黑花生花针期干旱后复水的生理响应
2022-04-06赵玉坤宁东贤杨秀丽
赵玉坤,李 楠,宁东贤,杨秀丽
(山西农业大学小麦研究所,山西 临汾 041000)
【研究意义】黑花生富含硒、锌等微量元素和多种氨基酸[1-2],产量与普通红衣花生相近,但其营养价值、商品附加值更好,经济效益更高,种衣提取色素可广泛应用于食药行业[3],加工开发前景广阔[4],对我国种植业优化调整中作物种类布局有较好的借鉴意义。山西省地处黄土高原干旱、半干旱地区,降水量少且时空分布不均[5],在作物生长季易形成“旱后降水”的农田小气候。作物干旱复水后的快速生长和生理修复过程[6-8],可部分补偿干旱胁迫对植株生长造成的损害[9-10],也是其对外界环境变化的一种适应机制[11],其生理响应及恢复程度与作物抗旱性密切相关。【前人研究进展】花生作为一种耐旱性油料和经济作物[12-13],花针期是水分需求关键时期之一[14]。前人对其干旱胁迫后的生理伤害和适应机制研究较多[15-17],而在干旱复水后花生生理恢复机理研究主要集中在叶片光合特性、叶绿素荧光参数、农艺性状和产量表现等方面[18-20],通过生物产量和叶片生理特性来综合评价花生耐旱性及旱后复水补偿能力。【本研究切入点】根系是花生首先感应土壤干旱胁迫的重要器官,叶片保护酶系统可主动保护膜结构使花生免受逆境胁迫伤害,针对干旱复水后花生根系活力、叶片保护酶系统的生理补偿机制研究尚不多见。【拟解决的关键问题】本文以山西南部种植的2个黑花生品种(系)为试验材料,分析花针期干旱处理后间隔不同时间复水条件下黑花生根系活力、叶片保护酶活性及光合特性变化规律,为黑花生旱作栽培提供理论参考。
1 材料与方法
1.1 试验材料
