畜禽消化道健康定量评估五环标准之
——稳定的微生物区系
2022-04-13杨小军武圣儒曾祥芳刘作华王佳堃孔祥峰姚军虎
■杨小军 武圣儒 苏 勇 曾祥芳 刘作华 王佳堃 孔祥峰 姚军虎*
(1.西北农林科技大学动物科技学院,陕西 杨凌 712100;2.南京农业大学动物科技学院,江苏 南京 210095;3.中国农业大学动物科学技术学院,北京 100193;4.重庆市畜牧科学院,重庆 402460;5.浙江大学动物科学学院,浙江 杭州 310058;6.中国科学院亚热带农业生态研究所,湖南 长沙 410125)
消化道是营养物质消化吸收、免疫防御的关键场所,也是畜禽微生物的主要定植场所。消化道健康标准的建立对于评估畜禽机体健康尤为重要。对于畜禽而言,强有力的消化吸收、完整的物理屏障、特异性的化学屏障、稳定的微生物区系和适度的黏膜免疫可以有效地帮助宿主维持健康高效的生长、生产及繁殖。基于西北农林科技大学动物营养与健康养殖科研创新团队主持的重点研发计划相关研究以及近年来对畜禽消化道生理的研究,我们基于以上五个方向定义了畜禽消化道健康的五环标准。文章就稳定的微生物区系及其影响因素进行综述。
动物胃肠道内栖息着数量庞大、种类繁多的微生物群落,在漫长的自然选择和共同进化中与宿主形成了紧密的共生关系。以往肠道微生物的研究主要依赖于传统的分离培养方法,无法获取消化道菌群的全面信息。近年来,随着高通量测序技术的飞速发展,我们能够精细地表征肠道微生物基因组,并进行深入的分类学鉴定和功能分析,增加了对复杂微生态系统的认知。胃肠道微生物作为机体微生物群的重要组成部分,在参与动物生理代谢、免疫防护和疾病发展过程中扮演着重要角色。然而,微生物的结构组成与多样性受到宿主遗传和外界环境等因素的影响,其中,日粮组成、年龄以及宿主基因型可以显著改变畜禽肠道微生物构成。稳定的胃肠道微生物区系对抑制病原菌在肠道内定植、保证肠道健康、提高畜禽生产性能以及改善饲料利用效率等具有重要意义。
1 畜禽胃肠道微生物区系的构成与区室化
胃肠道微生物区系的建立是一个复杂的过程。具体表现为:①微生物利用宿主肠道中特定营养环境或依靠种内和种间的相互作用,形成具有一定稳定性的微生物区系。②微生物的群体感应可以对微生物的相互作用以及新加入的微生物进行重新调整,以形成稳定的微生物区系。动态的胃肠道微生物区系以功能的多样性和较大的适应性来应对外界环境的改变。因此,胃肠道微生物区系的建立能够加强微生物利用营养物质的能力,使微生物有效利用群体优势以应对营养供给变化。
动物所采食的饲料主要在胃肠道消化吸收。胃肠道的物理性消化、化学性消化以及微生物发酵在饲料的消化吸收过程中发挥了关键且互补的作用。胃肠道不同部位均定植了大量的微生物,但由于胃肠道的生物学位置以及功能的差异,胃肠道不同区域间细菌丰富度和多样性有所差异。
1.1 反刍动物胃肠道微生物构成
反刍动物各个肠段中均定植了大量微生物,由于不同的pH、肠蠕动、氧化还原电位、营养物质供应和宿主分泌物等导致胃肠道不同区域间细菌丰富度和多样性有所差异。瘤胃、小肠和大肠中的细菌在空间上彼此分离,但瘤胃和大肠的细菌群落多样性相似,而小肠样本中却与其有显著差异。
