利用分子标记辅助选择技术培育抗螟虫转基因杂交稻恢复系
2022-04-06刘开强李孝琼王小姣李林娟郭嗣斌
刘开强,李孝琼,韦 宇,陈 颖,王小姣,李林娟,郭嗣斌
(广西农业科学院水稻研究所/广西水稻遗传育种重点实验室,南宁 530007)
【研究意义】水稻是我国的主要粮食作物之一,其产量和品质对保障我国粮食安全至关重要[1-2]。螟虫(二化螟、三化螟和稻纵卷叶螟等)是水稻的主要害虫,对世界水稻生产造成的损失巨大,其中,二化螟在我国稻区分布较广,年防治面积达1500万hm2,年防治费用约6.5亿美元,年产值损失8.4亿美元[3-4]。使用化学杀虫剂防治螟虫效果虽然较好,但耗费巨大,而且还会造成环境污染,其农药残留甚至威胁人类健康。培育抗螟虫水稻新品种是最经济有效的水稻螟虫防治措施,可在提高水稻产量的同时保护生态环境安全[5]。但我国的水稻品种资源多为低抗或中抗螟虫材料,抗性低且不稳定,高抗螟虫品种资源较少,采用常规杂交育种方法很难培育出抗螟虫水稻品种,而利用转基因技术改良水稻抗虫性已成为抗虫育种的新方法[6]。在前人研究基础上,以转Bt基因水稻材料为供体与农艺性状优良的水稻品系杂交、回交及多代自交,通过分子标记辅助选择(Molecular marker-assisted selection,MAS)技术结合田间农艺性状筛选,是选育兼具优良农艺性状和抗虫新品种的有效途径[7-10]。因此,通过MAS技术进行抗螟虫水稻新恢复系选育,对我国杂交稻抗性育种具有重要意义。【前人研究进展】MAS可有效缩短育种周期和提高育种效率,在水稻、玉米和小麦等粮食作物及多种经济作物上已广泛应用[11-12]。苏云金芽孢杆菌(Baclillusthuringiensis)蛋白(Bt蛋白)对鳞翅目昆虫具有专一毒性,对人类无毒害作用。Fujimoto等[13]利用转基因技术将cryIA(b)基因转入粳稻,并使crylA(b)蛋白成功表达,从而培育出抗螟虫粳稻品种。Cheng等[14]通过农杆菌介导的转化获得2600个转cryIA(b)和cryIA(c)基因水稻植株,并已证实这些基因能在R0和R1代中整合、表达和遗传。Tu等[15]在Bt明恢63植株中检测到Bt融合蛋白cryIA(b)/cryIA(c)含量为20.0 ng/mg;对转基因杂交组合Bt汕优63进行田间卷叶虫和黄茎螟抗性试验,发现在不降低水稻产量的情况下,转基因杂交稻高抗卷叶虫和黄茎螟。Wang等[16]利用转Bt基因水稻品系KMD1和KMD2与常规水稻品种杂交,并鉴定其后代F1、BC1和F2群体具有抗虫遗传性,其中Bt水稻对螟虫的抗性为显性遗传,易于在杂交水稻生产中利用。高方远等[17]将优质抗稻瘟病水稻恢复系成恢177与转基因水稻Bt明恢63杂交并回交1次,采用PCR分析、试纸条检测和田间抗性鉴定相结合的方法,育成兼具转基因抗虫性和稻瘟病抗性的水稻新恢复系Bt5198。李荣田等[18]、孔梦莹等[19]以水稻空育131为受体,利用根瘤农杆菌介导法创制了转cry1C*和cry2A*基因的抗虫水稻独立系,并对不同生长发育阶段Bt基因表达量进行了测定。