低鱼粉饲料中添加4种添加剂对花鲈生长性能、血清生化指标及养殖水体理化指标的影响
2022-04-13伏枥龙曹俊明王国霞朱喜锋赵红霞
■符 兵 伏枥龙 曹俊明 王国霞 彭 凯 朱喜锋 赵红霞 陈 冰*
(1.广东海洋大学,广东 湛江 524088;2.广东省农业科学院动物科学研究所,农业 农村部华南动物营养与饲料重点实验室,广东省畜禽育种与营养研究重点实验室,广东 广州 510640)
鱼粉作为优质饲料蛋白资源,因其必需氨基酸和脂肪酸含量高、适口性好、抗营养因子少、能够较好地被吸收消化等特点,已被广泛地应用于水产饲料中。但价格昂贵及资源有限,限制了其在水产中的应用。豆粕和花生粕因其来源广泛、价格低廉,且蛋白质含量高、氨基酸组成与动物接近,是最具有开发潜力的植物蛋白源。有研究发现,鱼类能耐受饲料中一定含量的植物蛋白,在适宜范围内不会显著影响水生动物的生长,也有一些鱼类耐受饲料中植物蛋白的剂量较高。但当饲料中只有植物蛋白时,会导致鱼类生长性能急剧下降。花鲈(Lateolabrax japonicas)又名海鲈、七星鲈,属于鲈形目、鮨科、花鲈属,为凶猛肉食性鱼类,鱼体蛋白质含量丰富,肉质鲜美,深受人们喜爱。由于花鲈具有生长速度快,对盐度和温度适应范围较广,无需室内越冬等特点,随着花鲈的集约化养殖规模程度逐渐提高,对于蛋白质饲料的需求量也逐渐增加。由于鱼粉的价格居高不下,因此提高花鲈对植物蛋白利用率,提高植物蛋白替代鱼粉水平,降低饲料成本以及节约鱼粉资源,是花鲈大规模养殖中亟待解决的重要课题。饲料中氨基酸的平衡程度是影响动物对其蛋白利用的关键因素。除了通过不同植物蛋白进行复合实现氨基酸的互补外,在水产饲料中往往也通过添加合成氨基酸以降低氨基酸不平衡所引起的负效应。核苷酸是生物体重要的组成成分,参与生物体内各种生长发育和代谢等重要过程如各种生物酶的产生与激活、遗传信息的传递和细胞信号转导,充足的核苷酸可为动物快速生长发育和发挥正常生理功能提供重要保障,酶制剂可提高饲料利用率,减少氮磷对水体的排放,补充内源性酶的不足和提高水生动物的免疫力。通过在饲料中添加诱食剂(核苷酸等)可提高饲料的适口性,添加外源酶制剂、益生菌等消除抗营养因子,添加必需氨基酸使其达到需求平衡从而提高鱼体对植物蛋白的利用率等。有研究表明,在饲料中添加核苷酸、酶制剂、中草药、益生菌等能有效改善水生动物对植物蛋白的吸收与利用。但关于在植物蛋白部分替代鱼粉的同时,添加赖氨酸和蛋氨酸、复合核苷酸、植酸酶及复合芽孢杆菌,改善饲料的氨基酸平衡、提高植物蛋白饲料适口性、降低植物蛋白的抗营养因子作用,进一步提高植物蛋白在花鲈饲料中的利用率,提高植物蛋白替代鱼粉水平还有待研究。因此,本试验以花鲈为研究对象,研究在低鱼粉(豆粕、花生粕部分替代鱼粉)饲料中添加4种添加剂(赖氨酸和蛋氨酸、复合核苷酸、植酸酶及复合芽孢杆菌)对花鲈生长性能、血清生化指标及养殖水体理化指标的影响,以期为植物蛋白在花鲈配合饲料中的应用提供理论依据。
1 材料和方法
1.1 试验饲料
