捕试验时间和地点
2020年8月20日至10月18日于广西省壮族自治区百色市田阳区新山村(23°50′N, 106°50′E)水稻田中,水稻处于分蘖期开始试验,在诱捕器设置时,还没有卷叶危害;2020年9月5日至10月25日在湖南省株洲市攸县新市镇(27°10′N, 113°14′E)水稻田中进行试验,开始试验时水稻处于分蘖期,田间已有大量的卷叶危害;2020年8月21-26日在四川省绵阳市江油市(31°54′N, 104°48′E)水稻田;2020年7月19日-9月29日在浙江宁波江北区慈城(29°58′N, 121°25′E)和宁波市东吴镇(29°49′N,121°74′E)水稻田。
1.3 解剖方法
将准备好的50%酒精放入培养皿中,雌虫腹部取下后放入培养皿中,用眼科镊子将表皮轻轻撕开,使卵巢暴露出来,去除脂肪体等组织,观察其卵的形态、大小等指标,确定卵巢等级拍照并记录。卵巢发育级别分5级,各等级区别标准参考张孝羲等(1979)。记录精包数量,精包数量计算为交配次数。解剖雄蛾时将其生殖系统与表皮及其他脂肪组织分开,精巢取出拍摄照片,方法参照冯波等(2019),将精巢作为椭球计算体积,利用软件Motic Plus Images 2.0分别测量了精巢赤道半径a,b(沿着x和y轴)和极半径c的长度,按照椭球体积公式V=4πabc/3计算桑螟精巢体积。拍照设备使用Motic显微镜SMZ-168和照相系统Moticam 2506(麦克奥迪, 厦门)。
1.4 田间调查
广西田阳、湖南攸县的水稻稻纵卷叶螟幼虫田间调查方法参照农业部稻纵卷叶螟测报技术规范(GB/T 15793-2011),蛾量突增后开始调查。选取长势好、中、差的3块田,采用双行平行跳跃式取样,每块田查10丛,记录卵量、幼虫数量及发育等级。每3 d调查一次。
1.5 田间诱蛾量的昼夜节律记录方法
采用宁波纽康生物技术有限公司生产的新型害虫自动监测工具“赛扑星”昆虫性诱电子智能测报系统(SPT-N-02)监测每日、每小时田间诱蛾动态。性诱智能测报系统每隔1 h发送一次数据。监测系统设置于湖北省黄冈市黄梅县采用花香诱监测(29°58′N, 115°53′E)和性诱监测(99°47′N, 116°10′E)。
1.6 数据分析
数据分析采用软件 SPSS 17.0。各试验点稻纵卷叶螟网捕、性信息素诱和花香诱的雄性精巢体积差异比较,以及四川江油稻田网捕、草丛网捕和花香诱雌性不同卵巢交配次数差异采用单因素完全随机设计的方差分析(one-way ANOVA),其中多组平均数差异性均采用 LSD(least significant difference)法比较。各试验点花香与性信息素诱捕雄蛾的数量差异、各试验点不同虫源类型网捕和花香诱雌性不同卵巢交配次数的差异,采用 Student氏t检验法比较。
2 结果
2.1 试验地稻纵卷叶螟虫源的迁飞和交配状态
参照张孝羲等(1981)的定义标准,各试验点采用网捕田间稻纵卷叶螟雌蛾,卵巢解剖、拍照并分级和统计卵巢发育级别、交配次数,以及田间虫量调查,从卵巢发育为Ⅰ级的雌蛾百分比、交配率以及种群增长和下降得到表1。由此,我们界定了每一个试验点在试验阶段种群雌蛾的迁飞状态。