瘤胃微生物区系是一个多样化的生态系统,主要由细菌、原虫和真菌组成。其中,细菌是反刍动物体内数量最多、种类最丰富、代谢活性最强的微生物,在瘤胃中发酵植物性蛋白、多糖以及纤维素等,进而产生宿主维持和生长所必需的营养物质。同时,在瘤胃中进行大量复杂代谢活动的多种代谢物也被微生物用于自身增殖。研究发现,牛瘤胃中主要存在的厚壁菌门和拟杆菌门,各约占细菌总量的40%,且在饲料纤维素、半纤维素、淀粉、蛋白质和脂类物质的降解及其细菌代谢产物的生成中发挥重要作用。在属水平,普雷沃氏菌属、拟杆菌属、疣微菌属为优势菌属。此外,不同的日粮水平和饲喂方式也会导致宿主微生物组成的不同。
小肠是水和营养物质吸收的重要场所,其菌群分布与瘤胃大有不同。拟杆菌门在小肠中丰度较低,而变形菌门丰度较高。十二指肠中厚壁菌门为主要优势菌门,拟杆菌门种群的丰度仅为0.4%~1.1%。在属水平上,瘤胃中的优势菌属,如普雷沃氏菌属在小肠中几乎不存在,该属细菌在瘤胃中的高丰度可能与其高遗传可变性有关,使得其成员能够在瘤胃内占据各种生态位。然而,目前关于普雷沃氏属在小肠含量中较少的机制尚不清楚,有待进一步研究。厚壁菌门中优势的菌群为梭菌科和毛螺旋杆菌属,与十二指肠相比,空肠中的优势菌群中除梭菌科以外,芽孢杆菌也占较大比例。而回肠中,变形菌门(57.0%)丰度最高,其次是放线菌门(23.8%)和厚壁菌门(18.5%)。
与前肠道一样,微生物在胃肠道远端的代谢、生理和免疫过程中也至关重要。随着食糜向后肠道的移动,细菌的多样性发生变化。细菌以1012~1014个/mL的水平存在于后肠。反刍动物后肠道不同位置(盲肠、结肠、直肠)的微生物比较相似,主要为厚壁菌门(60%)和拟杆菌门(30%)。其中厚壁菌门中的优势菌为梭菌科(30%)、瘤胃球菌科(23%)和毛螺旋菌科(19%),拟杆菌门中的优势菌为拟杆菌科和普雷沃氏菌科。综上,反刍动物胃肠道微生物构成具有显著的“区室化”特征。
1.2 单胃动物肠道微生物构成
家禽消化道微生物多是通过外源定植而产生,其中以拟杆菌门、厚壁菌门和变形菌门为主要的优势菌群。由于特殊的消化道解剖结构,鸡肠道微生物的数量和多样性远低于牛、羊、猪等哺乳动物。家禽每个胃肠段因空间结构、内容物pH及生理功能等不同,导致不同肠段中的微生物组成和数量存在较大差异[21]。鸡嗉囊中定植着乳酸杆菌、肠球菌、链球菌、梭状芽孢杆菌等,其中乳酸杆菌属作为优势菌群,其丰度约占总菌量的28.62%,且主要参与了淀粉的降解和乳酸的产生过程。肌胃与腺胃紧密连接,菌群以乳酸杆菌、梭状芽孢杆菌和大肠杆菌等为主,但因肌胃的酸性环境,微生物丰度较低,发酵能力相对较弱。在小肠段中,十二指肠的细菌丰度最低。这可能是由于食糜通过时间短且消化物被肝脏分泌的胆汁稀释所导致。该肠段的微生物群落主要由梭状芽孢杆菌、链球菌、肠杆菌和乳酸菌组成。家禽空肠微生物区系研究相对较少,而回肠菌群研究较多。通过16S rRNA基因测序技术检测回肠微生物,发现乳酸杆菌为主要菌群(70%),其次是梭状芽孢杆菌(11%)、链球菌(6.5%)和肠球菌(6.5%)。