包膜赖氨酸(L-lysine,纯度为40%)和包膜蛋氨酸(DL-met,纯度为50%)、晶体赖氨酸(L-lysine,纯度为78.8%)和晶体蛋氨酸(DL-met,纯度为99%)由广州某公司提供;枯草芽孢杆菌和地衣芽孢杆菌由湛江某公司提供,其中益生菌含量为5×109CFU/g;植酸酶(2 500 U/g)由广东某公司提供;复合核苷酸[腺苷酸二钠盐∶鸟苷酸二钠盐腺∶胞苷酸二钠盐∶尿苷酸二钠盐(AmpNa2∶GmpNa2∶CmpNa2∶UmpNa2)=1∶1∶1∶1,总核苷酸含量为62%]由南京某公司提供。以含28%鱼粉、30%豆粕和15%花生粕的饲料为基础饲料(G0组),在基础饲料中分别添加0.71%包膜赖氨酸和1.30%包膜蛋氨酸(G1 组)、0.06%复合核苷酸(G2 组)、0.04%植酸酶(G3 组)、0.10%复合芽孢杆菌(G4 组)配制4 种试验饲料,通过在G0、G2、G3 组和G4 组添加晶体赖氨酸和晶体蛋氨酸确保各组氨基酸含量一致,饲料的组成及营养水平见表1。原料经粉碎过60 目筛,采用逐级扩大法添加微量成分混合均匀后,使用SLX-80 双螺杆挤压机制成2.5 mm 的颗粒饲料,经55 ℃烘干,自然冷却后置于-20 ℃冰箱中保存。
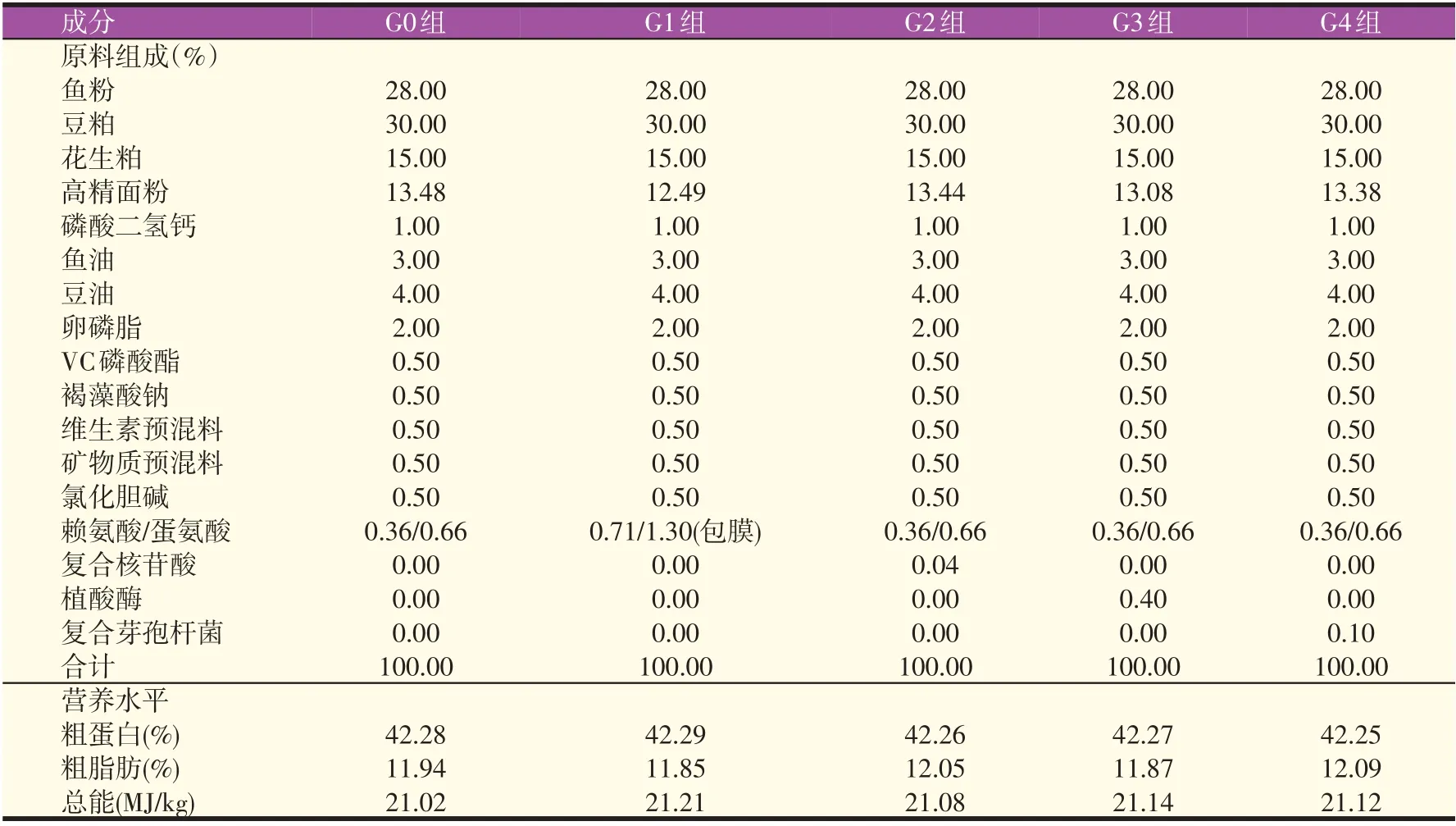
表1 饲料配方及营养水平(风干基础)
1.2 试验鱼与饲养管理
试验鱼购自福建诏安县英港育苗场,于3 m×2 m×1.2 m 水泥池暂养4 周,每天饱食投喂基础饲料2 次。养殖试验在广东省农业科学研究院动物科学研究所水产室室内循环养殖系统(玻璃纤维桶,体积约300 L)中进行,选取体重为(13.50±0.06)g健康、大小均匀的花鲈400 尾,随机分为5 组,每组4 个重复,每个重复20 尾鱼。分别投喂基础饲料和4种试验饲料。养殖水源为经过活性炭、珊瑚石过滤后的自来水,流水速率设置为5 L/min,养殖过程中不断充氧曝气,每天分别在09:00和18:00投喂饲料,投饲率为初始体重约4%~5%,养殖中期称重一次,调整投喂量,饲养周期为8周。饲养期间水温为27.5~32.5 ℃,溶氧量>6 mg/L,pH 7.3~7.6,氨氮<0.100 mg/L,亚硝酸盐<0.03 mg/L,自然光源。
1.3 样品采集
1.3.1 水质分析样品采集
在饲养试验末期,采用静水养殖方式进行试验,每个养殖桶水体体积保持一致,投喂相同重量的试验饲料,确保无残饵,不清理粪便,4 h 后进行中层水样的采集。采集的水样置于500 mL 的塑料瓶中,经过抽提过滤后于4 ℃保存,并在48 h内测定总磷、总氮、亚硝态氮、氨氮等指标。
1.3.2 血清和组织样品采集
饲养试验结束时,禁食24 h,称量终末体重(FBW),统计存活率(SR),计算试验花鲈增重率(WGR)、特定生长率(SGR)和饲料系数(FCR)。每个重复随机选取5尾鱼,测定体重、体长、内脏团和肝脏重,用于计算试验花鲈的形体指标;每个重复随机选取2尾鱼,用于全鱼常规营养成分的测定;每个重复随机选取6尾鱼,用1 mL无菌注射器于尾静脉取血,待斜放静置3~4 h后用冷冻离心机3 500 r/min离心10 min,将上清液分装于已做标记小管内,分装后置于-80 ℃冰箱保存备用。
