温室黄瓜连作土壤真菌多样性变化分析
2022-04-08孙玉禄高晓梅刘晓辉桓明辉池景良
李 杨,敖 静,孙玉禄,高晓梅,刘晓辉,桓明辉,池景良
(辽宁省微生物科学研究院,辽宁省微生物菌种选育重点实验室,辽宁 朝阳 122000)
黄瓜(Cucumis Sativas L.)是我国设施园艺生产的主要蔬菜作物,栽培面积占蔬菜总面积的60%,随着设施栽种面积的逐年增加,为追求高产超量施用农药和化肥,造成土壤理化条件改变、健康状况恶化、病虫害增多、产量下降、连作障碍严重,严重影响设施农业的可持续发展,同时食品安全也威胁着消费者的健康,因此黄瓜连作障碍问题越来越多地引发人们关注[1]。
土壤微生物是农业生态系统中的重要组成部分,土壤真菌在土壤中的数量仅次于细菌,承担着土壤分解者的角色,在物质循环和能量循环过程中发挥着重要作用,可以将复杂难降解的有机质转化为植物可以利用的营养元素[2-5]。有研究表明,作物连作栽培定向地影响土壤微生物的生长繁殖,细菌和有益微生物数量减少,病原真菌数量增加[6],导致土壤根际微生态系统失衡,是土壤连作障碍的主要原因[7-8]。
传统研究方法应用于微生物群落的多样性已取得较多的科研成果,但由于土壤中绝大部分微生物是不可培养的,传统的分离鉴定方法只能区分上述种类中的0.1%~10%,导致对环境微生物群落多样性的认识受到限制[9-11]。随着分子生物学技术的突破,新一代高通量测序技术为研究微生物丰度、组成、群落结构、遗传进化、代谢途径等提供大量的信息,为大规模、快速、准确、全面检测土壤微生物多样性提供了技术保障[12-13],使微生物和连作障碍之间的关系再次成为研究的热点。本试验采用Illumina MiSeq 高通量测序技术,以日光温室黄瓜连作不同年限土壤为研究对象,分析温室黄瓜不同连作年限根际土壤样品真菌种群结构和多样性的变化,获取了真菌在门、纲、目、科、属不同分类水平上的相对丰度、群落结构和遗传信息,探索了连作模式下土壤真菌中有益和有害菌群消长和演替的响应,阐明了连作年限-土壤微生物群落结构-环境因子之间的相互关系,为克服连作障碍提供理论依据,助力设施农业可持续发展。
1 材料与方法
1.1 试验地概况及试验设计
本试验样品来自辽宁省微生物科学研究院喀左平房子镇设施蔬菜科研基地,位于119°38′39′E、40°58′44′′N,海拔310 m。试验设4 个处理,根据种植年限分别命名为连作1 年(F1)、3 年(F3)、5 年(F5)和7 年(F7),每个处理设3 次重复,编号分别为F1A、F1B、F1C;F3A、F3B、F3C;F5A、F5B、F5C;F7A、F7B、F7C,共12 个样品,田间管理措施相同。
1.2 样品采集