随着对转基因抗虫水稻研究的深入,我国于2009年11月通过了对转Bt基因抗虫水稻华恢1号和Bt汕优63的生物安全认证,转Bt基因抗螟虫水稻技术已日趋成熟。华中农业大学已成功将多个Bt基因(cry1Ab/c、cry1C*和cry2A*)导入生产上大面积推广应用的三系杂交稻恢复系明恢63中,并筛选出标记基因已经丢失的抗虫恢复系材料TT51,以其配组的杂交组合抗虫1号和抗虫2号已完成在多个省市的生产性试验[20-21]。崔海瑞等[22]已育成对二化螟、三化螟和稻纵卷螟等鳞翅目害虫具100%抗性的克螟稻,以此为种质初步育成具有推广前景的大批籼稻新品系和新组合,并开始较大面积示范。实践证明,以转Bt基因抗虫水稻材料为供体,以农艺性状优良的水稻品系为受体,通过MAS技术结合农艺性状筛选,可选育出兼具优良农艺性状和抗虫的新品系,这些抗虫新品系均能稳定遗传Bt抗虫基因,并能成功表达Bt蛋白,且具有与供体亲本相当的抗虫能力[20-22]。【本研究切入点】近年来广西水稻螟虫发生面积不断增加,危害加重,造成产量损失增大,某些地区局部爆发螟虫危害产量损失达50%甚至绝收,而目前针对广西抗螟虫杂交稻恢复系选育的研究鲜见报道。【拟解决的关键问题】通过杂交、回交结合MAS技术将转Bt基因水稻中的抗虫基因cry1C*导入广西骨干恢复系中,培育适合广西稻区推广应用的抗螟虫转基因水稻恢复系,为杂交水稻抗虫育种提供参考依据及基础材料。
1 材料与方法
1.1 试验材料
试验于2017—2020年在广西南宁市进行。含抗虫基因cry1C*和与目标基因连锁的抗除草剂基因(bar基因)的抗螟虫转基因材料T1C-19由华中农业大学作物遗传改良国家重点实验室提供。其他水稻材料包括广西杂交稻骨干恢复系亲本广恢998、桂恢1561和桂恢110,以及利用这些恢复系回交转育而成的抗螟虫转基因恢复系材料KH998、KH1561和KH110均由广西农业科学院水稻研究所分子育种研究室提供。
1.2 试验方法
1.2.1 抗螟虫材料回交转育 以转基因材料T1C-19为供体,以广西杂交稻骨干恢复系亲本广恢998、桂恢1561和桂恢110为轮回亲本进行1次杂交、3次回交和连续8次自交,结合MAS和抗虫鉴定获得抗螟虫转基因恢复系材料KH998、KH1561和KH110。
1.2.2 抗螟虫材料及杂交组合的农艺性状考查和抗虫性鉴定 2017年晚季配制杂交组合野香A/KH998和天丰A/KH998,2018年早季以特优63为对照(CK1)种植抗虫转基因杂交组合野香A/KH998和天丰A/KH998。2020年早季配制杂交组合华浙2A/KH110、天丰A/KH1561、华浙2A/KH1561和天丰A/KH110,2020年晚季以天优华占为对照(CK2)种植抗虫转基因杂交组合华浙2A/KH1561、华浙2A/KH110、天丰A/KH1561和天丰A/KH110。试验设2次重复,每份材料种5行,每行种10株,种植密度为18.9 cm×16.5 cm。收获时,取行中间8株考察株高、每株有效穗数、穗长、每穗实粒数、每穗总粒数、结实率、千粒重和单株重等性状,取8株的平均值作为分析数据。
