马铃薯组织中SGAs含量差异及其基因表达分析
2022-04-06李红艳李洁雅叶广继
李红艳,李洁雅,叶广继,2,3,4,5,苏 旺,2,3,4,5,周 云,2,3,4,5,王 舰,2,3,4,5
(1.青海大学,西宁 810016;2.青海省农林科学院,西宁 810016;3.青海大学青藏高原生物技术教育部重点实验室,西宁 810016;4.青海省马铃薯育种重点实验室,西宁 810016;5.省部共建三江源生态与高原农牧业国家重点实验室,西宁 810016)
【研究意义】糖苷生物碱(Steroidal glycoalkaloids,SGAs)是一类重要的植物次生代谢产物,主要存在于茄科和百合科植物中[1-2]。马铃薯栽培种含有50多种SGAs,95%以上是α-茄碱和α-卡茄碱[3-5]。马铃薯块茎中,SGAs主要分布在1.5 mm厚的皮层下[6]。全球种植5000多个马铃薯品种,因品种及贮藏方式不同,各个马铃薯之间SGAs含量差异较大[7]。马铃薯中SGAs含量在20 mg/100 g(鲜质量)、1 mg/g(干质量)范围内可以安全食用,达到20 mg/100 g会引起中毒,高于30~60 mg/100 g则会致死[8-11]。SGAs不仅对植物具有一定的防御作用,而且它能抑制人体内胆碱酯酶的活性[12-15]。研究发现,SGAs具有抗癌、抗炎、抗菌等药理活性,其中α-卡茄碱的药理活性是α-茄碱的2倍[16-19]。但马铃薯中SGAs含量过多,其毒性会破坏人体细胞膜结构,危害正常的中枢神经系统,影响马铃薯的安全食用和生产加工。【前人研究进展】关于马铃薯中SGAs的研究已有许多报道,Li等[20]研究发现,樟脑可抑制马铃薯块茎的发芽,从而降低SGAs的积累,孟卫芹等[21]研究发现,通风、低氧等条件下可延迟马铃薯发芽,使SGAs积累减少,霍权恭等[22]研究发现,马铃薯贮藏时间越长,SGAs积累越多。SGAs来源于胆固醇,胆固醇经过多次羟基化、氧化和转胺化反应,最终生成α-茄碱和α-卡茄碱[23]。马铃薯茎和块茎等生长旺盛的组织中容易合成SGAs,其生物合成分为萜类、甾醇类与茄啶三个阶段,合成过程中所需要的关键结构基因(如StSGT和StSSR2等)已有研究报道[24-26]。马铃薯作为主粮作物,其块茎不仅供人类食用,也可用于薯片、薯条等加工。目前,国内在选育马铃薯品种时对产量、抗病性、淀粉含量等性状关注较多,但是块茎中SGAs含量缺乏系统的检测及评价,可能是因为SGAs的检测需要专业设备和人力物力。【本研究切入点】随着主粮化的进程,我国马铃薯人均消费量将增加,马铃薯的食用安全需得到保障。储藏条件会使马铃薯块茎的品质发生较大改变,例如低温糖化、变绿发芽都会影响马铃薯产业的健康发展。【拟解决的关键问题】选育块茎中低SGAs含量的马铃薯是今后育种的主要方向之一,而且SGAs具有药理活性,马铃薯芽中SGAs含量高,部分马铃薯资源块茎长期储藏后SGAs含量增加,无法食用,均可作为提取SGAs的原料,为马铃薯的综合利用提供新思路以及为马铃薯的安全食用提供参考。
1 材料与方法
1.1 材料
马铃薯资源1-76、6-36、5-36、6-1来自青海省农林科学院生物技术研究所。
1.2 方法
1.2.1 标准溶液的配制 将α-茄碱和α-卡茄碱标准品用40%的甲醇水溶液(含0.1%甲酸)配制成400 ng/mL的标准中间液,再分别将标准中间液用40%的甲醇逐步稀释成4 μg/mL的混合标准工作液,取适量混合标准工作液用40%甲醇逐级稀释成浓度为25、50、100、200、300、400、500、600 ng/mL的混合液,过0.22 μm滤膜后,4 ℃冰箱保存备用。