试验于2018年6—10月在山西省农科院小麦研究所试验基地进行。供试2个品种(系)为豫花黑1号(YH)、临黑3号(LH),各选用一级果仁作种,盆栽种植,盆规格为内径60 cm,深55 cm,盆底部垫15 cm左右的泡沫层。取花生田0~20 cm耕层土,置于烘箱105 ℃烘干30 min,研碎、过孔径1 mm筛,与洗净、晾干后的沙子按5∶1混合,每盆装混沙土30 kg。取样地块前茬为休闲用地,土壤石灰性褐土,基本理化性质为:有机质14.10 g/kg,碱解氮37.20 mg/kg,速效磷10.38 mg/kg,速效钾110.00 mg/kg,pH 7.46。
1.2 试验设计
试验处理每盆18穴,单粒穴播,足墒播种。干旱处理设置4组:①CK,对照组,正常浇水,土壤相对含水量约70%;②T1,持续干旱处理3 d,干旱条件为土壤相对含水量约30%,过低则适度补水;③T2,持续干旱处理5 d;④T3,持续干旱处理7 d。在黑花生花针期干旱处理组结束浇水后,土壤相对含水量自然下降至约30%时开始实验,参考Hsiao[21]的方法,结合土壤水分速测仪和烘干法监测盆中土壤相对含水量,每天早晚各1次,实验期间严格控制土壤含水量,遇雨时用遮雨棚。
分4组测定复水后各项指标:①R0,复水前1 d,即干旱处理时测定相关指标;②R1,复水1 d时测定;③R2,复水4 d时测定;④R3,复水8 d时测定。每处理3次重复,干旱控水到设定天数时复水,持续补水到对照水平(土壤相对含水量70%)。
1.3 测定项目与方法
光合参数测定:分别在复水前1 d,复水后1、4、8 d时,每处理选取长势均匀一致的3株黑花生,用LI-6400光合仪测定黑花生功能叶(主茎倒3叶)的净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs),测定时间为上午9:00—10:00,按公式WUE=Pn/Tr计算叶片水分利用效率。