在黄瓜种植后期,去除表层砂石和腐叶,每个处理在深度为20 ~30 cm 处采集3 份土壤,在每个区采用五点取样法取样,混匀成1 份装进自封袋放入冰盒带回实验室。一部分鲜土放入超低温冰箱,用于微生物多样性研究;另一部分鲜土放入-4℃冰箱保存,用于测定可培养微生物数量和酶活性;还有一部分土壤自然风干后过1 mm 筛,用于测定土壤基本理化性质。
1.3 土壤基本理化性质测定
采用ThermoA211 酸度计按水土比5∶1 测定土壤pH;采用ORIONTDS 型电导率仪测定EC 值;采用碱解扩散法测定碱解氮;采用盐酸-硫酸混合液浸提,北京吉天全自动流动注射分析仪iFIA7 测定土壤有效磷;采用醋酸铵浸提-火焰光度法测定速效钾。
1.4 土壤微生物总DNA 的提取
采用Omega soil DNA kit 试剂盒,称取0.5 g 鲜土,根据说明按步骤提取12 个土壤样品中的微生物基因组DNA,0.8%的琼脂糖凝胶电泳检测DNA质量并通过紫外分光光度计对DNA 进行定量。
1.5 真菌ITS1 区PCR 扩增及测序
采 用 特 异 性 引 物ITS5F:5’-GGAAGTAAAA GTCGTAACAAGG-3’、ITS1R:5’-GCTGCGTTC TTCATCGATGC-3’对 真 菌ITS rRNA 的ITS1 区 进行PCR 扩增。PCR 反应体系为Q5 DNA 聚合酶0.25 μL,5×扩增缓冲液5 μL,5×GC 缓冲液5 μL,10 mmol/L dNTP 0.5 μL,模板DNA 1 μL,10 μmol/L正向、反向引物各1 μL,水11.25 μL。PCR 扩增条件为:98℃预变性2 min;98℃15 s,50℃30 s,72℃30 s,循环25 ~27 次;72℃延伸5 min。PCR产物采用2%琼脂糖凝胶电泳检测,用Axy Prep DNA 凝胶回收试剂盒回收目的片段,样品送上海派森诺生物科技有限公司进行Illumina MiSeq 高通量测序。
1.6 序列获取与分析
为获得更为精准的生物学信息,首先使用Fast QC 进行原始数据的质量控制,对测序原始数据进行拼接、过滤,获得最终用于分析的序列。使用Usearch 在0.97 的相似度下进行去冗余、聚类和操作分类单元OTU 表格生成。采用Graphpad Prism 8 one-way ANOVA 方法进行差异显著性分析。利用Mothur[14]计算真菌群落的α 多样性,并对Shannon 指数和Chao1 指数进行分析。基于Bray_Curtis距离矩阵,采用主坐标分析(PCoA)进行真菌群落的群落结构和聚类分析。利用FUNGuild 数据库分析真菌的功能类别,进行功能预测和分类[15]。选择Pearson 分析土壤真菌门相对丰度与土壤理化性质的相关性,采用Mantel 检验分析真菌整体群落与土壤理化性质之间的相关性。