表1 各试验点稻纵卷叶螟雌蛾的生理状态(2020年)
2.2 性信息素诱与花香诱诱捕稻纵卷叶螟雄蛾数量比较
湖南攸县雌蛾为本地繁殖型,性信息素日诱捕稻纵卷叶螟雄蛾的平均数量显著高于花香诱捕的(P<0.05)(图1: A)。浙江宁波江北雌蛾为基本迁入型,性信息素诱日诱捕雄蛾平均数量与花香诱捕的比较无显著差异(P=0.416)(图1: B)。8月23日-9月4日的广西田阳种群为基本迁入型,性信息素诱日平均数量也显著高于花香诱捕的(P<0.05),后期(10月1-7日)是大部迁出型,性信息素诱日平均数量也显著高于花香诱捕的(P<0.05)(图1: C, D)。但四川江油的稻纵卷叶螟的生理状态和龄期则比较复杂,是田边草丛中的基本迁入型和稻田中的本地繁殖型混杂型,水稻发育期也不整齐,8月21-26日这一阶段性信息素诱捕日平均数量同花香诱的比较无显著差异(P=0.583)(图1: E)。

图1 各试验点花香与性信息素诱捕稻纵卷叶螟雄蛾的数量比较
2.3 田间网捕和花香诱的稻纵卷叶螟雌蛾卵巢发育及其交配状态
湖南攸县雌蛾为本地繁殖型,网捕雌蛾的卵巢发育级别较低,集中在Ⅰ级和Ⅱ级,花香诱的雌蛾则多为已经产过卵的Ⅳ级和产卵末期的Ⅴ级卵巢(图2: A)。Ⅰ级和Ⅱ级的卵巢没有交配,而Ⅲ级就有1次交配,花香诱到的雌蛾平均交配次数显著多于网捕的(P<0.05)(图3: A)。花香诱的Ⅴ级平均交配次数为3.5±1.0 次,最高5次,而网捕的平均为2.6±0.4次,最高3次。浙江宁波江北地区基本迁入型的花香诱捕的雌蛾Ⅴ级卵巢占比高达72%,而田间网捕的雌蛾Ⅴ级卵巢仅占3%,远低于花香诱捕的卵巢级别。网捕的雌蛾没有Ⅰ级卵巢(图2: B)。花香诱的Ⅱ级卵巢就有交配,Ⅲ-Ⅴ级交配次数接近,交配次数低于2次(图3: B)。广西田阳的雌蛾在8月23日至9月4日之间,也为基本迁入型。试验地在迁飞发生之前,稻田秧苗非常干净,没有稻纵卷叶螟幼虫。网捕和花香诱的雌蛾卵巢级别大致相同,都集中在Ⅳ级和Ⅴ级卵巢,网捕的雌蛾没有Ⅰ级卵巢(图2: C)。花香诱的Ⅱ级卵巢就有交配,网捕和花香诱捕的Ⅲ-Ⅴ级总交配次数接近(图3: C)。但同地在10月1-7日,网捕雌蛾卵巢级别集中在Ⅰ级和Ⅱ级。花香诱的雌蛾不多,但各级都有(图2: D)。交配次数只有在网捕极少数的Ⅴ级卵巢中有1次交配,因此是典型的迁出型。浙江宁波东吴在7月19-30日网捕迁入型的雌蛾Ⅰ级卵巢占比为4%。但花香诱捕的雌蛾中无Ⅰ级卵巢,Ⅴ级卵巢占比高达46.7%(图2: E)。花香诱到的雌蛾平均交配次数显著高于网捕(P<0.05)(图3: D)。同地在9月10-29日网捕雌蛾Ⅰ级卵巢占比达44%,但花香诱捕的Ⅰ级卵巢占比为0。Ⅴ级卵巢的比例网捕雌蛾仅有4%,但花香诱捕的却有80%(图2: F)。只有在Ⅳ级和Ⅴ级卵巢中才有交配,而且花香诱捕的雌蛾的交配次数显著高于田间网捕雌蛾的(P<0.05)(图3: E)。四川江油8月21-26日网捕和花香诱的雌蛾卵巢级别相差较大,花香诱捕的雌蛾Ⅴ级卵巢占比高达60%,而稻田网捕的雌蛾Ⅴ级卵巢仅占17%。草丛和稻田网捕有很大差异,显然稻田网捕的Ⅰ级和Ⅱ级占比要高,而Ⅴ级的占比极低(图2: G)。Ⅰ级和Ⅱ级卵巢没有发现有交配过的精包。其他级别的交配次数在1次左右,花香诱的交配次数稍高,但统计分析上不显著(图3: F)。