沿叶柄基部剪下功能叶,锡箔纸包裹置于液氮罐中,用于叶片酶活性测定,最后将3株黑花生根系挖出,抖净土壤后带回室内冲洗干净,置于0 ℃冰箱内备用,用于根系活力测定。
根系活力测定:根系活力采用改良TTC法[22]测定。
保护酶系统相关指标测定:参考李合生[23]的方法测定样品叶片的超氧化物歧化酶(SOD)、过氧化物酶(POD)活性和丙二醛(MDA)含量。
1.4 数据分析
采用Excel 2010和SPSS 24.0软件对测定数据进行整理和统计分析。
2 结果与分析
2.1 干旱后复水对黑花生根系活力的影响
随着干旱胁迫时间增加,黑花生根系活力逐渐下降(图1),干旱7 d后,豫花黑1号和临黑3号的根系活力与对照(CK)相比,分别降低了65.44%和62.26%。2个黑花生品种干旱后根系活力随着复水天数的增多呈缓慢恢复趋势。方差分析(表1)表明,不同干旱处理间、不同复水天数间、干旱与复水两因素交互间的差异变化均达到极显著水平(P<0.01)。
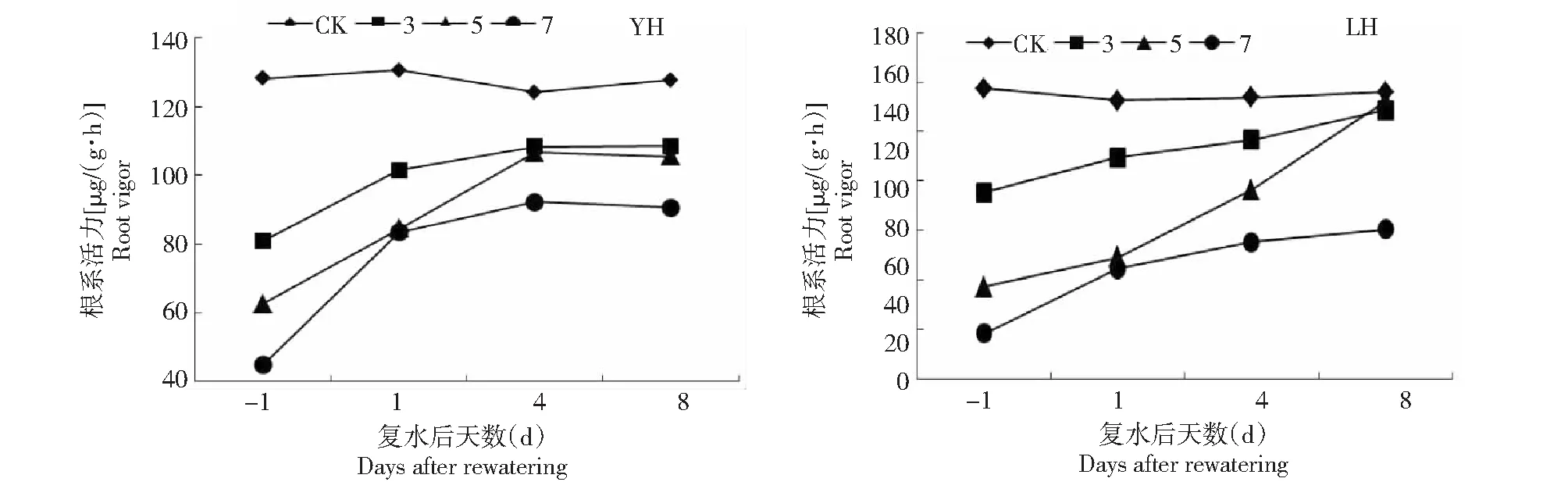
图1 花针期干旱复水对黑花生根系活力的影响Fig.1 Effects of black peanut root vigor under drought and rehydration in flower needle stage
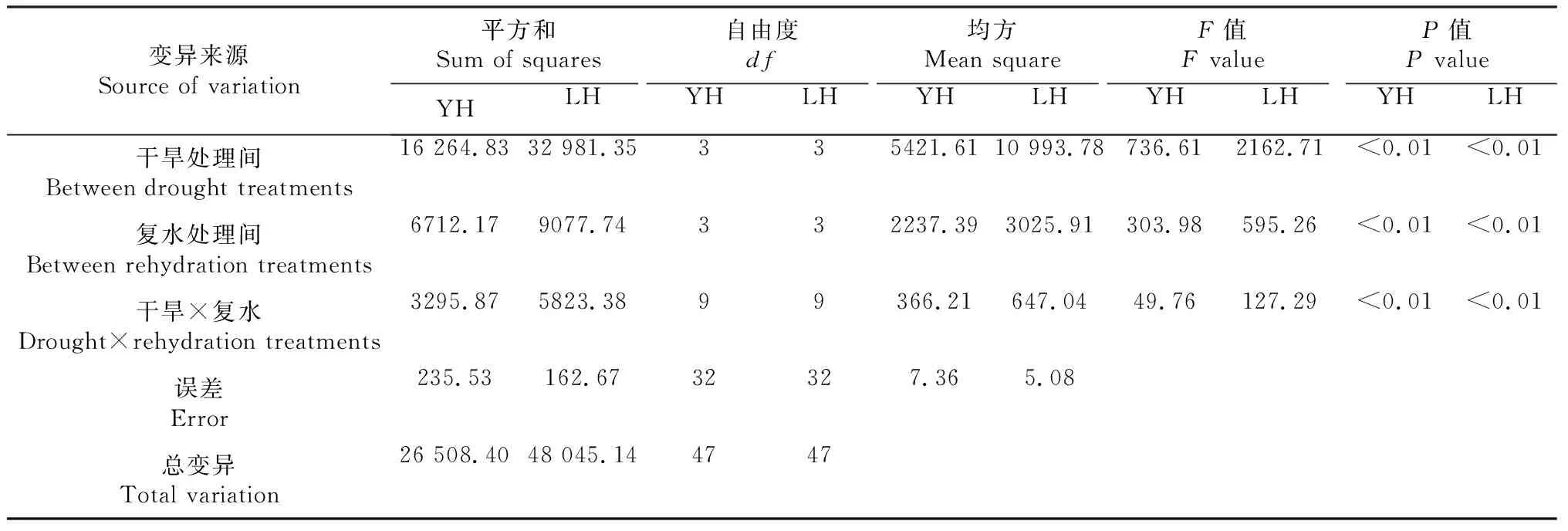
表1 不同黑花生品种干旱复水后根系活力方差分析