2 结果与分析
2.1 温室黄瓜连作不同年限根际土壤真菌相对丰度和群落组成的变化
通过Illumina Miseq 高通量测序分析,从12个样品中共获得786148 条优化序列,每样本为46932 ~91804 条优化序列(平均65512 条优化序列),213009495 优化碱基数,优化序列长度50 ~485 bp,平均长度270 bp(表1),共检测到14 个门,42 个纲,90 个目,197 个科,318 个属和1205 个OTU。连作不同年限后真菌群落组成在门和纲分类水平上发生变化(图1、2)。在门水平上,子囊菌门(Ascomycota)为优势菌门,相对丰度占55.74% ~77.78%,其次是未被归类的真菌(8.04% ~23.38%)、担子菌门(Basidiomycota)(3.59% ~19.63%) 和Mortierellomycota(0.20% ~6.49%), 壶 菌 门(Chytridiomycota)(0.08% ~1.82%)、Aphelidiomycota(0.01%~1.57%)和Rozellomycota(0.51%~1.12%)相对丰度占比很少,且在F1 中并未发现Rozellomycota 真菌门(图1a)。日光温室黄瓜连作不同年限后真菌在门水平上的群落组成变化明显,子囊菌门相对丰度在F3 样品中无明显变化,在F5 和F7 样品中相对丰度分别下降9.94%和28.24%,而担子菌门相对丰度在F3 样品中显著降低后又逐年上升。
在纲水平上,粪壳菌纲(Sordariomycetes,28.01% ~68.01%)、c_unclassified_k_Fungi(8.04% ~23.38%)、散囊菌纲(Eurotiomycetes,1.75% ~22.81%)、 伞 菌 纲(Agaricomycetes,1.13%~13.64%)和座囊菌纲(Dothideomycetes,2.27%~10.35%)的相对丰度较高,其次是c_unclassified_p_Ascomycota(0.18% ~6.15%)、 银耳纲(Tremellomycetes,1.05% ~4.40%)、被孢霉 纲(Mortierellomycetes,0.20% ~6.12%)、盘菌纲(Pezizomycetes,0.13%~5.0%)、酵母菌纲(Saccharomycetes,0.09% ~1.88%) 等( 图1b)。粪壳菌纲相对丰度变化显著,连作3 年样本中相对丰度最高,达68.01%,之后持续下降,到连作7年时剩余28.01%。

表1 真菌ITS1 Illumina MiSeq 高通量测序结果