图2 各试验点网捕和花香诱稻纵卷叶螟雌蛾卵巢级别比较
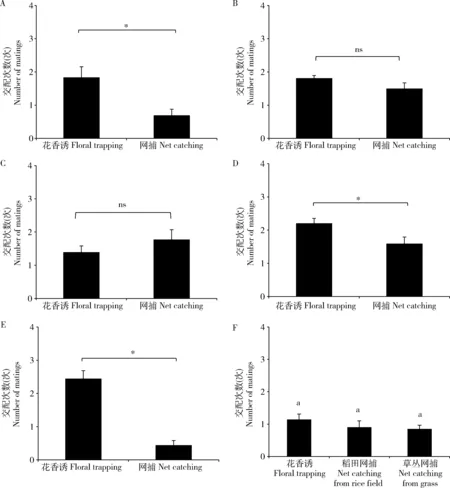
图3 各试验点网捕和花香诱稻纵卷叶螟雌蛾卵巢(Ⅲ-Ⅴ级)交配次数比较
2.4 田间网捕、性信息素诱和花香诱稻纵卷叶螟雄蛾精巢体积的比较
湖南攸县网捕雄蛾的精巢体积显著大于性信息素诱的(P=0.016)(图4: A)。广西田阳的网捕雄蛾精巢平均体积最大,且显著大于性信息素诱和花香诱的(P<0.05),但性信息素诱精巢体积和花香诱的精巢体积无显著性差异 (图4: B)。在四川江油稻田网捕雄蛾的精巢体积显著大于性信息素诱和花香诱的精巢的(P<0.001),性信息素诱精巢体积也显著大于花香诱精巢的(P=0.002),草丛网捕的雄蛾精巢显著小于稻田网捕的(P<0.001)(图4: C)。浙江宁波江北7月20-25日,网捕雄蛾的精巢体积最大,其次为性信息素诱精巢体积,花香诱精巢的最小,且三者存在显著差异(图4: D)。浙江宁波东吴7月19-30日网捕雄蛾的精巢体积最大,同性信息素诱的精巢体积无显著性差异(P=0.108),性信息素诱的雄蛾精巢体积显著大于花香诱的(P=0.002)(图4: E)。
2.5 性信息素诱与花香诱蛾的存活率和昼夜节律
每日清晨将诱捕器中的稻纵卷叶螟蛾类收集起来,记录存活数量,性信息素诱存活率可达82.5%,而花香诱蛾的存活率仅为31.5%。花香诱诱捕到的蛾死亡率较高,说明相对比较起来,花香诱捕的成虫年龄显然要大一些。花香诱发生在天黑之后、天亮之前的整个晚上,在19∶00-20∶00时为第一个高峰,天亮之前也有一个小高峰。但性信息素诱的时间不一样,性信息素诱捕的高峰时间在凌晨1∶00-06∶00时(图5)。

图5 性信息素诱和花香诱稻纵卷叶螟蛾的昼夜节律比较
2.6 性信息素诱和花香诱稻纵卷叶螟蛾量动态变化与田间下一代幼虫发育进程
将性信息素诱稻纵卷叶螟雄蛾量动态和花香诱蛾量动态对比,发现广西田阳地区试验点性信息素诱的峰期出现在8月26日,早于花香诱1 d,性信息素诱最高峰值出现在8月28日,花香诱的诱捕峰值出现在8月31日。性信息素诱的雄蛾数量均高于花香诱捕雄或雌蛾量,但花香诱雌蛾诱捕量是雄蛾的2倍多。花香诱捕在9月5日大幅度减少,9月15日以后诱捕量为零星。9月5-25日,性信息素诱蛾量远大于花香诱的,性信息素诱在9月30日-10月6日又出现另一峰值(图6: A)。田间种群调查显示,9月5日开始出现1龄幼虫,9月8日1龄幼虫到达高峰。2龄幼虫始见9月8日,9月11日到达高峰期,9月14日是2龄结束期。9月14日3龄幼虫到达高峰期,9月20日4龄、5龄幼虫到达高峰期。10月8日调查蛹量达到最高。性信息素诱始峰与2龄幼虫始见间距为13 d,性信息素诱最高峰值与2龄幼虫高峰期间距14 d。花香诱始峰与2龄幼虫始见间距为12 d,花香诱最高峰值与2龄高峰期间距11 d。