豫花黑1号干旱处理3、5 d后持续复水4 d,其根系活力可分别恢复至对照组(CK)的87.34%和85.92%,其后持续复水8 d,根系活力不再变化,干旱处理7 d后,根系活力遭受不可逆损伤,其后持续复水根系活力也仅可恢复至对照组(CK)的70.95%;其在适度干旱(如5 d)胁迫后复水根系活力恢复能力更强,但耐旱性较差,对干旱胁迫更敏感。
临黑3号干旱处理3、5 d后持续复水,根系活力可缓慢恢复,复水第8天,根系活力分别恢复至对照组(CK)的95.28%和97.72%,干旱7 d已对临黑3号根系造成损伤,后复水8 d仅可恢复至对照组(CK)的64.40%;其根系活力在适度干旱胁迫(如3、5 d)后复水可恢复至对照水平,耐旱性较好。
2.2 干旱后复水对黑花生叶片丙二醛含量及保护酶活性的影响
MDA是膜脂过氧化的分解终产物,其含量水平可代表植株遭受干旱逆境伤害程度。随着干旱胁迫时间的增加,豫花黑1号、临黑3号植株叶片中MDA含量逐渐增加(表2),干旱胁迫处理7 d后,与对照相比,2个黑花生品种(系)叶片MDA含量分别较对照增加288.56%和262.81%。干旱后持续复水可部分缓解胁迫对植株的伤害,在干旱5 d复水8 d处理后,2个黑花生品种(系)叶片MDA含量可恢复至对照(CK)水平,与对照相比无显著差异(P>0.05)。干旱胁迫处理7 d会对叶片造成不可逆损伤,其后持续复水8 d,叶片MDA含量仍维持在较高水平,2个黑花生品种分别达到对照的2.87和2.62倍,显著高于对照(P<0.05)。
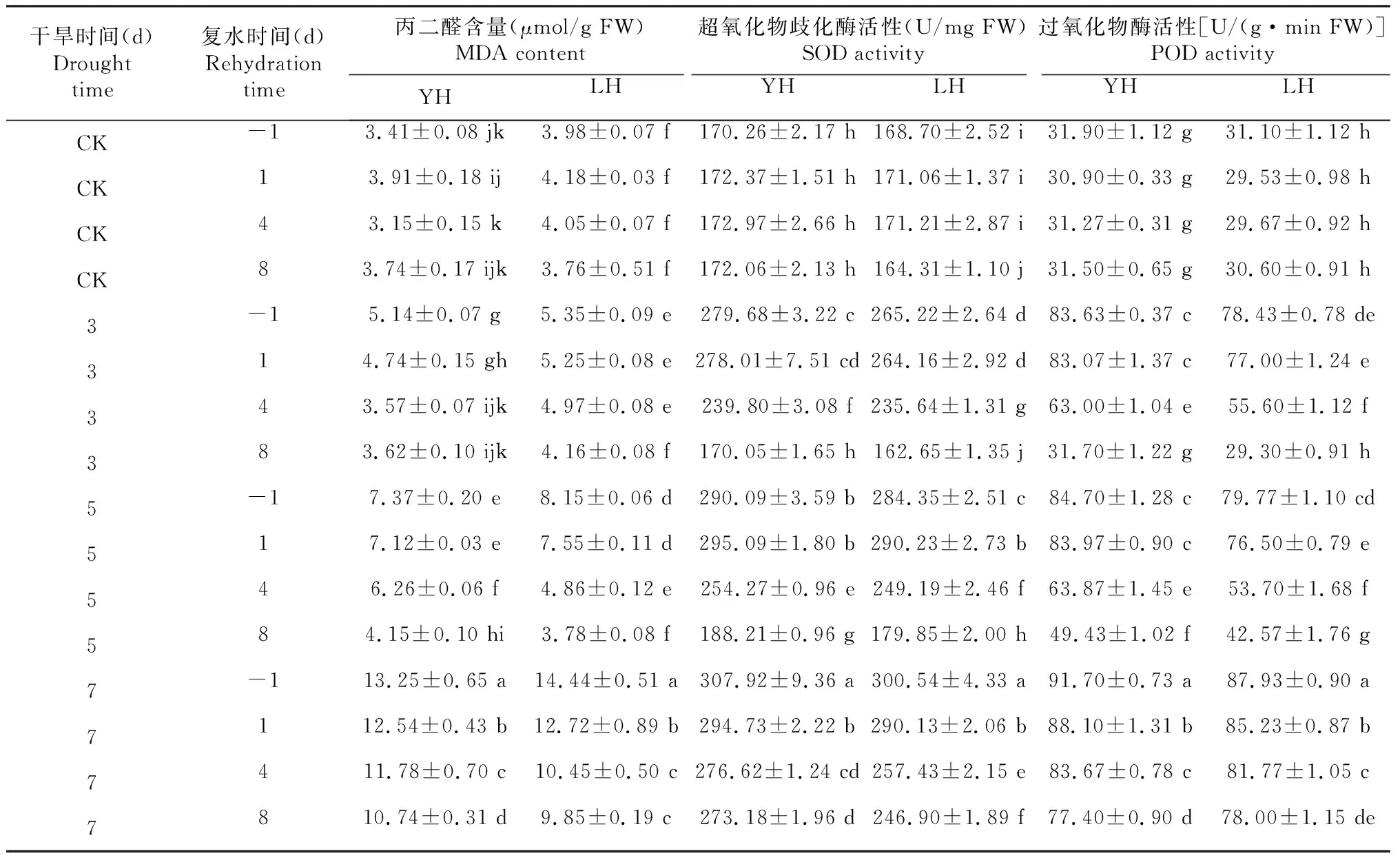
表2 不同黑花生品种干旱复水后叶片丙二醛含量及保护酶活性分析
