大蒜全基因组WOX基因家族鉴定及密码子使用偏性分析
2022-04-06董新玉张亚春陈国发李树锋
董新玉,邓 浪,张亚春,陈国发,李树锋
(1.大理州农业科学推广研究院经济作物研究所,云南 大理 671000;2.云南农业职业技术学院,昆明 650031)
【研究意义】大蒜(AlliumsativumL.)是百合科葱属植物,是一种重要的的蔬菜、香料和药用作物,2018年全世界大蒜种植面积高达154.7万hm2,产量达2849.4万t[1]。近年来,随着分子生物学技术的快速发展,越来越多的学者从传统的栽培育种转向从细胞分子层面研究大蒜的生长发育和遗传进化,并在大蒜遗传多样性分析、基因组学、转录组学和基因表达与功能验证等方面获得较多研究成果[2]。目前,大蒜全基因组数据已经公布[3],为加快研究大蒜基因功能和进化提供了更为有效的途径。【前人研究进展】WOX(WUSCHEL-related homeobox)转录因子属于HB(Homeobox)转录因子超家族,是植物中广泛存在的一类转录因子,也是植物所特有的一类转录因子[4]。WOX家族成员在拟南芥、棉花、水稻和番茄等植物中已得到鉴定[4]。其中,拟南芥WOX基因家族包含15个成员[5],棉花中有37个成员,水稻和番茄中分别有12和10个成员[6-8]。Graaff等对不同物种的WOX家族蛋白进行进化分析发现,WOX家族蛋白被分为远古支(Ancient clade)、中间支(Intermediate clade)和WUS支(WUS clade)[5]。功能研究发现,WOX家族蛋白在植物干细胞维持、器官发育、激素信号转导、初生和次生物质代谢及抗逆响应等过程中发挥着重要的调节作用[5,9-11]。有研究指出,密码子具有简并性,在遗传信息的传递过程中发挥着重要的作用[12]。在长期进化过程中,不同物种会形成独特的适应自身基因组环境的密码子使用偏性(Codon usage bias,CUB)[13]。密码子使用偏性会随着基因组中大部分基因或者特定基因的变化而改变[14]。研究表明,密码子偏好性分析能为预测基因功能、基因的表达水平、选择基因异源表达受体系统和优化密码子以提高异源表达效率等提供参考[13]。【本研究切入点】本研究利用生物信息学分析方法,对大蒜WOX家族蛋白进化关系、密码子使用偏性及表达进行分析。【拟解决关键问题】以期进一步为研究大蒜WOX基因在大蒜生长发育过程中的作用提供科学依据。
1 材料与方法
1.1 数据来源
大蒜全基因组数据从https://doi.org/10.6084/m9.figshare.12570947.v1下载[3],拟南芥全基因组WOX基因家族信息从TAIR(http://www.arabidopsis.org/)中下载[15],水稻、番茄、辣椒和猕猴桃全基因组WOX基因家族信息从iTAK(http://itak.feilab.net/cgi-bin/itak/db_home.cgi)中下载。大肠杆菌、酵母菌和拟南芥的基因组密码子使用偏性数据从Kazusa DNA研究所Codon Usage Database在线数据库(http://www.kazusa.or.jp/codon/)下载[16]。