湖南攸县自9月5日起至10月1日,性信息素诱的雄蛾数量均远高于花香诱捕的雄蛾和雌蛾,性信息素诱的首个高峰出现在9月10日,花香诱的首个高峰出现在9月12日,但诱蛾量很低。相反,性信息素诱数量较大,峰形明显。性信息素诱雄蛾的高峰期在9月10日和9月15日。9月15日后田间性信息素诱蛾量逐日减少。10月2-11日,性信息素诱和花香诱蛾量都极低(图6: B)。湖南攸县地区田间调查显示,2龄幼虫的高峰期出现在9月24日,9月28日出现3龄幼虫的高峰期,4、5龄幼虫的高峰期出现在9月30日,9月30日至10月24日都能看到3, 4和5龄幼虫。性信息素诱最高峰值与2龄幼虫高峰期间距14 d,与3龄幼虫高峰期间距为18 d。花香诱最高峰值与2龄幼虫高峰期间距12 d,与3龄幼虫高峰期间距为16 d。

图6 2020年广西田阳(A)和湖南攸县(B)性信息素和花香诱稻纵卷叶螟蛾量的动态与田间调查幼虫种群密度之间的关系
广西田阳试验点,8月31日至9月16日花香诱雌蛾和网捕雌蛾的成熟卵巢(卵巢发育Ⅲ, Ⅳ和Ⅴ级之和)比例达90%以上,但10月份成熟卵巢比例极低,且花香诱诱捕的雌蛾极少(图7: A)。田阳地区花香诱捕的雌蛾出现两个高峰期,在8月29日-9月4日这一阶段中卵巢发育等级较高集中在Ⅳ级卵巢,即产卵盛期。但在10月1-7日时,雌蛾的卵巢等级都较低,大部分为不成熟卵巢,集中在Ⅱ级卵巢。
湖南攸县试验点,9月8-20日花香诱雌蛾的成熟卵巢比例较高,日解剖都在60%~100%以上,但网捕的雌蛾成熟卵巢的比例较低,日解剖卵巢都在57%以下。说明花香诱的雌蛾卵巢成熟度明显高于田间网捕雌蛾的(图7: B)。

图7 2020年广西田阳(A)和湖南攸县(B)网捕和花香诱稻纵卷叶螟雌蛾成熟卵巢比例随生理状态和季节的变化
3 讨 论
植食性昆虫羽化后的首要任务是求偶、交配,为下一世代找到理想的可以完成世代的生境和寄主,并成功产卵。这对于成虫寿命短的昆虫尤其如此。因此,昆虫为了适应各种气候和生境质量的可预测或不可预测的变化,进化出滞育和迁飞两种应对策略。这种适应性策略的差异导致有迁飞性和非迁飞性(本地虫源)之分,而迁飞性的稻纵卷叶螟种群有迁入虫源、本地繁殖扩散虫源和迁出虫源3种不同的迁飞状态及其混合种群(张孝羲等, 1981)。不同迁飞生理状态的稻纵卷叶螟成虫在生境和寄主选择、交配和产卵行为习性及其调控机制上也因此具有极大的差异(黄学飞等, 2010)。对于迁飞性昆虫来说,生理状态、幼虫寄主植物的营养状态、成虫的取食、环境中生物和非生物因子对其性信息素和花香气味的嗅觉反应影响会非常大。因此,花香气味、性信息素的行为调控与稻纵卷叶螟的生理状态密切相关。
稻纵卷叶螟成虫在迁飞过程中表现出“飞行与卵子发生共轭”,这意味着迁飞发生在生殖的未成熟阶段,迁飞的稻纵卷叶螟通常具有较长的生殖准备和产卵前期,雌蛾交配率和生殖力降低(Guoetal., 2019)。但是在许多情况下迁飞性昆虫的生殖和迁飞行为并不是绝对和非常清晰的,例如东方粘虫在迁飞期间可以有卵巢发育和交配(Zhaoetal., 2009)。交配过的小地老虎Agrotisipsilon也还可以远距离迁飞(Thomas and William, 1992)。