超氧化物歧化酶(SOD)和过氧化物酶(POD)是植物主动应对膜脂过氧化伤害的2种重要保护酶。随着叶片膜脂过氧化产物(MDA)含量的增多,SOD、POD活性逐渐增强,2个黑花生品种峰值均出现在干旱7 d胁迫处理,分别较对照增加1.81、1.78倍和2.87、2.83倍,其后变化趋势与MDA含量变化趋势相似。在2个黑花生品种中,叶片MDA含量较低时,SOD、POD活性也较低,当植株受到干旱胁迫损伤较大时,叶片MDA含量升高,SOD、POD保护酶活性增强。方差分析(表3)显示,叶片MDA含量及SOD、POD活性在品种、干旱、复水处理间和3处理因素交互间差异极显著(P<0.01)。
2.3 干旱后复水对黑花生叶片光合参数的影响
随着干旱胁迫时间的增加,2个黑花生品种(系)的叶片净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)均呈下降趋势(表3)。干旱7 d后,2个黑花生品种(系)4个光合参数值分别较对照(CK)下降13.16%、33.36%、20.35%、56.58%和14.01%、23.61%、19.50%、27.88%。轻度干旱胁迫处理(3 d)提高了2个黑花品种的叶片Pn值,分别较对照增加了14.48%和1.64%,而Tr、Ci和Gs3个光合参数值均低于对照值(P<0.05)。中、重度干旱处理(5和7 d)后的2个黑花生品种叶片4个光合参数值均显著低于对照(P<0.05)。干旱胁迫5 d后持续复水8 d,豫花黑1号叶片4个光合参数值可分别恢复至对照的103.91%、98.78%、99.07%和92.92%,而临黑3号可恢复至对照的101.71%、101.05%、100.14%和100.00%,说明临黑3号的干旱后补偿修复能力强于豫花黑1号。2个黑花生品种在干旱胁迫处理7 d后持续复水8 d,Pn、Tr、Ci和Gs4个光合参数值均显著低于对照(P<0.05),表明重度干旱对叶片造成不可逆损害,即使持续复水也仅可部分恢复,达不到正常植株叶片的功能水平。