图1 黄瓜连作条件下土壤真菌在门和纲分类水平上的群落组成
LEfSe 分析显示,在属分类水平上,设施黄瓜连作1 年后起重要作用的属有Ramophialophora、锥盖伞属(Conocybe)、裂壳菌属(Schizothecium)、柄孢壳菌属(Podospora)和微囊菌属(Microascus),连作3 年后起重要作用的属有粪盘菌属(Ascobolus)、蜡蚧菌属(Lecanicillium)和被孢霉属(Mortierella),连作5 年后起重要作用的属有Pseudopithomyces、拟青霉属(Simplicillium)和嗜热真菌属(Thermomyces),连作7 年后起重要作用的属有曲霉属(Aspergillus)和栓菌属(Trametes)(图2)。
2.2 不同连作年限对温室黄瓜根际土壤真菌群落多样性的影响
Chao1 指数主要是计算群落丰度,用于估计群落样品中包含的物种总数,Chao1 指数越大,微生物群落物种数越大。Shannon 指数主要用于计算群落多样性,该指数受群落中物种丰富度和均匀度的共同影响,Shannon 指数越大,微生物群落的多样性越高。日光温室黄瓜长期连作增加了土壤中真菌的多样性,Shannon 指数随着连作年限的增加逐渐增高,但差异不显著(图3a);Chao1 指数在连作3年达到最高,5 年后明显降低(图3b),且差异显著(P<0.05)。真菌群落的多样性为F7>F5>F3>F1,而真菌总数为F3>F1>F7>F5。

图2 Lefse 分析连作不同年限真菌组成差异

图3 设施黄瓜不同连作年限土壤真菌多样性指数组间差异检验
对黄瓜连作不同年限的真菌群落整体进行主坐标分析(PCoA),结果(图4)显示黄瓜连作年限可以影响真菌群落结构。不同连作年限黄瓜根际真菌群落结构均差异极显著(P<0.01),在PCoA 上4种连作年限土壤样品分开,相同年限土壤样品聚在一起。基于Bray-Curtis 的ANOSIM 进一步分析表明,在不同连作年限之间的真菌群落结构的差异明显高于组内差异(图5)。
2.3 温室黄瓜不同连作年限对土壤功能真菌的影响
采用FUNGuild 对不同连作年限真菌群落进行功能预测,4 组样品共检测到1205 个OTU,其中479 个OTU 被划分为8 个真菌功能类群,约占总OTU 数量的39.75%。从图6 中可以看出,腐生菌(Saprotroph)是主要功能类群,包括240 个OTU,占总OTU 数量的19.92%。