1.4 指标测定
1.4.1 生长性能指标计算
增重率(WGR,%)=100×(终末体重+死亡鱼体重-初始体重)/初始体重
特定生长率(SGR,%/d)=100×(Ln终末体重-Ln初始体重)/饲养天数
饲料系数(FCR)=摄食量/(终末鱼体重+死亡鱼体重-初始鱼体重)
存活率(SR,%)=100×试验结束时鱼的尾数/开始时鱼的尾数
肝体比(HSI,%)=100×肝脏重(g)/体重(g)
脏体比(VSI,%)=100×内脏团重(g)/体重(g)
肥满度(CF,g/cm3)=100×体重(g)/[体长(cm)]3
1.4.2 常规营养成分分析用105 ℃烘箱干燥法测定饲料和全鱼的水分GB/T 6435—1986;用GB/T 6432—1994凯氏定氮法测定样品中含氮总量;使用GB/T 6433—1994索氏抽提法测定粗脂肪含量;采用GB/T 6438—1992测得灰分含量。能量值用IKA C200氧弹热量计测定。
1.4.3 血清生化指标分析
血清总蛋白(TP)、胆固醇(CHO)、血糖(GLU)、高密度脂蛋白(HDL)、低密度脂蛋白(LDL)、三酰甘油(TG)、谷草转氨酶(AST)、谷丙转氨酶(ALT)活性由广州金域医学检验中心采用日立7600全自动生化分析仪测定。抗氧化指标的测定均采用南京建成生物有限公司的试剂盒。
1.4.4 水质指标测定
水体总氮采用碱性过硫酸钾消解紫外分光光度法(GB/T 11894—1989)测定,氨氮采用纳氏试剂法(GB/T 7479—1987)测定,总磷采用钼酸铵分光光度法(GB/T 11893—1989)测定,亚硝态氮的测定采用萘乙二胺分光光度法(GB/T 5750.5—2006)测定。
1.5 数据统计
试验结果用“平均值±标准差(Mean±SD)”表示,采用SPSS 22.0 统计软件进行单因素方差分析(oneway ANOVA),利用Duncan’s 法进行组间显著性分析,差异显著性水平为P<0.05。
2 结果与分析
2.1 低鱼粉饲料中添加4 种添加剂对花鲈生长性能的影响(见表2)

表2 低鱼粉饲料中添加4种添加剂对花鲈生长性能的影响
由表2 可见,与G0 组相比,各添加组的终末体重、增重率和特定生长率均升高,且G2 组和G3 组达到显著水平(P<0.05),饲料系数G2 组和G3 组显著低于对照组(P<0.05)。各添加组花鲈的肝体比、肥满度、脏体比和存活率与G0 组相比没有达到显著水平(P>0.05)。
2.2 低鱼粉饲料中添加4 种添加剂对花鲈全鱼体成分的影响(见表3)

表3 低鱼粉饲料中添加4种添加剂对花鲈体成分的影响(湿重基础,%)
由表3 可知,各组间花鲈全鱼的粗蛋白、粗脂肪和粗灰分含量差异不显著(P>0.05)。G2组和G4组花鲈的水分显著高于G0、G1组和G3组(P<0.05)。
2.3 低鱼粉饲料中添加4 种添加剂对花鲈血清生化指标的影响(见表4)