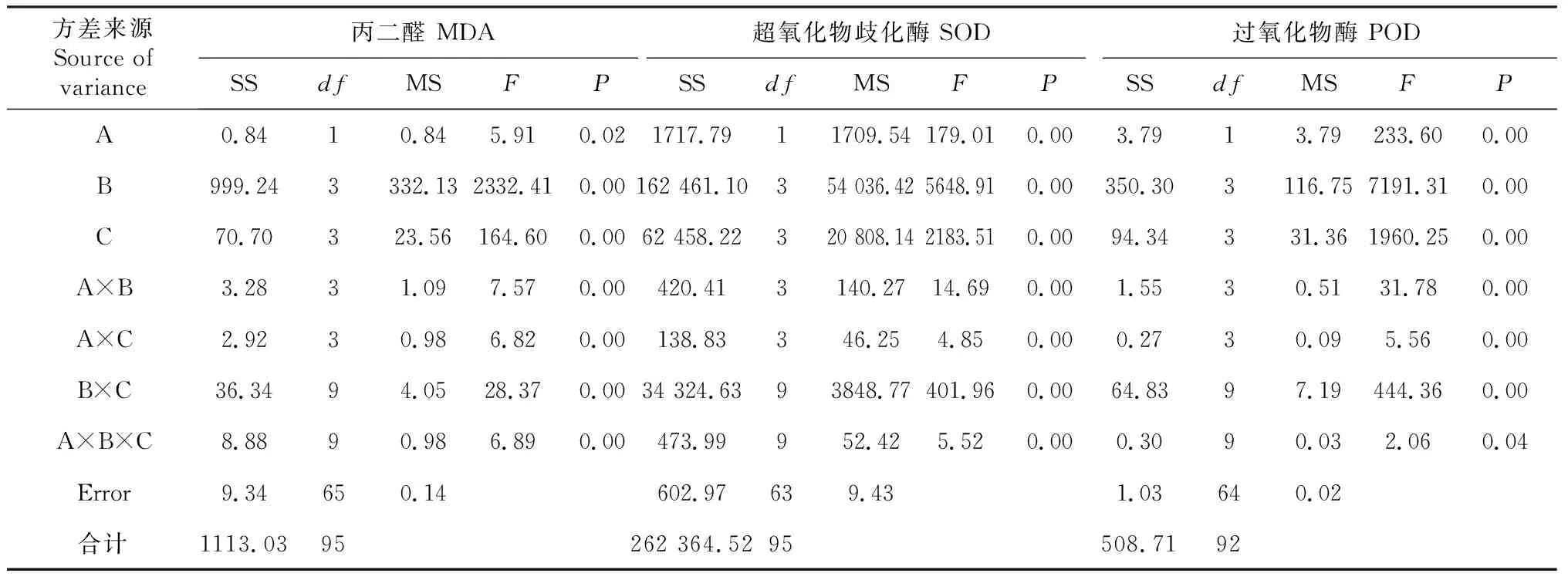
表3 不同黑花生品种干旱复水后保护酶活性及丙二醛含量的方差分析
净片净光合速率(Pn)和蒸腾速率(Tr)的比值(WUE)反映植物叶片中水分参与光合同化反应的比值,其值越高,叶片水分利用效率越高,光合源产物也相应增加。随着干旱胁迫时间的增加,2个黑花生品种水分利用效率(WUE)均有不同幅度的增加,峰值出现在干旱3 d后,2个黑花生品种WUE值分别较对照增加了36.59%和22.62%。适度干旱处理后,随着复水进程持续,2个黑花生品种WUE值逐渐降低,但均显著高于对照(P<0.05),说明适度干旱后复水可增强植株叶片耐旱性,提高水分利用效率。方差分析(表5)结果表明,叶片光合参数Pn、Tr、Ci、Gs和WUE在品种、干旱、复水三因素处理间及各因素交互间差异极显著(P<0.01)。
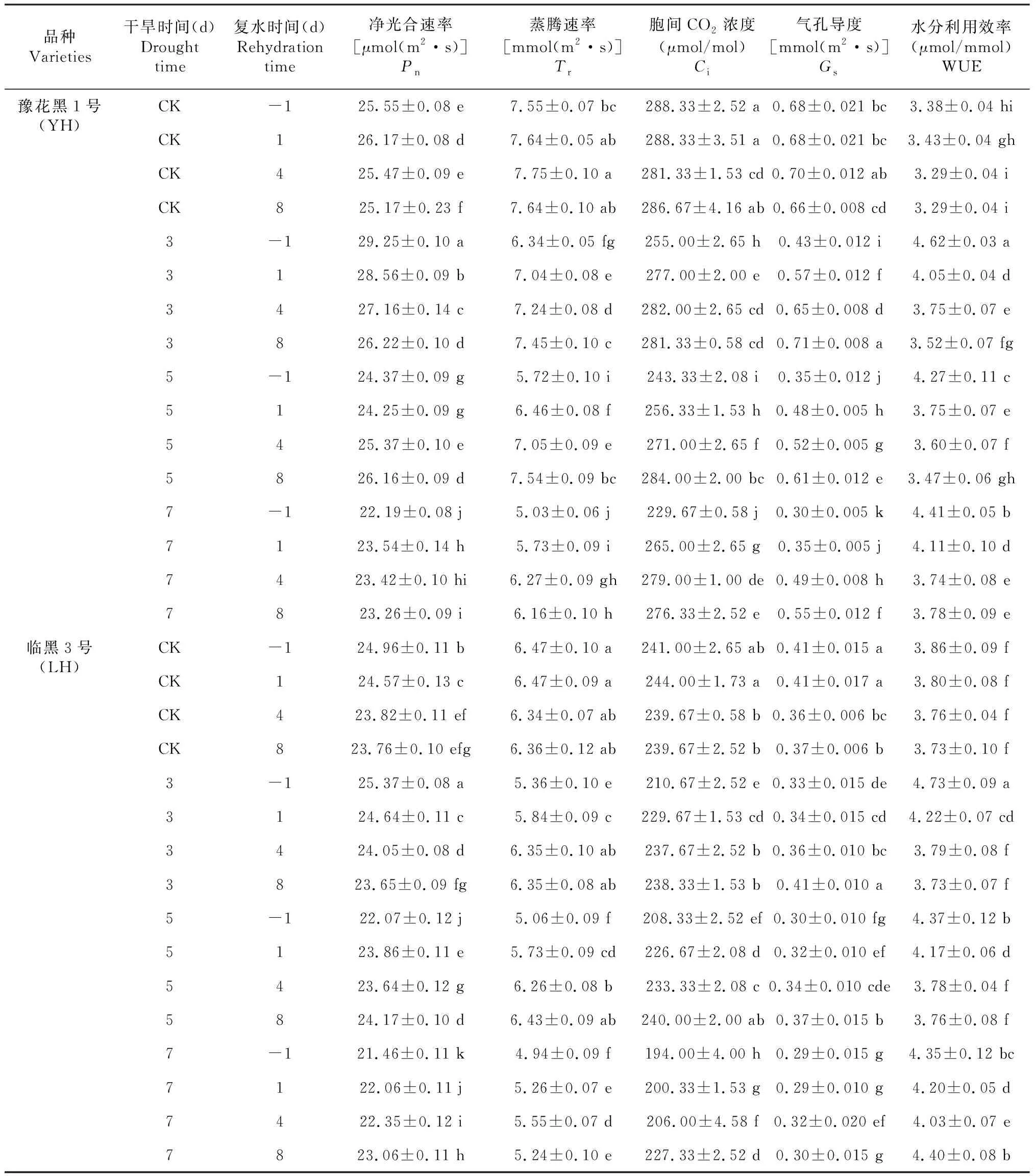
表4 不同黑花生品种干旱复水后叶片光合参数分析
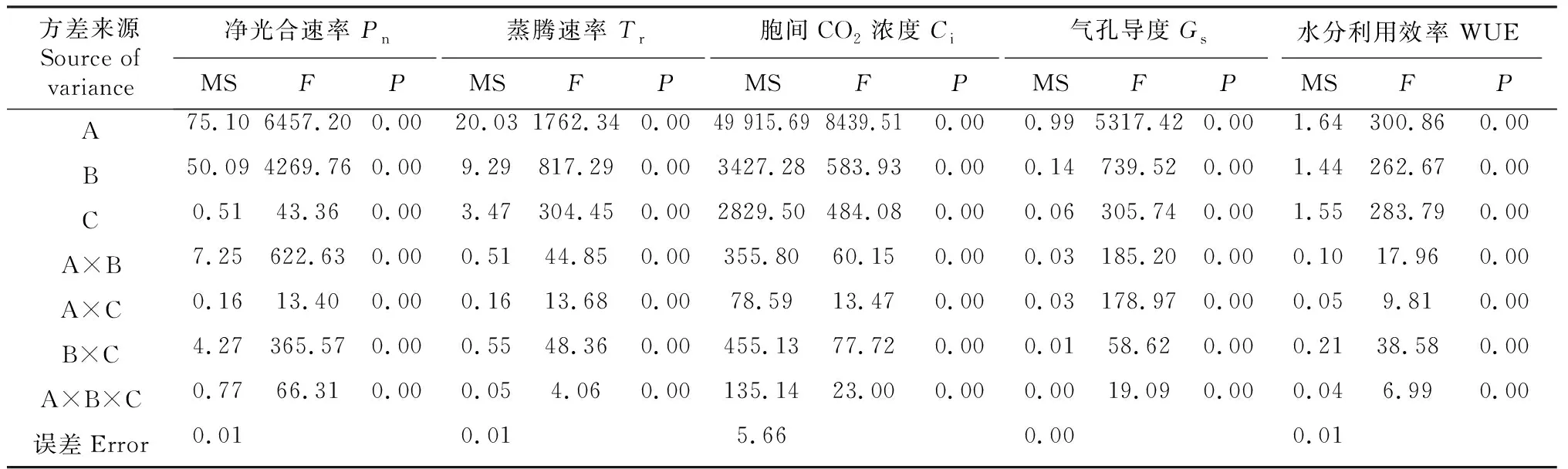
表5 不同黑花生品种干旱复水后光合参数值方差分析
3 讨 论
3.1 干旱复水后根系活力变化
根系是作物首先感应土壤干旱胁迫的重要器官。干旱环境胁迫影响作物根系形态发育及生理性状,进而影响地上部分(茎、叶)的生长。干旱条件下根系活力与叶片总叶绿素含量显著正相关[24]。适度干旱后,作物根系大量合成脱落酸这一根源逆境信号物质,调节植株生长发育,以度过干旱逆境[25-26]。