采用室内离体接虫鉴定法结合室外自然鉴定法进行转基因材料抗虫鉴定。室内离体接虫鉴定法具体操作如下:于拔节期从稻株上取茎秆,切成5.0 cm长的茎段,放入养虫管,每管放5个茎段,接入20头一龄三化螟,置于人工气候箱,5 d后统计三化螟死亡率,3次重复。
1.2.3 抗螟虫材料的分子标记鉴定 水稻全基因组DNA通过改良CTAB法[23]抽提获取。根据cry1C*基因的序列设计引物检测水稻基因组DNA中是否含有该基因。参考Tang等[24]的方法进行PCR扩增,扩增引物为F:5′-TTCTACTGGGGAGGACATCG-3′,R:5′-CGGTATCTTTGGGTGATTGG-3′,扩增片段长度为602 bp。PCR反应体系20.0 μL:DNA模板30~50 ng,10×Buffer 2.0 μL,2 mmol/L dNTP 1.5 μL,25 mmol/L MgCl22.0 μL,10 μmol/L引物(F/R)各0.5 μL,加ddH2O至20.0 μL;扩增程序:94 ℃预变性5 min;94 ℃ 1 min,57 ℃ 1 min,72 ℃ 1 min,进行35个循环;72 ℃ 5 min。扩增产物进行1%琼脂糖凝胶电泳检测。
根据bar基因序列设计引物检测水稻基因组DNA是否含有该基因。PCR扩增引物为F:5′-GGGTCTAGAATGGAGCCCAGAACGACGCCG-3′,R:5′-GCTCGGGATCCTCAGATCTCGGTGACG-3′,扩增片段长度为561 bp。PCR反应体系20.0 μL:DNA模板50 ng,10×Buffer 2.0 μL,2 mmol/L dNTP 1.5 μL,25 mmol/L MgCl22.0 μL,10 μmol/L引物(F/R)各0.5 μL,Taq酶1 U,加ddH2O至20.0 μL;扩增程序:94 ℃预变性5 min;94 ℃ 1 min,57 ℃ 1 min,72 ℃ 1 min,进行35个循环;72 ℃ 5 min。扩增产物进行1%琼脂糖凝胶电泳检测。
1.2.4 抗螟虫材料杀虫蛋白含量检测 于黄熟期取参试材料的新鲜绿色叶片检测杀虫蛋白表达量,参照cry1C平板试剂盒(美国Envirologix公司)说明进行检测。具体方法如下:称取20.0 mg水稻新鲜叶片,用试剂盒随带的抽提液进行研磨抽提,在酶标仪450 nm处读取吸光度值,绘制标准曲线,计算样品中的Bt蛋白含量。
1.3 统计分析
试验数据采用Excel 2007进行统计分析。
2 结果与分析
2.1 抗螟虫转基因恢复系的选育历程
抗螟虫转基因恢复系的选育过程见图1。以骨干恢复系(广恢998、桂恢1561和桂恢110)为母本,与T1C-19进行杂交,获得F1代杂种。种植F1植株,根据株叶形态特征去除假杂种,选择真杂种作父本与轮回亲本(骨干恢复系)进行回交,获得BC1F1代种子。种植BC1F1植株,移栽后逐株取叶片进行MAS,选取检测结果为阳性的植株为父本与轮回亲本继续进行回交,获得BC2F1种子;同样在BC2F1中选取阳性单株继续回交获得BC3F1,最终根据分子标记检测和农艺性状考察结果,在BC3F1中收获10~20株阳性植株,收取自交种。