由表4 可知,G3 组花鲈的血清总蛋白、白蛋白和高密度脂蛋白含量与对照组相比显著升高(P<0.05),其他添加组与G0组差异不显著(P>0.05)。相比G0组,各添加组花鲈的血清低密度脂蛋白、三酰甘油、血糖含量和谷丙转氨酶活性呈上升趋势,但未达到显著水平(P>0.05)。各组间花鲈的血清胆固醇、谷草转氨酶活性无显著性差异(P>0.05)。

表4 低鱼粉饲料中添加4种添加剂组合对花鲈血清生化指标的影响
2.4 低鱼粉饲料中添加4 种添加剂对花鲈血清免疫和抗氧化指标的影响(见表5)
如表5 所示,与G0 组相比,各添加组花鲈血清SOD 均升高,其中G2 组达到显著水平(P<0.05)。G1 组和G3 组花鲈血清CAT 活性与G0 组相比显著上升(P<0.05),G4 组显著下降(P<0.05)。各添加组花鲈血清T-AOC均比对照组高,其中G2组和G3组达到显著水平(P<0.05)。各组间花鲈血清ACP、AKP活性和HFB 水平无显著差异(P>0.05)。G4 组花鲈血清LZM活性显著高于G0组(P<0.05),G1组和G3组花鲈血清LZM 活性低于对照组,但差异不显著(P>0.05)。

表5 低鱼粉饲料中添加4种添加剂对花鲈血清免疫和抗氧化指标的影响(U/mL)
2.5 低鱼粉饲料中添加4 种添加剂对花鲈养殖水体理化指标的影响(见表6)
从表6可知,与G0相比,各添加组花鲈养殖水体总氮、亚硝态氮和氨氮含量均降低,但未达到显著水平(P>0.05)。各添加组花鲈养殖水体总磷含量均比对照组低,G3组达到显著水平(P<0.05)。

表6 低鱼粉饲料中添加4种添加剂组合对花鲈养殖水体理化指标的影响(mg/L)
3 讨论
3.1 低鱼粉饲料中添加4 种添加剂对花鲈生长性能的影响
本试验结果表明,低鱼粉饲料中添加包膜蛋氨酸+包膜赖氨酸、复合核苷酸、植酸酶和复合芽孢杆菌对花鲈生长性能的影响存在差异。在基础饲料中添加复合核苷酸和植酸酶能够显著提高花鲈的增重率、特定生长率,显著降低饲料系数。而添加包膜氨基酸和复合芽孢杆菌的作用效果接近。有研究结果表明,添加植酸酶能显著提高对斑点叉尾鮰(Ictalurus punctatus)、条纹鲈(Morone saxatilis)、奥尼罗非鱼(Oreochromis niloticus×O. aureus)、草鱼(Ctenopharyngodon idella)、花鲈(Lateo⁃labrax japonicus)等的生长性能。本试验研究结果显示,低鱼粉饲料中添加植酸酶可显著提高花鲈生长性能,降低饲料系数,同上述研究结果类似。其原因可能是植物中的磷主要存在形式以植酸及其盐类为主,难以被鱼体消化利用,而植酸酶可将植酸分解,提高植酸磷以及其他养分的利用率,从而促进花鲈生长。但也有研究表明,在饲料中添加植酸酶对黄颡鱼(Pel⁃teobagrus fulvidraco)的促生长作用不显著,这可能是饲料中添加植酸酶对机体的促生长机制和饲料系数的影响显著与否,与饲料组成、植酸含量以及养殖条件有关。