从文献报道来分析,成虫繁殖与产卵依赖于幼虫和成虫期积累的资源,通常成虫寿命短、非迁飞性的昆虫,如果幼虫寄主营养合适的话,羽化的成虫不需要额外的蛋白和脂肪供应而直接交配、产卵,如二化螟Chilosuppressalis(冯波等, 2020)及毒蛾科和蚕蛾科的昆虫(Han and Gatehouse, 1991)。二化螟雌蛾基本上只有1次交配,而部分雄蛾可以有2~3次交配,所以,在田间花香诱捕器中没有二化螟雌蛾,但会有少量雄蛾(杜永均, 未发表数据)。迁飞性昆虫为了迁飞需要保存足够的能量,以甘油三酸酯形式的脂类存储为主。虽然通常人们认为,昆虫羽化后需要补充营养,但其实,昆虫羽化后更需要水(Gunn and Gatehouse, 1985)。补充营养对雌蛾繁殖能力的影响依害虫种类而有差异。 补充营养能够增加大豆夜蛾Anticarsiagemmatalis和烟芽夜蛾Heliothisvirescens的繁殖力和成虫寿命(Milanoetal., 2010)。水分、喂食糖和蜂蜜以及多次交配可以显著提高粉纹夜蛾Trichoplusiani雌蛾的繁殖力。烟芽夜蛾成虫补充营养得到的碳水化物还是卵生成或性信息素生物合成的前体(Fosteretal., 2014)。雌蛾能够获得水、食物和多次交配时,繁殖力最大,主要体现在产卵后期(Landolt, 1997)。稻纵卷叶螟仅喂食水的成虫卵巢发育水平显著低于提供补充营养的雌蛾(王凤英等, 2010)。稻纵卷叶螟的能量储备与迁飞状态有关,迁飞型的胸部存储的脂肪多于居留型,而迁入型则脂肪含量最低(张孝羲等, 1983)。
因此,虽然许多昆虫的某一特定时期与花之间存在密切的营养关系,特别是一些传粉昆虫。但昆虫是否能对花香气味起嗅觉行为反应,取决于昆虫的生理状态和需求。花香气味物质,特别是苯乙醛、苯乙醇、水杨酸甲酯等植物和花香挥发性化合物,引诱鳞翅目成虫的文献报道比较多(Meagher, 2002; Landoltetal., 2006; Meagher and Landolt, 2008, 2010; Landoltetal., 2014),还包括对许多传粉昆虫(Huberetal., 2005)、自然天敌(Rohrigetal., 2008)的引诱作用,花香气味对膜翅目、鞘翅目等其他昆虫,甚至有益生物的引诱作用也比较强(Meagher and Mitchell, 1999)。本研究比较了稻纵卷叶螟的不同迁飞类型对花香气味的嗅觉反应,结果显示,迁入型成虫对花香诱芯的反应比较强,可能与迁入型脂肪含量低,部分成虫需要寻找蜜源补充营养、延长寿命和尽可能多产卵有关。花香诱的诱捕峰型比较狭窄,在广西田阳试验点只有不足短短的10 d,可能是种群中只有部分成虫需要补充营养,这与其幼虫期的种群密度和营养状态有关。从花香诱捕雌蛾多于雄蛾、卵巢发育级别高、精巢体积稍小、雌雄死亡率高等指标可以推算,稻纵卷叶螟对补充营养的需求发生在产卵后期。