将收获的10~20株阳性BC3F2群体各种植100苗,结合分子标记检测结果和农艺性状表现,最后在15个群体中选择30个Bt阳性单株收种。种植来自BC3F2的30个BC3F3株系,每个株系50苗,苗期逐株进行PCR检测,筛选出10个Bt基因纯系。10个Bt基因纯系田间自然鉴定均无螟虫发生,根据农艺性状表现,选择与轮回亲本差异最小的株系,再经过连续5代自交稳定后,正式定名。通过此方法选育出与广西杂交稻骨干亲本广恢998、桂恢1561和桂恢110分别对应的抗螟虫转基因恢复系材料KH998、KH1561和KH110。
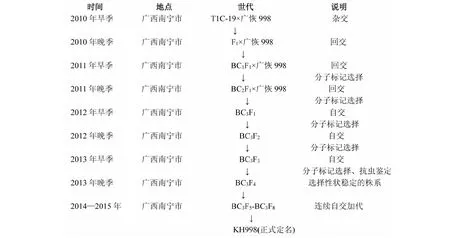
图1 抗螟虫转基因恢复系KH998的选育过程Fig.1 Breeding process of insect-resistant restorer line KH998
2.2 抗螟虫转基因恢复系及其杂交组合的性状表现
2016年早季种植KH998、KH1561和KH110及其原始亲本广恢998、桂恢1561和桂恢110,以T1C-19为对照(CK3),考查农艺性状表现。由表1可知,KH998、KH1561和KH110与对应的原始亲本相比,均对螟虫具有抗性,且抗性与CK3相当;在农艺性状方面较其对应的原始亲本有所改善,如KH998、KH1561和KH110的千粒重均低于CK3,KH1561的千粒重比其原始亲本桂恢1561变小;而KH998和KH110的千粒重较其相应原始亲本的千粒重分别增加1.40和3.40 g,产量优势变大。

表1 T1C-19衍生品系的农艺性状和抗螟虫表现
2018年早季和2020年晚季种植抗螟虫转基因杂交组合、亲本、CK1和CK2,收获后考查其农艺性状表现,结果(表2)表明,天丰A/KH110表现抗虫,其产量比CK2增加1.29%;野香A/KH998和天丰A/KH998表现抗虫,但产量比CK1分别减产8.69%和0.49%;华浙2A/KH1561、天丰A/KH1561和华浙2A/KH110表现抗虫,但产量比CK2分别减产1.89%、3.24%和5.56%。
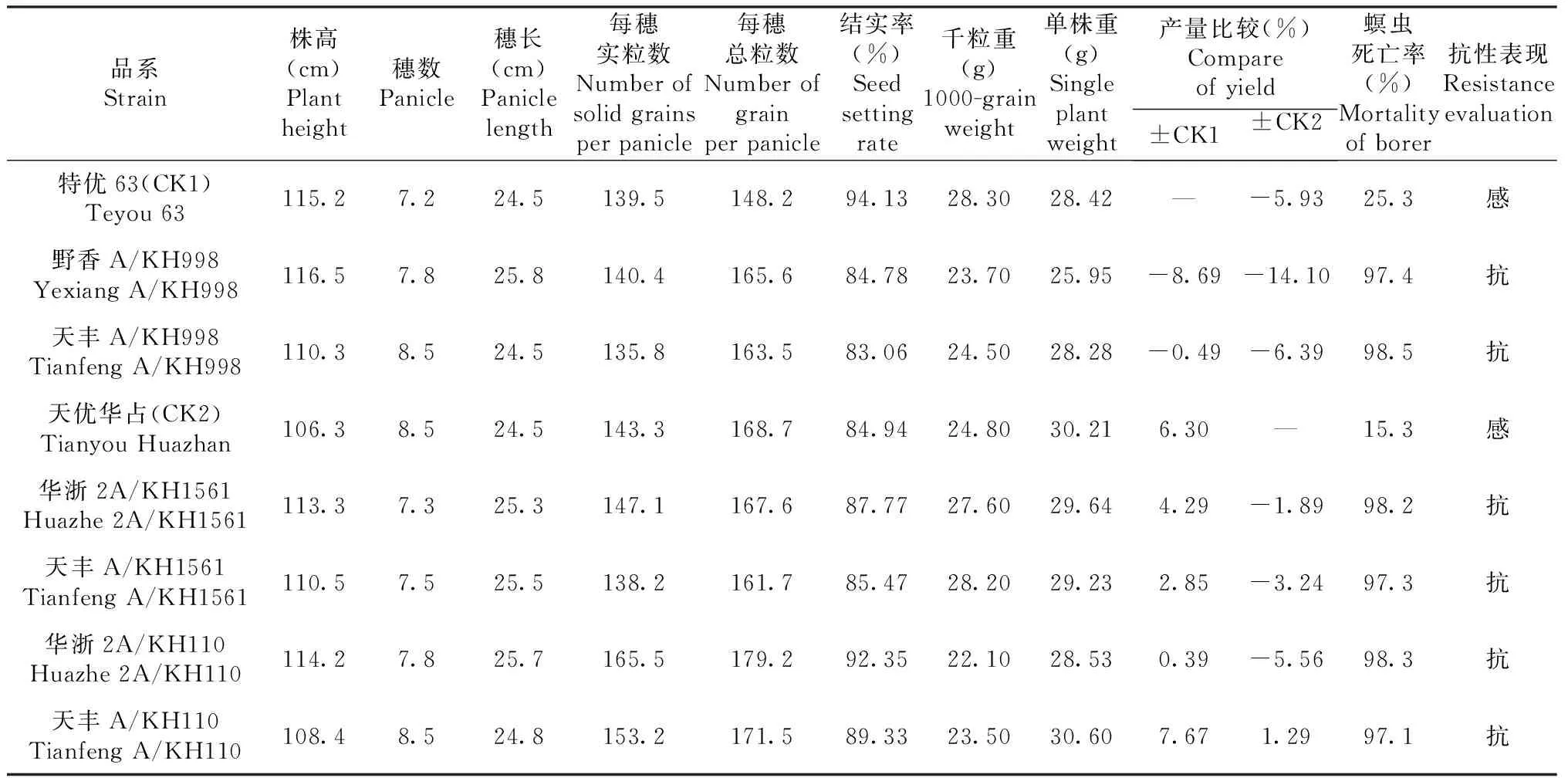
表2 T1C-19衍生品系杂交组合的农艺性状和抗螟虫表现
2.3 抗螟虫转基因恢复系分子标记检测结果
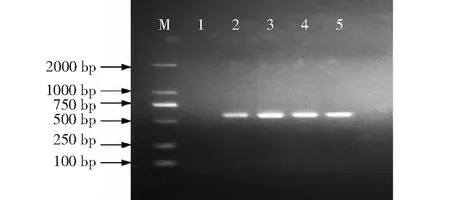
水稻移栽返青后,在KH998、KH1561和KH110及其原始亲本广恢998、桂恢1561和桂恢110中分别随机选取10株的叶片抽提基因组DNA作模板,分别用于与cry1C*基因和bar基因对应的特异引物进行PCR扩增,cry1C*基因在KH998、KH1561和KH110中均能扩增出长度为602 bp的片段,而在阴性对照广恢998、桂恢1561和桂恢110中无条带显示(图2);bar基因在KH998、KH1561和KH110中均扩增出长度为561 bp的片段,而在阴性对照中无条带显示(图3)。说明抗虫基因cry1C*和耐除草剂基因bar均已转入受体亲本中,并能在受体亲本中稳定遗传。