与回肠相比,盲肠拥有更丰富多样和稳定的微生物群落,且以厌氧菌为主。Oakley 等(2014)和Kogut等(2016)研究表明,商品肉鸡从孵化开始到6 周龄,盲肠微生物的丰富度和多样性显著增加,到3 周龄时,群落的分类组成由变形菌门、拟杆菌门和厚壁菌门迅速转变为厚壁菌门。然而,Kumar等(2018)研究发现,除盲肠第42 d 存在大量的拟杆菌门外,鸡的所有日龄(第0~42 d)的盲肠和回肠中均以厚壁菌门最为丰富。在属水平上,主要以瘤胃球菌属、乳杆菌属和梭菌属等厌氧菌为主。由此可见,家禽肠道微生物同样具有“区室化”分布的特点。
猪消化道中的微生物同样具有区域性,主要以厚壁菌门和拟杆菌门为主,且后肠道微生物数量显著高于前肠道。在菌群结构上,小肠中的优势菌主要为厚壁菌门的梭菌属、乳杆菌属和链球菌属;其中乳杆菌属和梭菌属为十二指肠—空肠段的优势菌;梭菌属和变形菌门的埃希氏菌属为回肠段的优势菌。后肠道中的优势菌主要为双歧杆菌属、瘤胃球菌属、链球菌属和乳杆菌属,其中瘤胃球菌属、普雷沃氏菌属和拟杆菌属为盲肠—结肠段的优势菌。可见,十二指肠—空肠微生物相似度较高,而盲肠—结肠微生物相似度较高。此外,猪肠道内还存在丁酸梭菌属、克里斯滕森氏菌属、颤杆菌属和棒杆菌属等,这些微生物可影响肥育猪的生长。
2 非健康肠道微生物区系
畜禽稳定的胃肠道微生物区系能够通过争夺致病菌的生存空间来抑制其生长。当肠道益生菌数量减少而病原菌数量增加时,其正常的生理平衡被打破,导致肠道菌群失调。
肠道微生物根据其与宿主的关系可分为共生菌、条件致病菌和致病菌三类。共生菌能够与动物保持良好的互利共生关系(如乳杆菌、双歧杆菌等),是肠道的优势菌群,具有营养及免疫调节作用。条件致病菌是保持动物肠道微生态平衡的必需组成部分(如肠球菌、肠杆菌等),正常情况下这些细菌不会致病,但在特定条件下具有侵袭性,对动物产生不利影响。致病菌长期定植机会少(如沙门氏菌、致病性大肠杆菌和变形杆菌、弯曲杆菌等),生理条件下数量极少,不会致病,但在机体抵抗力减弱时可以迅速增殖并引发肠道黏膜损伤等一系列有害影响。
2.1 致病菌及条件性致病菌感染对肠道菌群的影响
细菌感染会引起宿主发生免疫反应,这对清除肠道内病原菌(如沙门氏菌)是有益的,但对共生菌的生存却是不利的。沙门氏菌是一种常见的人畜共患食源性病原菌,可在肠道内定植并诱发早期肠道炎症反应。研究显示,感染沙门氏菌的肉鸡肠道微生物丰度随日龄的增加会发生明显变化,且与42日龄感染鸡相比,14日龄感染鸡肠道菌群变化程度最大,尤其是兼性厌氧菌和潜在病原体的增加使肠道菌群生态系统紊乱,导致肠道炎症发生。然而,沙门氏菌定植对肉鸡的生长一般没有明显的抑制作用,但诸如环境污染和食品安全等潜在问题仍然存在。弯曲杆菌感染的禽类盲肠微生物种类和数量也会发生改变,且该病原菌的入侵会持续在肠道内定植,进而产生促炎反应。最近一项关于致病性大肠杆菌攻毒试验的结果表明,致病性大肠杆菌诱发犊牛腹泻可降低肠道微生物的多样性,抑制抗菌物质的产生,破坏免疫系统。由此可知,细菌感染机体后导致肠道菌群失调,进而影响动物健康。
2.2 病毒感染对肠道菌群的影响