有研究表明,饲料中添加核苷酸对大西洋鲑(Salmon salarL.)和苏氏芒鲶(Pangasius sutchi Flow⁃er)有显著的促生长作用。本试验结果与之相似。原因可能是核苷酸作为激素的介导,参与机体各种代谢过程,可促使细胞分化和发育。环核苷酸可以活化DNA聚合酶,使核糖体中蛋白质发生磷酸化,加速基因的转录和翻译,促进蛋白质的合成。而添加核苷酸对梭鲈(LuciopercaL.)、欧洲海鲈、虎龙杂交石斑鱼(Epi⁃nephelus fuscoguttatus♀×E. lanceolatus♂)、凡纳滨对虾(Penaeus vannamei)和草鱼的生长性能均无显著影响。一般认为添加包膜氨基酸可提高蛋白质的利用效率,促进机体生长。补充包膜赖氨酸和蛋氨酸可显著提高日本对虾(Marsupenaeus japonicus)的增重率和特定生长率,陈庭(2015)研究发现添加包被氨基酸可显著促进凡纳滨对虾的生长。王冠等(2006)发现,异育银鲫(Carassius auratus gibelio)饲料补充包膜氨基酸后,显著提高增重率,适当降低饲料中粗蛋白和鱼粉含量,节约蛋白质和鱼粉资源最终降低成本。王洪涛等(2009)通过补充微囊赖氨酸提高异育银鲫生长性能,钱前(2014)发现添加羟甲基纤维素包膜赖氨酸可提高湘云鲫的生长性能,而本试验添加包膜氨基酸组对于花鲈的增重率以及特定生长率影响不显著,可能的原因是花鲈肠道较短,包膜氨基酸在前肠还未得到全部释放就已经到达中场或后肠等部位,机体吸收的游离氨基酸不足以满足自身生长所需。必需氨基酸的缺乏会降低肌肉中饲料中氮的沉积,增加氨基酸的分解供能,对生长产生不利影响,且饲料中必需氨基酸的缺乏还会导致鱼体摄食量的降低。因此植物蛋白部分替代鱼粉时,赖氨酸和蛋氨酸等限制性氨基酸的补充非常重要,但对于水产动物,氨基酸平衡的作用效果与氨基酸的添加量、添加形式以及动物物种有关。本试验中,复合芽孢杆菌组对花鲈的促生长作用效果不显著。有研究表明,在凡纳滨对虾饲料中添加芽孢杆菌均可显著提高其增重率和特定生长率,其促生长机制可能主要和改善肠道菌群以及免疫和抗氧化功能有关,芽孢杆菌作为饲料添加剂可促进鱼类免疫和抗氧化功能的提高,进而提高抗病力并促进生长。本试验结果与之不同,可能在于本试验中添加的复合芽孢杆菌在肠道中未定植,在消化道绒毛壁上占据的位点不够。对照组生长性能最差的原因可能是花鲈为肉食性鱼,且本试验花鲈为幼鱼,需要更多的能量来满足花鲈生长,当饲料中植物蛋白替代鱼粉较高水平时,难以消化吸收的非淀粉多糖的含量增加,饲料中可消化能减少,导致增重率较低。
3.2 低鱼粉饲料中添加4 种添加剂对花鲈全鱼体成分的影响