M:DNA Marker;1:T1C-19(阳性对照);2~7分别为KH998、广恢998 (阴性对照)、KH1561、桂恢1561(阴性对照)、KH110和桂恢110 (阴性对照)的单株;目标产物片段为602 bpM:DNA Marker;1:T1C-19(positive control);2-7 were the individual plants of KH998,Guanghui 998 (negative control),KH1561,Guihui 1561(negative control),KH110 and Guihui 110(negative control);the target product size fragment was 602 bp图2 KH998、KH1561和KH110的cry1C*基因PCR检测结果Fig.2 PCR detection results of cry1C*gene in KH998,KH1561 and KH110
M:DNA Marker;1:阴性对照;2:T1C-19(阳性对照);3~5分别为KH998、KH1561和KH110 的单株;目标产物片段为561 bpM:DNA Marker;1:Negative control;2:T1C-19(positive control);3-5 were the individual plants of KH998,KH1561 and KH110;The target product size fragment was 561 bp图3 KH998、KH1561和KH110的bar基因PCR检测结果Fig.3 PCR detection results of bar gene in KH998,KH1561 and KH110
2.4 抗螟虫恢复系的杀虫蛋白含量检测与分析
在KH998、KH1561和KH110的单株中各随机选择30个单株,测定其Bt蛋白的表达量(Bt蛋白含量),结果(图4)显示,在3个群体中,Bt蛋白含量最大值为1.70 μg/g FW,最小值为0.02 μg/g FW,平均为0.77 μg/g FW,单株间Bt蛋白含量存在较大差异;KH998的Bt蛋白含量较总体水平偏低,平均为0.24 μg/g FW,且个体间差异不明显,说明该抗性群体的Bt蛋白能稳定表达;KH1561的Bt蛋白含量平均为0.65 μg/g FW,且个体间差异不明显,说明该抗性群体的Bt蛋白也能稳定表达;KH110的Bt蛋白含量较总体水平偏高,平均为1.41 μg/g FW,个体间差异较明显(0.02~1.70 μg/g FW),说明该抗性群体的Bt蛋白表达不够稳定。

KH998:1~30;KH1561:31~60;KH110:61~90图4 抗性植株中cry1C*杀虫蛋白含量检测结果Fig.4 Detection of cry1C*insecticidal protein content in resistant plants
3 讨 论