然后为植物病原菌/腐生/共生过渡型菌(Pathotroph-Saprotroph-Symbiotroph),包括75 个OTU,约占6.22%;植物病原菌(Pathotroph), 包 括50 个OTU, 约 占4.15%;腐生/共生过渡型菌(Saprotroph-Symbiotroph),包括40 个OTU,约占3.32%;植物病原菌/腐生菌过渡型(Pathotroph-Saprotroph),包括36 个OTU,约占2.99%。共生菌(Symbiotroph)仅有19 个OTU,占1.58%;植物病原菌/共生过渡型菌(Pathotroph-Symbiotroph),12 个OTU,约占1.0%;植物病原/腐生/共生过渡型菌(Pathogen-Saprotroph-Symbiotroph)只有7 个OTU,所占比例为0.58%。本研究中仍有60.25%的OTU 不能进行功能分类,需要进一步进行分析。
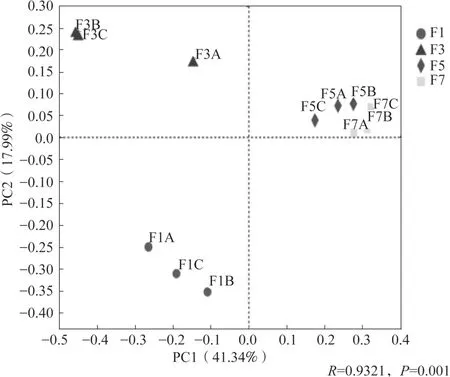
图4 基于属水平的Bray-Curtis 距离矩阵不同连作年限土壤真菌群落的主坐标分析
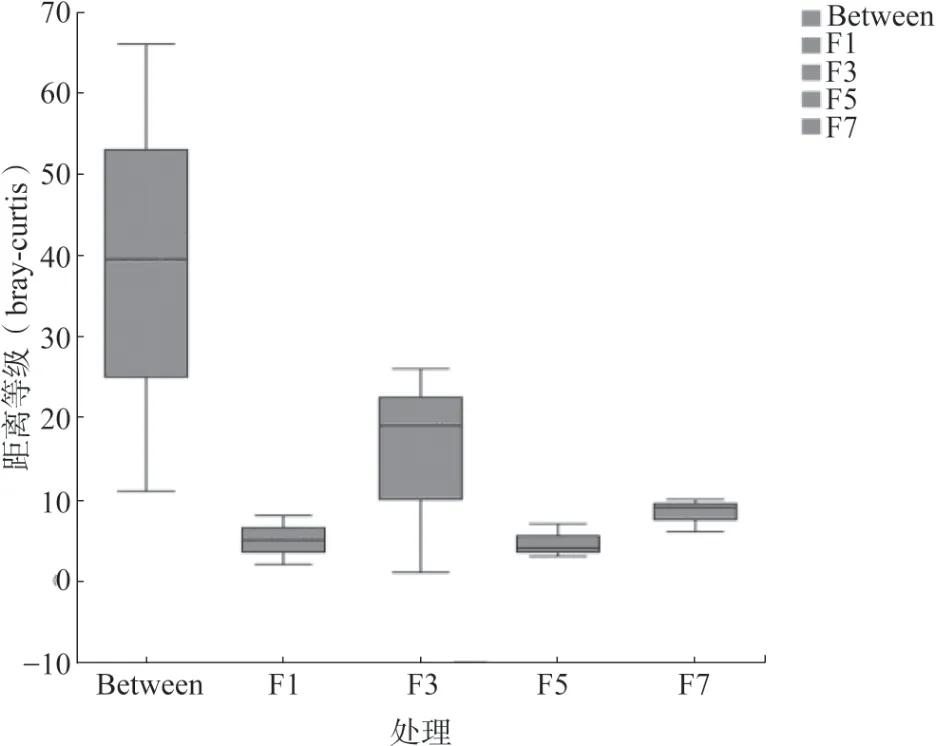
图5 基于Bray-Curtis 距离矩阵ANOSIM 差异分析
从图6 可以看出,设施黄瓜连作不同年限根际土壤中真菌功能类群的相对丰度不同。其中,连作3年(F3)根际土壤中的腐生/共生过渡型菌(6.76%)和植物病原菌/腐生/共生过渡型菌(44.75%)的相对丰度明显高于F1、F5 和F7(0.94%~2.42%、12.15%~14.76%),连作5 年土壤中植物病原菌/腐生/共生过渡型菌(0.91%)、植物病原菌/共生过渡型菌(18.22%)和植物病原菌(8.70%)的相对丰度最高,而共生菌(1.69%)和腐生菌(35.45%)的相对丰度在连作7 年样品中最高。但是仍有很大一部分真菌不能进行功能归类(Unassigned)。
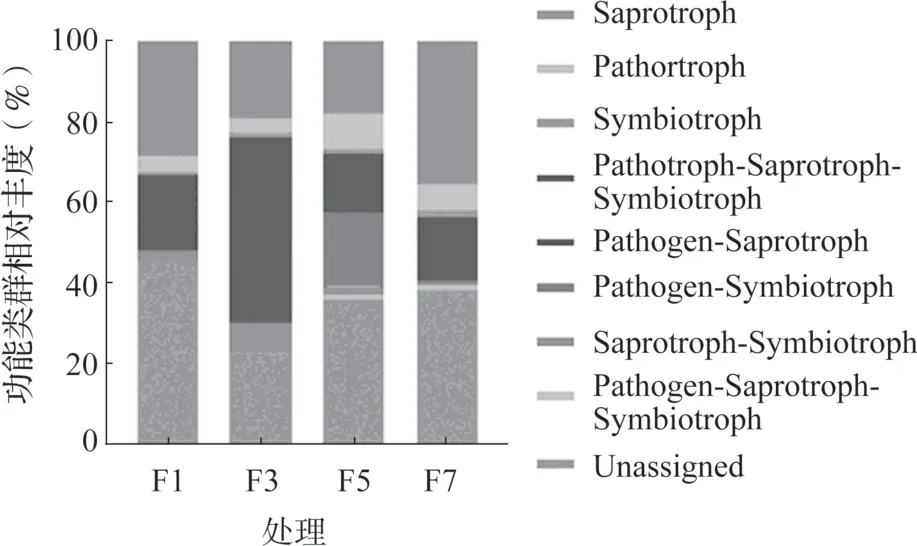
图6 不同连作年限黄瓜根际真菌菌群功能组成与丰度
2.4 土壤真菌群落与土壤理化性质间的相关分析
土壤环境因子pH、碱解氮(AN)、脲酶(SUR)等与土壤真菌群落结构之间存在密切关系。如图7所示,土壤pH 与Aphelidiomycota(R=-0.651)和毛霉门(Mucoromycota)(R=-0.704)呈显著负相关关系(P<0.05)。土 壤AN 和SUR 与Mortierellomycota(R=0.741,R=0.713)呈极显著正相关关系(P<0.01),与梳霉门(Kickxellomycota)(R=0.65,R=0.591)呈显著正相关关系(P<0.05),与担子菌门(Basidiomycota)(R=-0.762,R=-0.762)呈极显著负相关关系(P<0.01);土壤AN 与油壶菌门(Olpidiomycota)(R=0.708)呈极显著正相关关系(P<0.01),SUR 与捕虫霉门(Zoopagomycota)(R=0.78)呈极显著正相关关系(P<0.01),与油壶菌门(Olpidiomycota)(R=0.658)和壶菌门(Chytridiomycota)(R=0.58)呈显著正相关关系(P<0.05)。 