病毒感染宿主后可导致肠道菌群失调,不利于动物生长。据报道,鸡感染H9N2 亚型流感病毒后会提高回肠黏膜促炎细胞因子[如干扰素-α(IFN-α)、白细胞介素17A(IL-17A)、干扰素-γ(IFN-γ)和白细胞介素-22(IL-22)]的表达水平,并促进肠杆菌科的增殖和易位,尤其是大肠杆菌。这可能是由于肠上皮黏液层和紧密连接的完整性破坏而引起。小鼠感染流感病毒后,肠道微生物组成发生明显改变,其中肠杆菌和肠球菌数量明显增加,而乳酸杆菌和双歧杆菌数量明显减少。随着感染时间的延长,其与正常小鼠肠道菌群组成差异愈加显著。Mishra等(2018)分析发现,肠道菌群与免疫之间存在着平衡关系,流感病毒感染导致局部炎症反应,从而引起肠道菌群紊乱,降低机体对流感病毒的清除作用。就人类而言,儿童的肠道菌群稳定性较差,生物代谢速度快,更易受到病毒的感染,进而增加病原菌在肠道中的定植。老年人感染流感病毒后,由于肠屏障功能减弱以及自身免疫功能下降,肠道菌群容易出现质和量的变化,腐败菌占比显著提高,双歧杆菌减少,这也是导致老年人对各种疾病易感性增加、身体功能衰退的重要原因之一。
病毒感染猪同样会改变肠道菌群的组成和功能。如感染流行性腹泻病毒的猪肠道中梭菌和大肠埃希氏杆菌等致病菌丰度显著增加,而大多数共生菌嗜冷杆菌属、普雷沃氏菌属和粪杆菌属的丰度则减少。可见,肠道菌群、病原微生物和宿主三者相互作用,形成一个动态的平衡关系,在感染的进程中不断地共进化。
2.3 抗生素对肠道菌群的影响
肠道菌群失调除由病原微生物和宿主共生引起外,还由一些环境因素(如抗生素、日粮营养水平等)所导致。抗生素不仅可以促使病原菌入侵和繁殖,还会降低宿主原生菌群的丰度和多样性,对宿主产生长期的有害影响。研究显示,鼠伤寒沙门氏菌在健康小鼠肠道中很难定植,但仍可以增殖,使用抗生素处理后,肠道菌群被破坏,继而引发严重的肠道炎症。
不同抗生素的使用剂量持续时间和抗菌谱不同,对肠道菌群的作用也不尽相同。氟喹诺酮类抗生素是用于治疗家禽各种细菌感染的重要抗生素。有文献报道,高剂量恩诺沙星对鸡肠道沙门氏菌有良好的杀灭效果,但同时也会导致肠道菌群的结构和组成发生改变,如乳球菌和芽孢杆菌数量显著增加,肠球菌数量减少,且在停药期间明显富集,这可能是由于机体对抗生素产生耐药性的结果。也有研究认为,早期接触抗生素可引起仔猪肠道菌群失调,并显著降低免疫相关基因的表达。产蛋雏鸡饲喂抗生素对肠道菌群的丰度和分布产生负面影响,且抗生素干预后的菌群和免疫力很难完全恢复,结果进一步证实了抗生素引起的肠道菌群失调对畜禽免疫能力产生长期影响。肉鸡服用抗生素6 d,肠道菌群的组成和多样性受到显著影响,但服用16 d后,这种差异明显消失,表明抗生素的使用时间尤为关键。此外,短期(7 d)使用广谱抗生素(如克林霉素),可长期影响人体肠道菌群的丰度,且对微生物多样性产生不可逆的损伤。而对肠道菌群本身就不稳定的个体而言,抗生素对其影响更为复杂。因此,在畜禽养殖业中,寻找有效的抗生素替代品对维持肠道菌群稳态、保障动物健康至关重要。
综上,细菌、病毒感染以及抗生素滥用都会导致机体肠道微生物区系紊乱,具体表现为益生菌数量减少而有害菌数量增加,不利于动物生长,可定义为非健康的肠道微生物区系。因此,健康肠道微生物区系的必要条件是无肠道致病菌、条件性致病菌以及病毒的感染,且没有受到抗生素等不利因素的影响。
3 健康肠道微生物区系的形成
广义上定义,没有致病菌及病毒感染,同时没有受到抗生素等药物明显影响的微生物区系可定义为健康肠道微生物区系。健康的微生物区系具有良好的营养和免疫调节作用。
3.1 反刍动物健康肠道微生物区系及定植规律