作物干旱后复水,根系活力可快速恢复,其恢复能力与作物类型和品种有关,但重度干旱胁迫会对根系造成不可逆的损伤,进而影响植株生长发育[27]。在本研究中,豫花黑1号和临黑3号在适度干旱胁迫(5 d)后持续复水8 d,根系活力可分别恢复至对照的85%和95%以上,临黑3号的旱后复水补偿生长能力优于豫花黑1号。重度干旱胁迫(7 d)处理后,2个黑花生品种(系)的根系均有不同程度地损伤,持续复水8 d也不能恢复至对照水平,植株生长发育受到抑制。
3.2 干旱复水后叶片丙二醛含量和保护酶活性变化
丙二醛(MDA)作为植物遭受逆境胁迫时膜质过氧化的最终产物,其含量高低与植物细胞膜受损程度密切相关[28-29]。干旱胁迫时,植物叶片中活性氧代谢平衡被打破,自由基增多,植物会主动调节保护酶系统(SOD、POD等)活性维持在较高水平,清除自由基,保护膜结构,减小膜质过氧化对植物细胞的伤害[30-32]。叶片保护酶活性在干旱胁迫几天后迅速增加,达到峰值,然后随着干旱程度地加剧,酶结构遭到不可逆破坏,其活性又会迅速下降[33]。适度干旱胁迫后持续复水,叶片保活酶活性呈“凸”字形变化趋势,可有效保护作物免受干旱损害,锻炼品种抗旱性[34-35]。在本研究中,随着干旱胁迫天数的增加,豫花黑1号、临黑3号叶片丙二醛含量逐渐增加,与之对应的是叶片中超氧化物歧化酶、过氧化物酶活性升高,保护叶片膜结构免受自由基损害。中度干旱(3~5 d)经持续复水处理后,2个黑花生品种(系)叶片丙二醛含量会逐渐下降至对照水平,SOD、POD活性也随之降低,重度干旱(7 d)后2个黑花生品种(系)叶片保护酶系统均遭受不可逆损伤,不能恢复至对照水平,这种趋势也反映出2个黑花生品种(系)叶片保护酶系统在旱后复水条件下的补偿恢复能力:豫花黑1号和临黑3号叶片保护酶恢复能力相近,但临黑3号对重度干旱胁迫的调节恢复能力要强于豫花黑1号。