土 壤 有 效 磷(AP) 与 隐 真 菌 门(Rozellomycota)(R=0.599)呈显著正相关关系(P<0.05),与蛙粪霉门(Basidiobolomycota)(R=-0.640)和芽枝霉门(Blastocladiomycota)(R=-0.592)呈显著负相关关系(P<0.05)。土壤速效钾(AK)与总丰度前14 的真菌门均无显著相关性。环境因子聚类分析表明,AN和SUR 聚为一类,AP 和AK 聚为一类。
采用Mantel 检验设施黄瓜连作不同年限土壤真菌群落结构和土壤理化性质之间的相关性(表2),可以看出,土壤真菌群落与pH、AN、SUR 和AP具有显著(P<0.05)或极显著(P<0.01)相关性,对真菌菌群影响表现为SUR>AP>AN>pH。
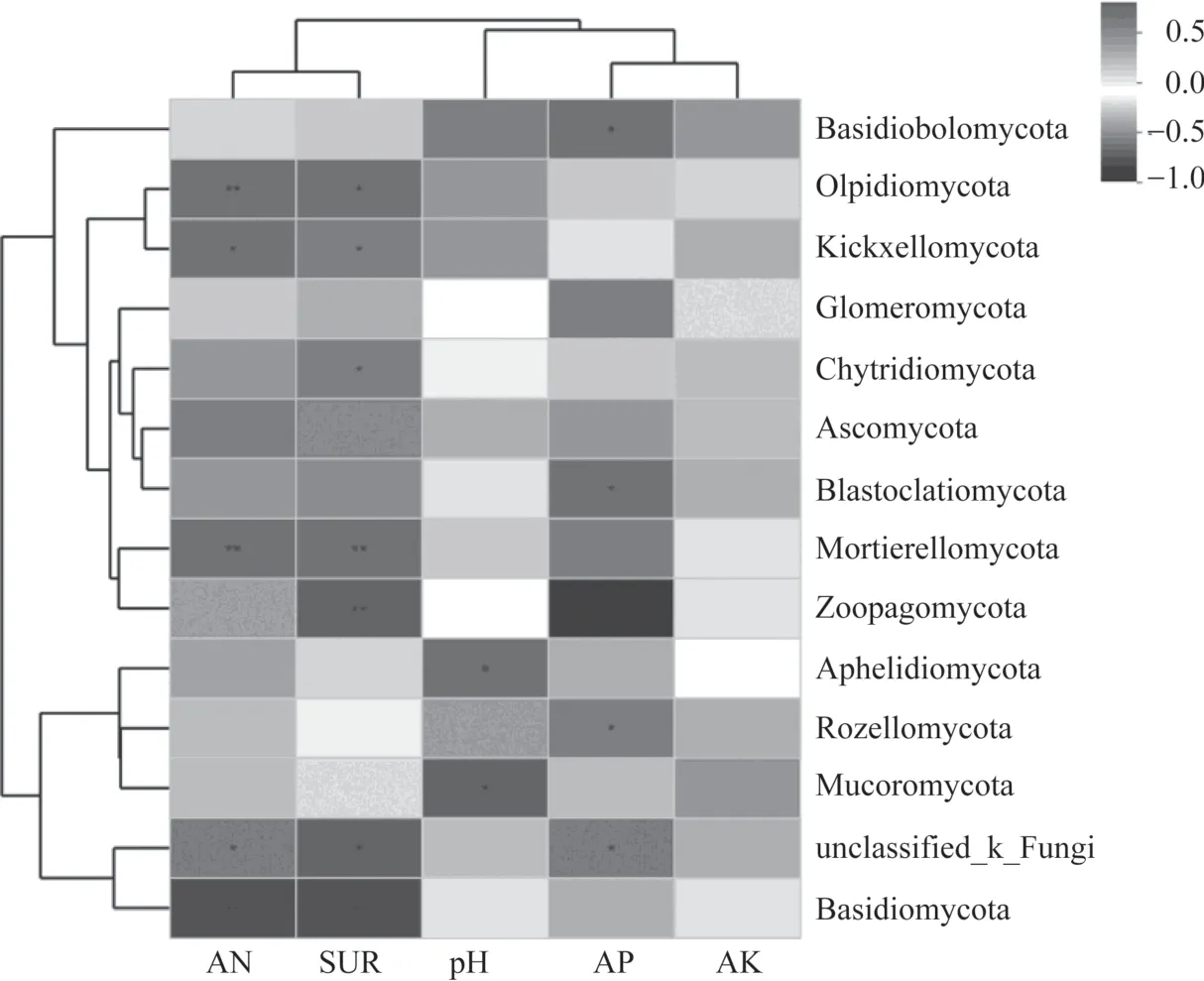
图7 总丰度前14 真菌门与土壤理化性质及酶活性相关性热图
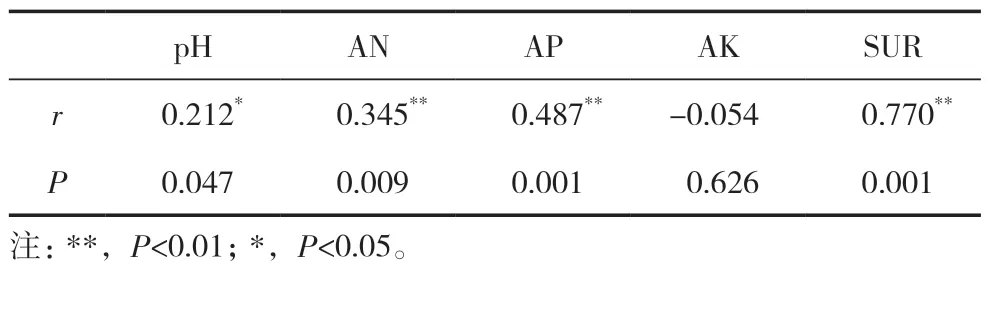
表2 温室黄瓜不同连作年限土壤真菌群落与土壤理化性质的Mantel 检验
3 结论与讨论
作物长期连作会导致土壤理化性质改变,土壤酶活性降低,土壤微生物总量、群落结构及多样性降低,从而导致其产量和品质降低[16-17]。真菌对土壤有机质的分解起着重要的作用,动植物残体的分解主要依靠腐生真菌来进行,土壤真菌的多少与土壤有机质含量密切相关[18]。长期集约化的温室连作栽培导致病害增多,尤其真菌型病害更为普遍[19-20]。随着新一代高通量测序技术的发展,关于土壤微生物群落和多样性的研究越来越多集中在连作与微生物群落的关系上。
本研究中,温室黄瓜连作不同年限后真菌群落组成在门和纲分类水平上发生变化。在门水平上,子囊菌门(Ascomycota)为优势菌门,相对丰度占55.74%~77.78%,其次是未被归类的真菌、担子菌门(Basidiomycota)和Mortierellomycota,壶菌门(Chytridiomycota)、Aphelidiomycota 和Rozellomycota 相对丰度占比很少,且在F1 中并未发现Rozellomycota 真菌门。日光温室黄瓜连作不同年限后真菌在门水平上的群落组成变化明显,子囊菌门相对丰度在F3 样品中无明显变化,在F5 和F7 样品中相对丰度分别下降9.94%和28.24%,而担子菌门相对丰度在F3 样品中显著降低后又逐年上升。随着连作年限的增加,微生物菌群的改变和丰度的变化会扰乱土壤微生态的平衡,更严重的会导致植物的死亡[21]。Schoch 等[22]研究表明,子囊菌门在根际土壤有机物的降解中起着重要作用,推测子囊菌门相对丰度的降低可能与土壤有机质和土壤肥力下降有关。