本试验结果表明,低鱼粉饲料中分别添加包膜蛋氨酸+赖氨酸、复合核苷酸、植酸酶和复合芽孢杆对花鲈全鱼粗蛋白、粗灰分和粗脂肪含量没有显著影响。在饲料中添加包膜赖氨酸能显著提高尼罗罗非鱼体蛋白水平,显著降低粗脂肪水平,本试验结果与之不同。其可能的原因是在本试验中,包膜氨基酸未能解决氨基酸的平衡情况,鱼体蛋白周转能力较弱,而用于提供能量的蛋白量增加,导致了鱼体粗蛋白含量降低,脂肪升高,也可能是体内的矿物质以及营养元素因为植酸等抗营养因子的存在而利用率较低,导致饲料中缺乏矿物质元素,而这可能抑制了脂肪酸氧化过程,使鱼体粗脂肪含量较高。在日本对虾的研究中发现,饲料中同时补充赖氨酸和蛋氨酸对全虾粗脂肪和粗蛋白含量无显著影响,这可能与基础饲料中粗蛋白和粗脂肪水平有关。本试验结果表明,添加复合核苷酸显著升高花鲈全鱼水分含量,全鱼粗蛋白含量较对照组有升高趋势,全鱼粗脂肪含量呈下降趋势。在饲料中添加核苷酸后对锦鲤鱼体水分影响不显著,这可能与核苷酸的品质以及鱼的种类、饲养条件等的差异有关。在对凡纳滨对虾研究表明,饲料中添加核苷酸、核苷酸粗提物可提高凡纳滨对虾粗蛋白水平;在对异育银鲫研究表明,饲料中添加核苷酸鱼体粗蛋白含量显著提高,粗脂肪水平显著降低,本试验结果与之相似。可能原因是外源性核苷酸诱导一些代谢酶和激素的合成,促进蛋白质的合成代谢,减少脂肪积累。在本试验饲料中添加植酸酶对全鱼体成分的作用与核苷酸组类似,都是呈现粗蛋白和粗灰分上升,脂肪下降的趋势。郑涛等(2006)在奥尼罗非鱼饲料中添加植酸酶后显著提高鱼体粗蛋白水平,本试验结果与之类似。这说明植酸酶在水解植酸释放出饲料中磷的同时,破坏了与植酸螯合的蛋白质,激活了胰蛋白酶等酶的活力,提高了鱼体对蛋白质的利用。张璐等(2006)研究表明,在饲料中添加植酸酶后大黄鱼鱼体的粗灰分含量呈上升趋势,在草鱼饲料中添加植酸酶也得到类似研究结果,可能的原因是植酸酶能够解除植酸和矿物质之间的螯合作用,从而提高对矿物质的吸收利用率。本试验中,添加复合芽孢杆菌组也呈现全鱼粗蛋白水平上升、粗脂肪含量下降的现象。其可能的原因是芽孢杆菌可以产生许多消化酶,这些酶参与机体的代谢过程中,提高了蛋白质的积累。在凡纳滨对虾饲料中添加芽孢杆菌,虾体粗蛋白含量差异不显著,出现这些差异的原因可能是试验动物种类的不同所致。
3.3 低鱼粉饲料中添加4 种添加剂对花鲈血清生化指标以及免疫抗氧化指标的影响
血清生化指标可以反映机体生理健康状况,是疾病诊断的依据之一。谷丙转氨酶活性升高常常表明肝功能发生障碍;血糖含量的高低可以反映机体营养状况,含量较高时鱼体表现出较强的摄食能力,表明健康状态良好。本试验中,与对照组相比,各试验组谷丙转氨酶活性、谷草转氨酶活性以及血糖含量无显著差异,谷丙转氨酶有升高的趋势,但不代表影响鱼体肝脏等组织功能,而血糖的升高代表花鲈健康状态良好,这与在虹鳟(Oncorhynchus mykiss)、草鱼、异育银鲫上的研究结果相似。与对照组相比,植酸酶组血清总蛋白、白蛋白、高密度脂蛋白含量显著升高,说明植酸酶能使花鲈蛋白质和脂类代谢水平显著提高。各组间低密度脂蛋白、胆固醇差异不显著。血清免疫和抗氧化指标也能直接反映机体的氧化应激以及抗病能力,本试验中与对照组相比,各试验组血清超氧化物歧化酶活性均有升高趋势,植酸酶组花鲈血清过氧化氢酶活性、总抗氧化能力显著升高。在草鱼饲料中添加植酸酶能显著提高血清超氧化物歧化酶活性,凡纳滨对虾饲料中添加植酸酶显著降低血清碱性磷酸酶活性。