1.2.2 样品处理 取地窖贮藏60 d的紫皮马铃薯资源(1-76、6-36)和白皮马铃薯资源(5-36、6-1)的块茎,光照60 d后的发芽马铃薯芽冷冻干燥后研磨。分别称取块茎和芽样品粉末0.2 g,加入5%的乙酸5 mL,涡旋混匀后超声处理30 min,8000 r/min离心5 min后收集上清液,上清液用40%的甲醇稀释50或500倍,过0.22 μm滤膜,待测。
使用美国Agilent公司生产的Agilent 1260 InifityⅡ-Agilent 6470高效液相色谱—串联质谱仪检测α-茄碱和α-卡茄碱含量。α-茄碱和α-卡茄碱标准品购自美国Sigma公司。
1.3 SGAs生物合成相关基因表达分析
1.3.1 引物设计及合成 根据SGAs合成通路,选取StHMG1,StSQS1,StCAS,StSSR2,StGAME4,StGAME8a,StSGT1,StSGT2,StSMT1和StCYP51G等结构基因,送上海生物工程公司合成。
1.3.2 总RNA的提取及cDNA的合成 采用TaKaRa提取试剂盒提取马铃薯资源1-76、6-36、5-36、6-1块茎和芽中的总RNA(图1)。
M. DL2000;1~4为马铃薯块茎(1-76、6-36、5-36、6-1)的RNA;5~8为马铃薯芽(1-76、6-36、5-36、6-1)的RNAM. DL2000; 1-4 is the RNA of potato tubers (1-76, 6-36, 5-36, 6-1);5-8 is the RNA of potato buds (1-76, 6-36, 5-36, 6-1)图1 马铃薯块茎、芽RNA电泳图Fig.1 RNA electrophoresis of potato tubers and buds
cDNA的合成参照5×Prime Script TMRT Master Mix(Perfect RealTime)(TaKaRa)说明书进行,整个过程在冰上。反应体系20 μL:5×Prime Script RT Master Mix(Perfect Real Time)4 μL,总 RNA体积等于1000除以RNA浓度,用RNase Free ddH2O补充至20 μL。反应条件:37 ℃ 15 min,85 ℃ 3 s,4 ℃无限循环。将得到的cDNA溶液稀释至200 ng/μL,-20 ℃保存备用。
1.3.3 qRT-PCR检测 以cDNA为模板,Actin为内参基因。反应体系20 μL:cDNA模板2 μL,引物-F(10 μmol/L)0.4 μL,引物-R(10 μmol/L)0.4 μL,TB Green premix Ex Taq II 10 μL,ddH2O 7.2 μL。Real Time PCR反应程序为:95 ℃ 30 s;95 ℃ 10 s,61 ℃ 32 s, 40个循环;95 ℃ 60 s,60 ℃ 60 s,55 ℃ 10 s。
2 结果与分析
2.1 4种马铃薯资源在不同储藏条件下的表型变化
对4种马铃薯资源分别进行地窖和光照储藏60 d(图2)后,地窖贮藏的马铃薯开始发芽,而光照下的马铃薯芽长至1.5 cm左右,皮色发绿。推测由于储藏温度对马铃薯的表型有一定影响。

a为地窖贮藏60 d,b为光照贮藏60 da is cellar storage for 60 days, b is light storage for 60 days图2 4种马铃薯资源储藏60 d后的变化Fig.2 Changes of four kinds of potato resources after storage for 60 days
2.2 糖苷生物碱含量分析
高效液相色谱—串联质谱法(HPLC-MS/MS)测定显示,马铃薯块茎中SGAs含量变化范围为0.034~3.360 mg/g,芽中SGAs含量变化范围为24.518~41.579 mg/g。2种紫皮马铃薯资源(1-76、6-36)块茎和芽中的SGAs含量变化无显著差异,2种白皮马铃薯资源(5-36、6-1)中SGAs含量在块茎中差异性显著,但在芽中差异不显著(表1)。因此,选用2种白皮马铃薯资源为试验材料进行相关基因表达性分析。