设施黄瓜长期连作增加了土壤中真菌的多样性,黄瓜连作后Shannon 指数随着连作年限的增加逐渐增高,但差异不显著,说明土壤样本的群落多样性逐渐升高;估计物种总数的Chao1 指数呈现先上升后下降的趋势且差异显著,表明连作5 和7 年土壤中真菌总数都相对较少,这一结果与不同连作年限怀牛膝的土壤微生物多样性的研究结果相一致[23]。冯翠娥等[24]同样在研究硒砂瓜连作对土壤真菌群落结构的影响时得到了一致的结果。这可能是由于随着连作年限的延长,长期高强度、重复的农业耕作破坏了土壤稳定结构,降低了土壤中微生物的丰度和均匀度,使得土壤由“细菌型”向“真菌型”转化,土壤中细菌群落多样性降低,真菌群落多样性增高。
研究表明,腐生菌参与有机物降解,植物病原菌/腐生/共生过渡型容易受到人活动的影响从而进行功能转换[25-27]。在本研究中,设施黄瓜连作不同年限根际土壤中真菌功能类群的相对丰度不同,腐生菌和植物病原菌/腐生/共生过渡型菌是黄瓜根际中两个主要功能类群。其中,腐生/共生过渡型菌(6.76%)和植物病原菌/腐生/共生过渡型菌(44.75%)两种过渡型在连作3 年土壤中相对丰度最高,植物病原菌/共生过渡型菌(18.22%)和植物病原菌/腐生/共生过渡型菌(0.91%)两种过渡型在连作5 年土壤中相对丰度最高,表明黄瓜根际土壤在连作3 和5 年逐渐过渡为共生菌(1.69%)和腐生菌(35.45%)型,其相对丰度在连作7 年样品中最高,植物病原菌(8.70%)的相对丰度在连作5 年达最高。本研究有50.24%的OTU 在数据库中无法进行功能分类,需要采用其他手段进一步研究。
在连作条件下,土壤理化性质可以有效地体现土壤的健康状况,影响土壤微生物种群数量及分布,土壤碱解氮、有效磷含量对土壤微生物区系从细菌型向真菌型转化和病害发生率都起着至关重要的作用[28],pH 是影响土壤真菌的主要环境因子[29],不同植被的营养代谢活动导致土壤中的养分含量具有一定的差异[30]。本研究中环境因子聚类分析表明,土壤AK 与总丰度前14 的真菌门均无显著相关性。环境因子聚类分析表明,AN 和SUR 聚为一类,AP 和AK 聚为一类,土壤真菌群落与pH、AN、SUR 和AP 具有显著(P<0.05)或极显著(P<0.01)相关性,对真菌菌群影响表现为SUR>AP>AN>pH。