3.3 干旱复水后叶片光合特性变化
干旱条件下,作物叶片光合效率(Pn)的降低主要原因是气孔部分关闭,同时蒸腾速率(Tr)和胞间CO2浓度(Ci)也随之下降[36-37]。光合效率是作物产量形成的基础,干旱胁迫影响叶片光合产物的分配格局[38-39],降低光合效率,复水后叶片中的光合产物向外运输,在植株水平上表现出生长补偿[40-41]。在本研究中,随着干旱胁迫天数的增加,叶片Pn、Tr、Ci、Gs4个光合参数值均呈下降趋势,复水后豫花黑1号Pn恢复迅速,而临黑3号恢复持续时间长,且复水后最终Pn值略高于对照(CK)。适度干旱处理(3~5 d)后复水,2个黑花生品种(系)叶片水分利用效率(WUE)值均显著高于对照(P<0.05),说明适度干旱锻炼可提高作物植株抗旱性。与根系活力、保护酶活性变化趋势一致,重度干旱胁迫(7 d)亦可对2个黑花生品种(系)叶片光合特性造成不可逆损害,最终影响光合产物的分配格局。
4 结 论
适度干旱胁迫可在一定程度上锻炼黑花生品种(系)的抗旱性,其通过根系活力、叶片保护酶活性、光合特性等生理过程综合调节植株整体抗旱性,复水后这些指标可恢复至正常水平,部分生理指标在适度干旱胁迫复水后甚至要高于对照,抗旱性更强;但重度干旱胁迫会对黑花生植株造成不可逆生理损害。本试验中豫花黑1号复水响应迅速,但对干旱胁迫更敏感;而临黑3号复水持续响应时间较长,耐旱性相对较好。
干旱复水条件下黑花生根系活力、叶片丙二醛含量、保护酶活性及光合参数的变化规律,可为黑花生抗旱性研究和复水恢复能力强的品种(系)筛选提供理论依据,下一步应对不同品种旱后复水条件下“根茎叶”各器官生理协同调节的物质分配格局及分子调控机理进行更深入地研究。