2.3 SGAs生物合成相关基因在马铃薯资源中的表达量分析
使用qRT-PCR方法检测了2种白皮马铃薯资源(5-36、6-1)SGAs合成途径中StHMG1,StSQS1,StCAS,StSSR2,StGAME4,StGAME8a,StSGT1,StSGT2,StSMT1和StCYP51G10个相关基因的相对表达量。块茎中,SGAs含量5-36显著高于6-1(表1),StCAS,StSSR2,StGAME4,StSGT1,StSGT2和StSMT1这6个基因的表达量5-36显著高于6-1,其他基因在5-36中的表达量低于或接近6-1(图3-a);芽中,SGAs含量5-36略高于6-1,表达量检测发现,除了StSMT1基因外,其他9个基因的表达量在5-36显著高于6-1(图3-b)。通过基因表达量结果发现,SGAs生物合成途径中部分基因(StCAS,StSSR2,StGAME4,StSGT1,StSGT2)的表达量与SGAs含量之间的关系密切。
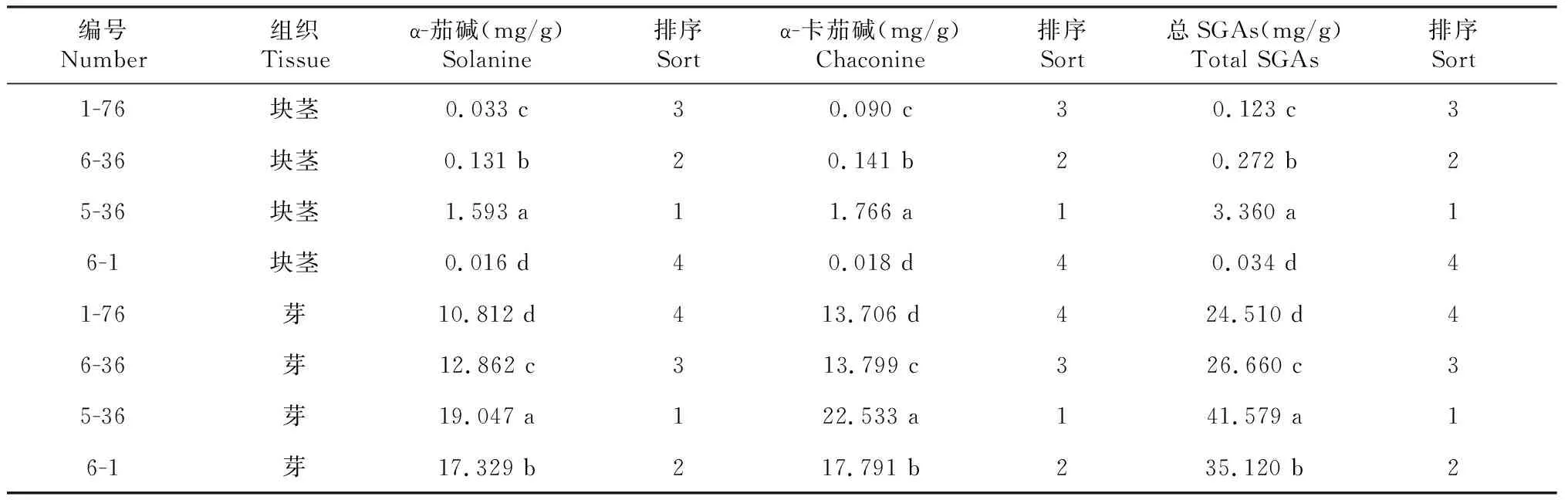
表1 不同马铃薯资源块茎、芽中的SGAs含量

图3 SGAs生物合成相关基因在马铃薯资源(5-36、6-1)块茎(a)和芽(b)中的表达量变化Fig.3 Changes in expression of SGAs biosynthesis-related genes in potato resources (5-36, 6-1) tubers (a) and buds (b)
由表1可知,所有马铃薯芽中的SGAs含量显著高于块茎。在马铃薯资源5-36块茎和芽不同组织中,参与SGAs合成的相关基因表达量存在差异,其中StHMG1,StSSR2和StGAME8a基因的表达量在芽组织中高于块茎,其他基因的表达则相反或相近。然而在马铃薯资源6-1块茎和芽不同组织中,基因表达量分析结果发现,仅有StSSR2和StGAME8a基因的表达量在芽中显著高于块茎,其他基因在芽中的表达量低于块茎或二者相当(图4)。表明不同马铃薯资源在调控不同组织SGAs的合成途径中存在共同机制,但有细微差别。此外,参与SGAs合成的部分基因可能参与了其他生物合成过程。

图4 SGAs生物合成相关基因在5-36(a)和6-1(b)块茎和芽中的表达量变化Fig.4 Changes in expression of SGAs biosynthesis-related genes in 5-36(a) and 6-1(b) tubers and buds
3 讨 论
马铃薯中SGAs具有多种生物学功能,尤其马铃薯块茎中其含量过高将影响食用安全,因此SGAs合成调控机理一直以来是研究的热点。本试验中,不同马铃薯资源块茎和芽中的SGAs含量差异较大,这与前人研究结果相符[27-29]。测定的不同马铃薯资源块茎中α-茄碱含量为0.016~1.593 mg/g,α-卡茄碱含量为0.018~1.766 mg/g,马铃薯资源芽中α-茄碱含量为10.812~19.047 mg/g,α-卡茄碱含量为13.706~22.533 mg/g,α-卡茄碱含量比α-茄碱含量高,符合前人研究结果[30-31]。4个马铃薯资源块茎中SGAs含量为0.034~3.360 mg/g,芽中SGAs含量为24.510~41.579 mg/g,在储藏60 d后有2个马铃薯资源1-76(0.123 mg/g)和6-1(0.034 mg/g)块茎中的SGAs含量未超出安全食用范围(1 mg/g)(干质量),其余马铃薯资源块茎和芽中的SGAs含量均已超出安全食用标准。