判断迁飞状态的3个重要指标就是卵巢发育为Ⅰ级的雌蛾百分率、交配率以及种群是上升还是下降。稻纵卷叶螟的交配行为已有详细观察(Hou and Chen, 1988)。温度是一个重要因素,在25℃下的交配率只有50%, 14.5℃就没有任何交配(Miyahara, 1981)。交配也与水稻植株的成熟期密切相关(Wada and Kobayashi, 1982)。稻纵卷叶螟可交配1~5次,一般是3次,雌雄蛾的多次交配提高产卵量和卵孵化率(Kawazuetal., 2014b),但交配虫龄的增加减低了繁殖能力(Kawazuetal., 2014a)。稻纵卷叶螟的成虫寿命长,因此,前后交配之间的时间间隔稍长。成虫性成熟时才释放性信息素,相对应的雄蛾也是这样。但与稻纵卷叶螟的迁飞性密切相关。本地繁殖扩散型(居留型)和迁飞的扩散型的雄蛾卵巢发育与性信息素诱关系存在线性相关性(冯波等, 2017a),但强迁飞型的稻纵卷叶螟,不管是南迁还是北迁,在迁飞前卵巢分级低,性发育不成熟,因此,相对应的雄蛾对性信息素没有嗅觉行为反应,田间性诱没有效果。迁入代在迁飞前或途中可能已经交配过,与下一次交配有一定的时间间隔,期间则有一定时间的交配静止期,需要一段时间后,才再次释放性信息素,在这之前,田间性诱也没有效果。
稻纵卷叶螟成虫寿命长,可以多次交配和产卵(Kawazuetal., 2014a),因此,以成虫龄期来推算产卵时间和卵期,其时间范围跨度较大,但如果以交配时间,也就是说,以性诱的诱蛾时间来计算,时间范围的跨度就比较狭窄。从广西田阳和湖南攸县2个试验点的性诱、花香诱捕和田间各龄幼虫的发育时间进程来比较、分析,结合张润杰和古德祥(1989)的温度与幼虫发育速率关系,除迁出型外,对于迁入型和本地繁殖型的稻纵卷叶螟,可以采用性诱来推算下一代的发育进程。性诱和花香诱之间的间隔时间基本上与产卵前期一致。计算不同温度下的各龄期时间,依据性诱和花香诱的始蛾期和高峰期,可以推算出各龄期的始期和高峰期,由此,可以比较精准地确定防治对策。迁出型的雌成虫卵巢发育还处于Ⅰ级,相应的雄蛾也处于性不成熟阶段,对性信息素没有嗅觉行为反应,性诱对迁出型没有引诱作用,但既然迁走了,所以,不影响对本地的发生和危害。因此,性诱其实可以很好地确定稻纵卷叶螟的迁飞类型,赶蛾、花香诱和灯诱无法确定稻纵卷叶螟的迁飞状态。稻纵卷叶螟的产卵前期在试验室饲养环境下只有1 d,依据张润杰和古德祥(1989)不同温度下的发育速率就可以推算出1-2龄期,由此可以确定打药时间,幼龄打药可以降低用药量,提高效果。
综上分析,不管是性诱还是花香诱稻纵卷叶螟,一个重要的前提是清楚田间成虫的生理状态。从本研究结果稻纵卷叶螟各迁飞状态的卵巢发育、精巢体积和成虫寿命分析可知,性诱成虫的虫龄较早,花香诱捕到的更大比例的是产卵接近尾声的雌蛾和交配结束的雄蛾。花香诱捕对部分迁入型成虫有效,迁入型和本地繁殖型的稻纵卷叶螟对性诱反应强烈,但前后2次交配之间有一段静止期,而对花香气味的反应则非常弱。迁出型的成虫雌蛾卵巢发育级别低,性发育不成熟,田间性诱和花香诱捕都没有效果。此外,花香诱捕在靶标害虫种类上并不专一,田间环境中存在的天敌或其他金龟子、蜜蜂和鳞翅目昆虫甚至二化螟雄蛾都可能进入,因此,可能破坏田间自然天敌系统。本研究结果可以提升我们利用和分析性诱数据,或与花香诱相结合,更准确预测稻纵卷叶螟的田间幼虫发生和迁飞。