1.2 方法
1.2.1 大蒜WOX基因家族的鉴定与分析 利用Sun等[3]2020年公布的大蒜全基因组数据,以大蒜全基因组蛋白序列和核酸序列以及拟南芥WOX基因家族序列构建FASTA文本,并利用BioEdit软件构建大蒜全基因组本地数据库,同时以构建的拟南芥WOX基因家族为探针,进行本地BLAST,重复上述操作去冗余后,结合NCBI(National Center for Biotechnology Information)、SMART(http://smart.embl-heidelberg.de/)和MEGA 6.0软件进行结构域和同源性分析,对大蒜WOX基因家族进行鉴定筛选。同时利用MEGA 6.0和DNAMAN 6.0软件对大蒜WOX基因家族进行系谱进化分析及保守结构域分析[17]。
1.2.2 密码子偏好性分析 利用CodonW软件和EMBOSS(http://emboss.bioinformatics.nl/cgi-bin/emboss/cusp)在线软件计算大蒜WOX基因家族编码蛋白的密码子偏性参数,并用SPSS 19.0、Excel 2010和MEGA 6.0进行大蒜WOX基因家族的密码子碱基组成与GC偏好性(GC3s)的相关性、整体RSCU值、系统聚类及最佳外源受体表达系统选择分析,并参照牛元[18]和Sueoka[19]等使用方法进行最优密码子、ENC-Plot和PR2-plot分析,分析大蒜WOX基因家族的密码子使用偏性强弱及受突变压力或选择压力的影响,同时用Origin 2017、RStudio、MEGA 6.0和TBtools软件作图。
1.2.3 大蒜WOX家族表达谱分析 利用Sun等[3]上传于GEO数据库(登录号:GSE145455)中的表达数据,提取大蒜WOX基因家族在大蒜鳞茎膨大8个时期(鳞茎未膨大开始,每隔5 d取样1次)、假茎、根、叶的表达数据,并用TBtools软件作图。
2 结果与分析
2.1 大蒜WOX基因家族鉴定及进化分析
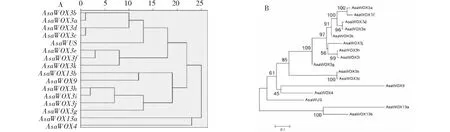
通过本地BLAST去冗余后,最终从大蒜基因组数据库中获得16个WOX家族成员,根据NCBI同源比对及构建进化树中与拟南芥WOX基因家族的同源关系(图1-A),分别将这些成员命名为AsaWUS、AsaWOX3a、AsaWOX3b、AsaWOX3c、AsaWOX3d、AsaWOX3e、AsaWOX3f、AsaWOX3g、AsaWOX3h、AsaWOX3l、AsaWOX3j、AsaWOX3k、AsaWOX4、AsaWOX9、AsaWOX13a和AsaWOX13b。进一步将大蒜、拟南芥、番茄、辣椒、水稻和猕猴桃WOX基因家族做进化分析发现(图1-B),5种不同植物的WOX家族成员被划分为WUS支(WUS clade)、中间支(Intermediate clade)和远古支(Ancient clade)。