SGAs的合成是通过甲羟戊酸途径实现的,其中羟基-3-甲基戊二酰辅酶A还原酶(HMG1)基因是SGAs甲羟戊酸合成途径中的第一步催化酶,马铃薯块茎中SGAs含量与HMGR和鲨烯合成酶(PSS1/SQS)基因的表达量呈正相关[32],但本试验结果中StHMG1基因的表达量与SGAs含量变化规律不一致,实验材料的不同是造成与前人研究存在差异的原因之一,StHMG1基因在马铃薯中以基因家族的形式存在,但是该家族中不同基因的功能存在差异。环阿屯醇合成酶(CAS1)位于2,3-环氧化鲨烯合成环阿屯醇的分支点,StCAS基因表达量在不同资源中与SGAs含量变化规律一致,但是StCAS基因表达量在同一资源不同组织中与SGAs变化规律相反,SGAs含量低的块茎中,该基因的表达量高,推测其不仅参与SGAs的生物合成过程,还参与了其他生物合成过程。马铃薯甾醇侧链还原酶2(StSSR2)是以环丙烯醇为原料合成SGAs的前体胆固醇的关键酶,位于茄碱和油菜素内酯合成的分支点,其不仅催化甾醇类物质合成,而且同家族的StDWF1基因催化油菜素内酯合成[33-34]。本试验中,StSSR2基因主要在马铃薯芽中表达,块茎中几乎不表达,并且,在SGAs含量高的马铃薯资源(5-36)不同组织中,该基因的表达量均高于SGAs含量低的马铃薯资源(6-1),表明该基因可能主要负责马铃薯SGAs的合成。此外,敲除或沉默StSSR2基因均可以降低马铃薯中SGAs含量[35-37]。
研究发现,马铃薯中StSSR2基因主要在块茎和茎中表达,叶片次之,根和匍匐茎中表达量最少[38],但在芽中还未见报道,StSSR2基因的功能有待进一步深入研究。细胞色素P450单加氧酶基因(Glycoalkaloid Metabolism 4,GAME4)催化SGAs生物合成途径下游的生化反应,沉默该基因使马铃薯块茎中SGAs含量显著降低[39-40]。本研究中,StGAME4基因的表达模式与StCAS基因一致,同一资源不同组织中该基因的表达量与SGAs含量变化规律相反。SGAs生物合成过程中的糖基化过程由半乳糖基转移酶(SGT1)和葡糖基转移酶(SGT2)催化合成。安然等[41]研究发现SGT1、SGT2和SGT3基因被敲除后,马铃薯块茎中SGAs含量降低。McCue等[42]抑制StSGT1基因的表达,结果发现块茎中的α-茄碱几乎被完全抑制,但α-卡茄碱的积累水平却提高了,导致总体SGAs的含量没有太大改变。本试验结果表明,StSGT1,StSGT2基因表达模式与StCAS基因一致,推测这2个基因负责不同马铃薯资源中SGAs含量,而对同一资源不同组织中SGAs含量贡献较小。甾醇C-24-甲基转移酶1(SMT1)作为植物甾醇类生物合成分支的关键酶,将环丙烯醇转化为24-亚甲基环丙烯醇,前人研究发现在马铃薯中过表达大豆中GmSMT1基因,叶片和块茎中总SGAs及游离胆固醇含量显著下降,与SGAs含量改变一致[43]。然而,本实验中StSMT1基因的表达量与SGAs含量变化规律存在差异。钝叶醇14-α-去甲基化酶(StCYP51G)主要参与马铃薯中植物甾醇(菜油甾醇和谷甾醇)的生物合成,StCYP51G基因在不同马铃薯资源块茎和芽中的表达量与SGAs含量变化规律相关性不高。
4 结 论
4种马铃薯芽中的SGAs含量显著高于块茎,块茎中SGAs含量变化为0.034~3.360 mg/g,芽中SGAs含量变化为24.510~41.579 mg/g,4种马铃薯芽及马铃薯6-36(0.272 mg/g)和5-36(3.360 mg/g)块茎中的SGAs含量均已超出安全食用值(1 mg/g,干质量),对人类安全食用存在风险。不同马铃薯资源中,SGAs的调控机制与不同组织中SGAs合成机制可能存在差异,为深入解析不同马铃薯资源中SGAs的合成机理以及马铃薯的综合开发利用提供一定的理论依据。