人们普遍认为,新生反刍动物肠道处于无菌状态,出生后通过母体和环境获得的微生物很快定植于肠道。然而,近期的一项研究表明,羔羊在出生前,肠道就已经存在低多样性和低生物量的微生物组,且以变形菌门、放线菌门和厚壁菌门为主。可见,新生牛羊肠道是否有微生物定植目前存在争议,有待进一步深入研究。
反刍动物瘤胃中主要以厚壁菌门和拟杆菌门为主,其比例升高与动物体重增加有关。犊牛断奶后,随着高纤维高能量日粮的增加,瘤胃中普雷沃氏菌和拟杆菌的数量也会提高,前者可降解瘤胃中的非纤维性碳水化合物,同时也是主要的蛋白降解菌。厚壁菌门中存在分解纤维的菌属(如瘤胃球菌属、丁酸弧菌属等)。肠道中同样以厚壁菌门和拟杆菌门为主,但与瘤胃相比,拟杆菌门丰度下降,而变形菌门丰度提高。其中普雷沃氏菌、拟球梭菌和直肠真杆菌作为核心菌长久存在,而奇异菌属、柔嫩梭菌属以及一些益生菌属(如乳酸菌属、双歧杆菌属)的数量会随着宿主年龄的增长呈下降趋势。由于肠道各个部位承担的生理功能不同,因此在科水平上存在的主要微生物也不同。奶牛前肠道中毛螺旋菌科、消化链球菌科和肠杆菌科为核心菌,后肠道消化链球菌科丰度明显提高。瘤胃球菌是消化链球菌科中的菌属,在后肠道的丰度为24%,其作为混合发酵菌,可利用后肠道的纤维素和纤维二糖发酵为乙酸和甲酸。与前肠段相比,乳酸菌科的丰度升高,其作为肠道有益菌,可以抑制大肠杆菌和沙门氏菌的活性,进而减少犊牛腹泻的发生;提高机体的免疫水平,包括特异性免疫和非特异性免疫等。
3.2 单胃动物健康肠道微生物区系及定植规律
正常情况下,新生仔猪肠道是无菌的,随着与母猪产道、外界环境、粪便等的相互作用,多种微生物能够在肠道内按照一定顺序定植。然而,Liu 等(2019)在新生仔猪的胎粪中观察到丰富的微生物多样性,发现仔猪在母体子宫时,肠道就已存在微生物,并在出生一周后建立起稳定的微生物群落。也有研究表明,健康新生儿胎粪中的微生物不仅仅是在产后出现的。早产儿脐带血中存在许多可培养的微生物,羊水中也检测到大量不可培养和未分类的微生物。综上,新生仔猪或胎儿肠道微生物可能是在分娩前通过母源传播的,但这又与母体子宫基本无菌的结论存在冲突,因此,仍需要更多的研究来加以探讨。
仔猪出生2 d后,乳酸杆菌属、拟杆菌属等丰度逐渐增加,成为肠道内的优势菌群。14 d 后,小肠后端与结肠中的乳酸菌含量最为丰富。与断奶仔猪相比,乳酸杆菌数量在哺乳仔猪肠道中会更高,而大肠杆菌数量则相反。这归因于断奶仔猪胃内升高的pH导致乳酸杆菌减少,大肠杆菌快速增殖,从而抑制乳酸杆菌的生长。与母猪分离后,断奶仔猪的饲粮由液体的母乳换为固体饲料,其肠道生理和微生态环境发生巨大改变。随着肠道发育的不断完善,各肠段微生物组成等日渐成熟,“区室化”特征逐渐分明,所以成年猪肠道菌群的组成结构相对稳定。肠道中的核心微生物在营养物质吸收代谢、提高机体免疫力等方面发挥着不可或缺的作用。例如,粗纤维作为猪饲料中的主要成分,机体自身无法很好地分解利用,但后肠道中存在大量的纤维素降解菌(如瘤胃球菌),可以将粗纤维发酵成短链脂肪酸。乳杆菌在代谢过程中可以产生细菌素、有机酸、过氧化氢和短链脂肪酸,抑制病原菌增殖,同时也可帮助肠道中初级胆汁酸转化为次级胆汁酸,促进脂质的吸收等。猪肠道微生物组成受诸多因素的影响(如宿主遗传、环境、饮食、免疫力和抗生素等)。因此,通过调控肠道菌群结构来提高猪的生长效率和健康水平,具有非常广阔的应用前景。