本试验中,添加包膜氨基酸组血清过氧化氢酶活性与对照组相比显著上升,而血糖、谷丙转氨酶、谷草转氨酶以及超氧化物歧化酶活性与对照组相比有上升趋势,而碱性磷酸酶和酸性磷酸酶活性与对照组相比刚好相反,呈下降趋势。复合核苷酸组花鲈血清超氧化物歧化酶活性以及总抗氧化能力与对照组相比显著升高,且血糖、谷丙转氨酶、溶菌酶以及过氧化氢酶活性与对照组相比有上升趋势。向枭等(2011)发现,添加519~774 mg/kg核苷酸在鲤鱼饲料中能显著提高鲤鱼血清溶菌酶、超氧化物歧化酶活性。凡纳滨对虾饲料中添加核苷酸混合物显著提高血清超氧化物歧化酶活性以及总抗氧化能力。本试验结果与之一致,说明外源核苷酸可参与到机体免疫及抗氧化调控机制中,清除体内氧自由基,增强吞噬细胞的吞噬作用,提高机体的免疫及抗氧化能力。而在团头鲂(Megalobrama amblycephala)饲料中添加核苷酸对血清总抗氧化能力无显著影响。本试验中,与对照组相比,添加复合芽孢杆菌组花鲈血清过氧化氢酶活性显著下降,溶菌酶活性显著上升,血清总蛋白、血糖含量、超氧化物歧化酶活性和总抗氧化能力与对照组相比呈升高趋势。这与在牙鲆(Paralichthys olivaceus)、三角帆蚌(Hyriopsis cumin⁃gii)上的研究结果类似,可能的原因是芽孢杆菌通过作用于肠道集合淋巴结的抗原结合位点或通过动物体内的微生物群发挥作用进而提高机体免疫力。有研究发现,在南亚野鲮(Labeo rohita)饲料中添加芽孢杆菌可显著提高血清总蛋白含量,芽孢杆菌能提高金头海鲷(Sparus aurata)、凡纳滨对虾的免疫功能,芽孢杆菌提高免疫功能的机理一方面可能是芽孢杆菌在肠道内产生乳酸、乙酸等挥发性物质,降低了肠道pH,使得有害微生物的生长、繁殖、代谢受到抑制,间接提高了机体的免疫功能;另一方面,芽孢杆菌自身又能合成维生素以及许多辅酶,对机体的免疫起促进作用。
3.4 低鱼粉饲料中添加4 种添加剂对花鲈养殖水体理化指标的影响
低鱼粉饲料中分别添加包膜赖氨酸+包膜蛋氨酸、核苷酸混合物、植酸酶、复合芽孢杆菌组合对花鲈养殖水体的总磷、总氮、亚硝态氮以及氨氮含量均比对照组有不同程度的降低,这与杨艳玲等(2019)在花鲈中的研究结果一致。与对照组相比,添加植酸酶组水体总磷含量显著降低,而其他试验组与对照组相比差异不显著。饲料中植酸酶可以提高植酸磷的利用率,减少磷在水体里的排放从而降低水体总磷的含量。有研究表明,在凡纳滨对虾饲料中添加枯草芽孢杆菌能显著降低水体氨氮含量,抑制亚硝态氮的产生。在饲料中添加芽孢杆菌也得到类似的结论,可能是芽孢杆菌提高了消化酶活性,降低了氮排放量,且产生的酶内分解水体营养物质时将氨气、氮气等排出水体;另外芽孢杆菌可改变肠道和水体微生物的组成,如硝化-反硝化菌,可将氨氮和亚硝态氮转化成氮气,降低水体氨氮和亚硝态氮的含量。
4 结论
低鱼粉饲料中添加植酸酶和复合核苷酸,能显著提高花鲈的生长性能和免疫、抗氧化能力,增强花鲈对植物蛋白的利用率,其中以植酸酶的作用最明显。此外,添加植酸酶能显著降低花鲈养殖水体总磷含量。