前人研究认为,培育抗螟虫水稻新品种才是最经济有效的水稻螟虫防控措施[5],但我国水稻种质资源中抗螟虫材料较少,采用常规育种方法很难培育出抗螟虫水稻品种,而以转Bt基因水稻材料为供体,通过MAS技术结合农艺性状筛选,是选育抗螟虫水稻品种的有效途径[17,20-21]。
已有研究表明,在通过遗传转化获得转基因抗虫水稻的基础上,借助MAS和回交育种手段可改良转Bt基因抗虫水稻中的一些不良性状,选育出既具有高抗螟虫又高产优质特性的水稻新品种或品系[25-27],如高方远等[17]将优质抗稻瘟病水稻恢复系成恢177与转基因水稻Bt明恢63杂交并回交1次,采用PCR分析、试纸条检测和田间抗性鉴定相结合的方法,育成兼具转基因抗虫性和稻瘟病抗性的水稻新恢复系Bt5198;刘艳等[20]将转基因水稻Bt明恢63与水稻三系恢复系先恢207杂交并回交1次,采用PCR分析和田间抗性鉴定相结合的方法,育成抗螟虫水稻新恢复系;田雨等[21]以转基因水稻Bt明恢63为供体,以水稻品种9311为受体,利用MAS技术对9311的螟虫抗性进行改良,进一步筛选出抗螟虫杂交稻组合。这些利用MAS技术进行的水稻抗螟虫育种,既减少了烦琐的遗传转化工作,又能为转基因抗虫育种提供更多优良的亲本材料。本研究以T1C-19为抗虫基因供体,通过回交转育结合MAS技术育成的抗螟虫恢复系KH998、KH1561和KH110,均能稳定遗传cry1C*基因,能成功表达Bt蛋白,且具有与供体亲本T1C-19相当的抗虫能力,能有效抵御螟虫危害,且农艺性状与原始亲本广恢998、桂恢1561和桂恢110相近,所配制的杂交组合野香A/KH998、天丰A/KH998、华浙2A/KH1561、华浙2A/KH110、天丰A/KH1561和天丰A/KH110也具有与供体亲本T1C-19相当的抗虫能力,但产量与对照相当或减产,分析其产量优势不强的原因,一方面选用的都是小粒型、米质优的不育系,所配制杂交组合的千粒重明显较对照偏低,因此产量优势不明显,另一方面也说明杂交组合的特殊配合力不高,下一步需加强杂交组合优势选育方面的研究,在兼顾米质的同时提高产量。
利用MAS和回交育种技术选育的抗螟虫水稻新品系,其抗螟虫性状均得到显著提高,表现抗虫[17-20],但Bt蛋白在不同材料中的表达量不一致,有的能稳定表达,有的表达量差异较明显。本研究育成的3个抗螟虫恢复系中,KH998和KH1561的Bt蛋白含量在个体间差异不明显,说明Bt蛋白能稳定表达;而KH110的Bt蛋白含量个体间差异较明显(0.02~1.70 μg/g FW),说明该抗性群体的Bt蛋白表达不够稳定,与田雨等[21]在育成具有9311背景的抗虫水稻品种时获得同一时期单株间Bt蛋白含量存在较大差异的研究结果相似。可见,通过转育获得的抗螟虫水稻新品系均表现抗虫,但Bt蛋白的表达量可能不一致。因此,在转育抗螟虫水稻新品系时,在获得稳定抗虫性新材料后,建议对其进行Bt蛋白含量的单株测定,以获取Bt蛋白能稳定表达的株系。
4 结 论
通过回交转育结合MAS育成的3个抗螟虫转基因恢复系KH998、KH1561和KH110均可稳定遗传cry1C*基因,成功表达Bt蛋白,且具有与供体亲本T1C-19相当的抗虫能力。