鸡胚胎形成与哺乳动物的胎儿有所不同,虽然受精卵在母体内形成,但其胚胎发育在蛋壳内进行,与母体之间没有直接的联系。通过宏基因组测序技术对鸡胚中的肠道微生物区系进行检测,结果发现,鸡胚中存在丰富的微生物群落,其中变形菌门和盐单胞菌的丰度最为明显。进一步对3个不同发育阶段(鸡胚、雏鸡、母鸡)的肠道微生物进行对比,结果发现,65 个优势菌属在3个时期始终存在,暗示这些菌属在鸡的整个生命过程中扮演重要角色。
鸡肠道微生物的组成随年龄的变化发生短暂改变。高日龄比低日龄鸡盲肠微生物组成更复杂。1~3日龄雏鸡盲肠微生物种类较少,但菌群丰度及数量与成年鸡相比并无显著差异。孵化后7 d 内,盲肠微生物的变化较大,小肠微生物大概在14 d 内建立,14 d前回肠与盲肠的微生物结构相似,之后微生物结构及多样性存在明显差异。40 d 左右盲肠微生物建立并趋于稳定。总之,孵化后的早期阶段是鸡肠道微生物建立的关键时期。
3.3 健康肠道微生物区系与肉鸡生产性能、肠道形态结构及免疫指标的Meta分析
健康的肠道微生物区系能够与动物保持良好的互利共生关系。在畜禽养殖中,健康的肠道微生物区系还应该具有益生的调控效应。在此我们以肉鸡微生物区系为例,通过Meta分析初步研究了健康肠道微生物区系对生产性能、肠道形态结构及免疫指标的潜在作用。
3.3.1 文献检索与数据筛选
利用Science Direct、Web of Science、NCBI和中国知网(CNKI)等数据库筛选相关文献。关键词包括chicken、broiler、cecal microbiota、gut microbiota、intestine microbiota、cecum microbiota、villous height、crypt depth、feed conversion efficiency、feed conversion ratio、feed efficiency、residual feed intake 及immunity 的不同组合。文章采纳标准:试验动物为肉鸡;试验指标为盲肠/小肠微生物、小肠形态[包括绒毛高度、隐窝深度和绒毛高度隐窝深度的比值(V/C)]、饲料转化效率(FCR)和免疫因子A、G、M(IgA、IgG、IgM)。筛选结果见图1。

图1 文献筛选流程
3.3.2 结果与分析
3.3.2.1 盲肠微生物与饲料利用效率(FCR)
通过不同数据库检索获取的文章,大部分集中于粪杆菌属和乳杆菌属研究,且这两种菌属被认为是对机体有益生作用的优势菌属。因此,选用这两种菌属进行分析。
饲料利用效率(FCR)与盲肠粪杆菌属、乳杆菌属的回归关系见图2。Meta 分析结果表明,FCR 与粪杆菌属的回归方程系数大于0.05(P>0.05),未达到显著水平;FCR 与乳杆菌属呈显著负相关(P<0.05),达到显著水平。说明肉鸡盲肠乳杆菌属丰度的升高与FCR的降低有关,但与粪杆菌属之间并无显著关系。

图2 饲料利用效率(FCR)与盲肠粪杆菌属、乳杆菌属的回归关系
3.3.2.2 盲肠微生物与小肠形态结构
在盲肠/小肠微生物与肠道形态结构部分,检索获取的文章主要研究对象为大肠杆菌和乳杆菌属,其中,大肠杆菌是肠道中普遍存在的一种微生物,乳杆菌作为益生菌已被广泛应用于生产。因此,选用这两种菌进行Meta分析。
十二指肠上皮形态与盲肠乳杆菌属、大肠杆菌之间的回归关系见图3。Meta 分析结果表明,十二指肠绒毛高度与大肠杆菌、乳杆菌属的相关性不显著(P>0.05);隐窝深度与大肠杆菌呈先降低后升高的二次曲线变化(P<0.01),而与乳杆菌属呈先升高后降低的二次曲线变化(P<0.01);V/C 与大肠杆菌呈极显著正相关(P<0.01),与乳杆菌属丰度呈先降低后升高的二次曲线变化(P<0.01)。


图3 十二指肠上皮形态与盲肠乳杆菌属、大肠杆菌之间的回归关系
空肠上皮形态与盲肠大肠杆菌和乳杆菌属之间的回归关系见图4,Meta分析结果表明,空肠绒毛高度与大肠杆菌呈先升高后降低的二次曲线变化(P<0.01),与乳杆菌属之间无显著相关性(P>0.05);空肠隐窝深度与大肠杆菌、乳杆菌属都呈极显著负相关(P<0.01);V/C与大肠杆菌和乳杆菌属之间存在显著的正相关关系(P<0.01)。

图4 空肠上皮形态与盲肠乳杆菌属、大肠杆菌之间的回归关系
回肠上皮形态与盲肠大肠杆菌和乳杆菌属之间的回归关系见图5。Meta 分析结果表明,回肠绒毛高度与大肠杆菌丰度呈显著正相关(P<0.05),与乳杆菌属无显著相关性(P>0.05);回肠隐窝深度与大肠杆菌和乳杆菌属呈极显著负相关(P<0.01),V/C 与大肠杆菌和乳杆菌属呈极显著正相关(P<0.01)。

图5 回肠上皮形态与盲肠乳杆菌属、大肠杆菌之间的回归关系
3.3.2.3 小肠微生物及其形态结构
回肠绒毛高度与回肠大肠杆菌和乳杆菌属的回归关系见图6。Meta 分析结果表明,绒毛高度与大肠杆菌呈先升高后降低的二次曲线变化(P<0.01),与乳杆菌属之间存在极显著的正相关关系(P<0.01)。
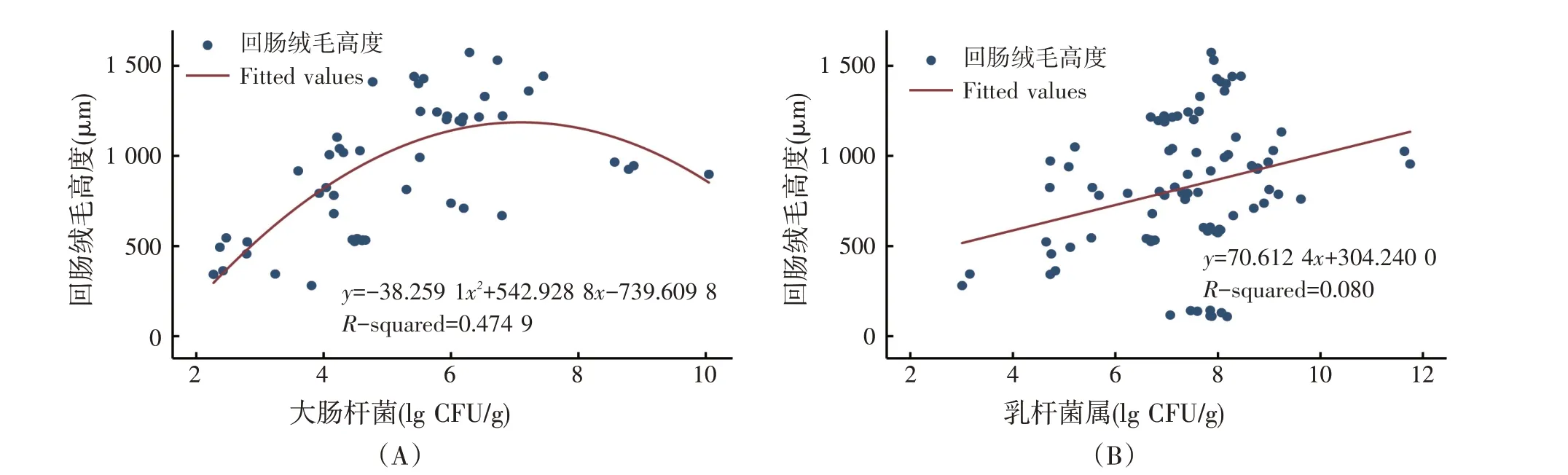
图6 回肠绒毛高度与回肠大肠杆菌和乳杆菌属之间的回归关系
回肠隐窝深度与回肠大肠杆菌和乳杆菌属的回归关系见图7。Meta 分析结果表明,隐窝深度与大肠杆菌、乳杆菌属的回归方程系数均未达到显著水平(P>0.05)。说明回肠的隐窝深度与大肠杆菌和乳杆菌属丰度之间无显著关系。
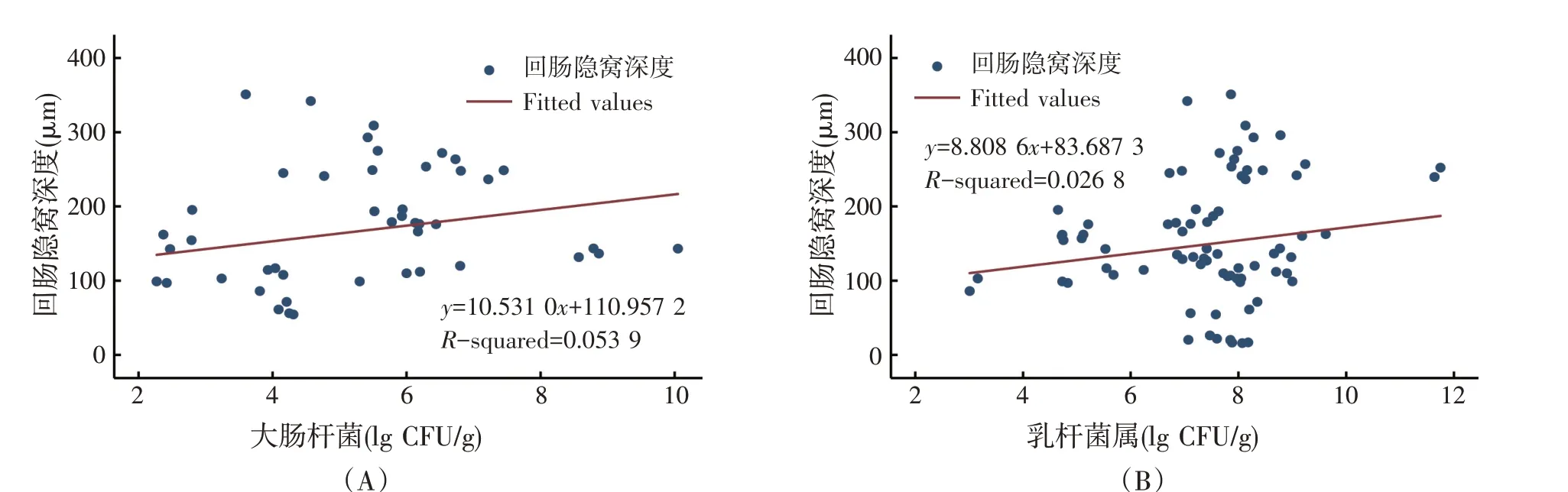
图7 回肠隐窝深度与回肠大肠杆菌和乳杆菌属之间的回归关系
回肠V/C 与回肠大肠杆菌和乳杆菌属的回归关系见图8。Meta分析结果表明,V/C与大肠杆菌、乳杆菌属的回归方程系数均未达到显著水平(P>0.05)。说明回肠的V/C与回肠大肠杆菌、乳杆菌属丰度之间无显著关系。
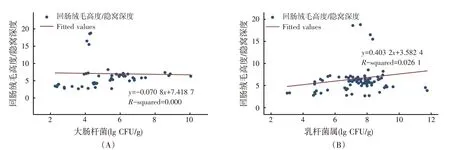
图8 回肠V/C与回肠大肠杆菌和乳杆菌属之间的回归关系
3.3.2.4 盲肠微生物与免疫因子
大肠杆菌是肠道中普遍存在的一种微生物,其菌群丰度可反映机体免疫水平和肠道健康状况,故此部分选用大肠杆菌作为分析对象。
肉鸡血清中免疫因子与盲肠微生物之间的回归关系见图9。Meta分析结果表明,血清IgA、IgG和IgM浓度与盲肠大肠杆菌属丰度呈显著负相关(P<0.01)。说明肉鸡血清中的免疫球蛋白随盲肠大肠杆菌丰度的增加而降低。
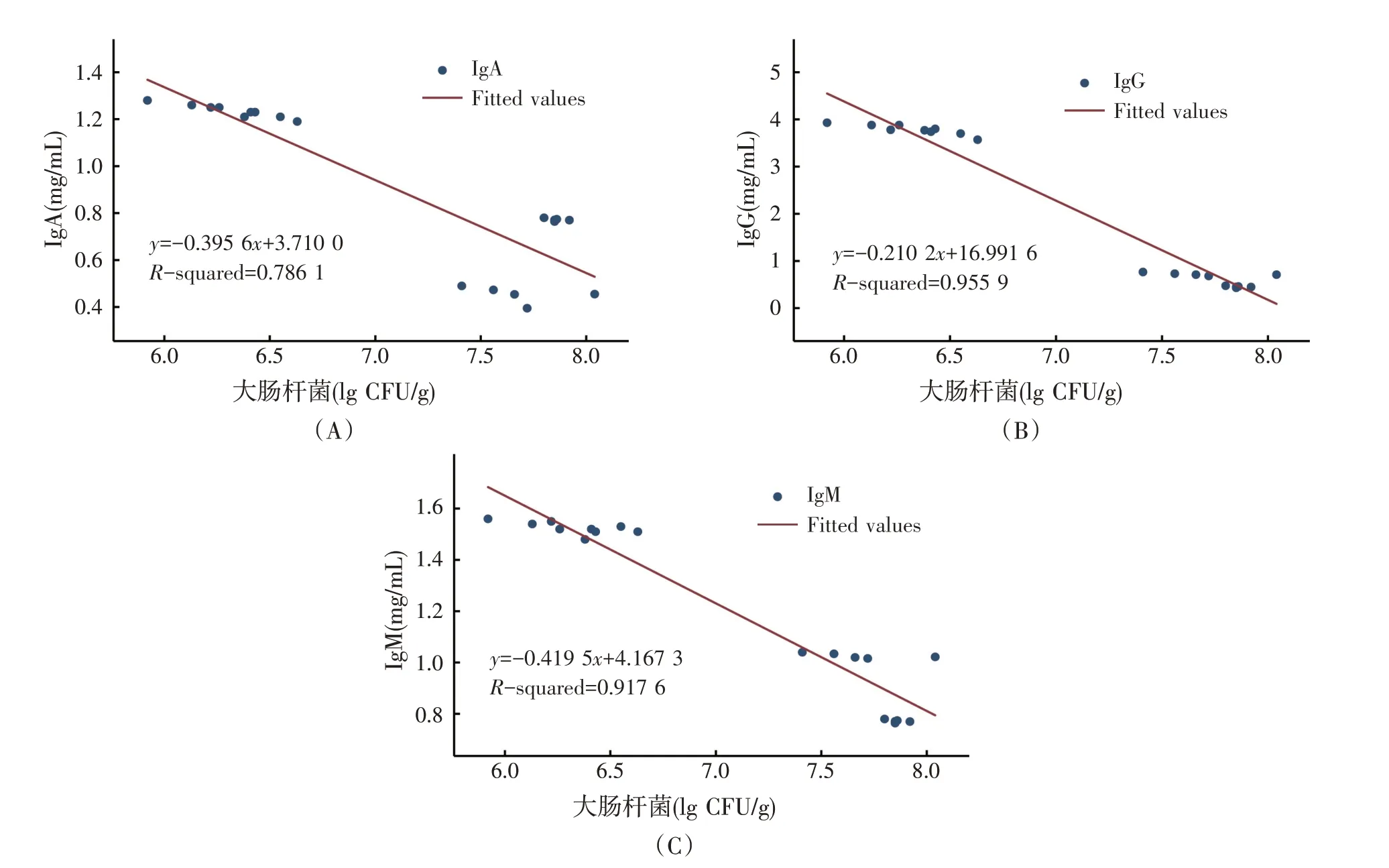
图9 免疫球蛋白与盲肠微生物之间的回归关系
4 讨论
健康的肠道菌群受宿主及环境双重因素影响下处于动态平衡,一旦失衡或紊乱则不利于畜禽肠道健康,甚至导致系统性代谢疾病的发生,最终导致其生产性能下降。在文章中通过Meta 分析证明,肠道微生物区系以及一些优势菌群的丰度变化与动物的生产性能、肠道结构形态和免疫反应息息相关。大量研究证实,健康的肠道菌群具有促进营养代谢、维持肠黏膜屏障、调节免疫系统、抑制病原菌感染、改善畜禽生产性能等多重功能。首先,微生物具有一些特异性的多糖利用位点,可以利用宿主无法直接利用的多糖,将其转化为宿主可以利用的短链脂肪酸等。且短链脂肪酸还可以为肠道细胞给养,从而维持上皮屏障功能,调控上皮增生,维持肠道免疫稳态及促进骨骼肌生长等,从而全方位地维护畜禽健康并促进其生长发育。其次,肠道微生物还具有蛋白质代谢功能,可以合成一些宿主自身无法合成的必需氨基酸,从而利于宿主的生长发育过程。特别地,后肠道菌群含有大量必需氨基酸合成相关的基因及基因簇,表明宿主与肠道菌群间氨基酸的交换是双向的。这种共生关系保证了宿主对某些必需氨基酸的摄入,同时通过后肠道发酵增加了无法通过宿主酶解吸收营养物质的利用效率。最后,肠道微生物合成的维生素K和部分维生素B,同样是宿主的正常代谢所必需的。综上,微生物的肠道给养功能所产生的短链脂肪酸等营养物质,不仅有利于肠道发育及免疫功能完善,还有利于动物的生长发育过程。
肠道菌群与宿主的免疫反应相关。稳定的微生物区系可以阻止病原菌繁殖,对宿主肠道屏障起到保护作用。与正常小鼠相比,无菌或肠道菌群多样性较低的小鼠存在许多免疫缺陷,对肠道病原菌的易感性更高,说明健康的胃肠道微生物区系在宿主免疫成熟与调控过程中发挥着重要作用。此外,肠腔内特定的共生细菌能够促进T 细胞[如调节性T 细胞(Treg)、辅助性T细胞(Th17)]不同亚群的生成,维持肠道在免疫耐受与炎症反应之间的平衡,有利于肠黏膜稳态的建立。另外,肠道菌群的代谢产物[如胆汁酸、维生素、短链脂肪酸(SCFAs)、多糖(PSA)、共轭脂肪酸等]可以直接作用于肠道或通过体液循环,影响肠道及其他组织的免疫调控,最终将肠道菌群与全身免疫系统有机联系在一起。由此可见,肠道菌群的定植与代谢刺激了宿主免疫系统的发育与成熟,宿主免疫系统的不断发育成熟则反过来影响着肠道菌群的组成结构与功能等。
5 小结与展望
畜禽消化道微生物区系的平衡是保障宿主健康状况和发挥正常生产性能的必要条件。文章Meta 分析结果表明,肠道微生物的改变对动物的生产性能、肠道形态结构和免疫反应产生不同影响。可见,保证胃肠道优势菌群的稳定在维持机体健康、促进肠道发育和营养物质吸收方面至关重要。胃肠道微生物的区室化及其功能的差异,导致其在畜禽的胃肠道发育、生长及免疫调控中发挥着不同的作用。随着高通量测序技术的发展,我们应运用系统思维看待胃肠道微生物区系,即“消化道一体化”,将有助于业界深入探讨微生物菌群和各种调控因子间的协同作用,对提高动物生产性能和改善饲料利用效率具有重要意义。