5种植物均含有WUS支、中间支和远古支成员。大蒜WOX家族16个成员中,WUS支包含:AsaWUS、AsaWOX3a、AsaWOX3b、AsaWOX3c、AsaWOX3d、AsaWOX3e、AsaWOX3f、AsaWOX3g、AsaWOX3h、AsaWOX3l、AsaWOX3j、AsaWOX3k和AsaWOX4;中间支包含:AsaWOX9;远古支包含:AsaWOX13a和AsaWOX13b。5个物种的WOX家族成员在3个分支中均有分布,说明WOX家族的分化在大蒜、拟南芥、水稻、番茄、辣椒和猕猴桃物种分化之前。
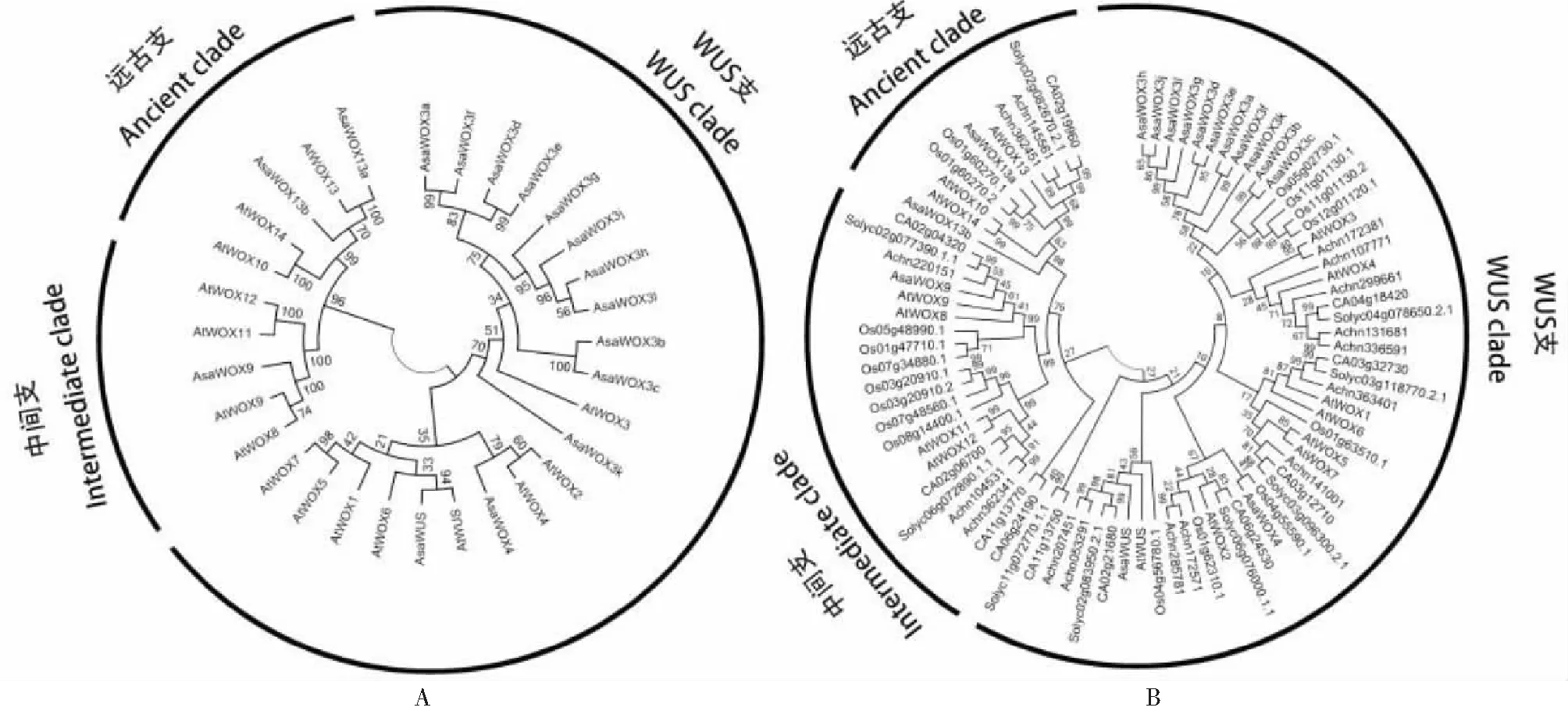
A为大蒜与拟南芥WOX基因家族的进化关系分析;B为大蒜、拟南芥、番茄、辣椒、水稻和猕猴桃WOX基因家族的进化关系分析A is evolution analysis of WOX genes family between garlic and Arabidopsis;B is evolution analysis of WOX genes family among garlic, Arabidopsis, tomato, pepper, rice and kiwi fruit图1 大蒜WOX基因家族鉴定及进化分析Fig.1 Identification and evolution analysis of garlic WOX genes family
2.2 大蒜WOX家族成员保守结构域分析
利用DNAMAN 6.0对大蒜WOX家族成员蛋白序列进行多序列比对,结果表明,大蒜WOX家族成员大多含有68个氨基酸组成的“螺旋—环—螺旋—转角—螺旋”HOX保守结构域(图2)。其中,AsaWOX3k保守结构域不完整,AsaWOX13a、AsaWOX13b、AsaWOX9、AsaWOX3g、AsaWOX3h、AsaWOX3l和AsaWOX3j保守结构域序列较其他成员有较大差异,预示它们可能在功能上存在较大差异。

图2 大蒜WOX家族成员保守结构域分析Fig.2 Analysis of conserved domains of garlic WOX family members
2.3 大蒜WOX家族成员密码子使用偏性分析
2.3.1 密码子偏性相关参数分析 分析了大蒜WOX家族成员碱基组成及相关密码子偏性参数(表1),其中平均AU含量和GC含量分别为58.02%和41.98%,密码子第3位碱基平均含量A3s>T3s>G3s>C3s,GC1s、GC2s和GC3s平均含量分别为0.450、0.414、0.395,均小于0.5,揭示大蒜WOX家族成员编码序列密码子中AU使用频率高于GC。CBI均值为-0.038,Fop均值为0.407,ENC平均值为50.53,偏向61,CAI均值为0.186,远低于1.0,说明大蒜WOX家族成员密码子使用偏性较弱。
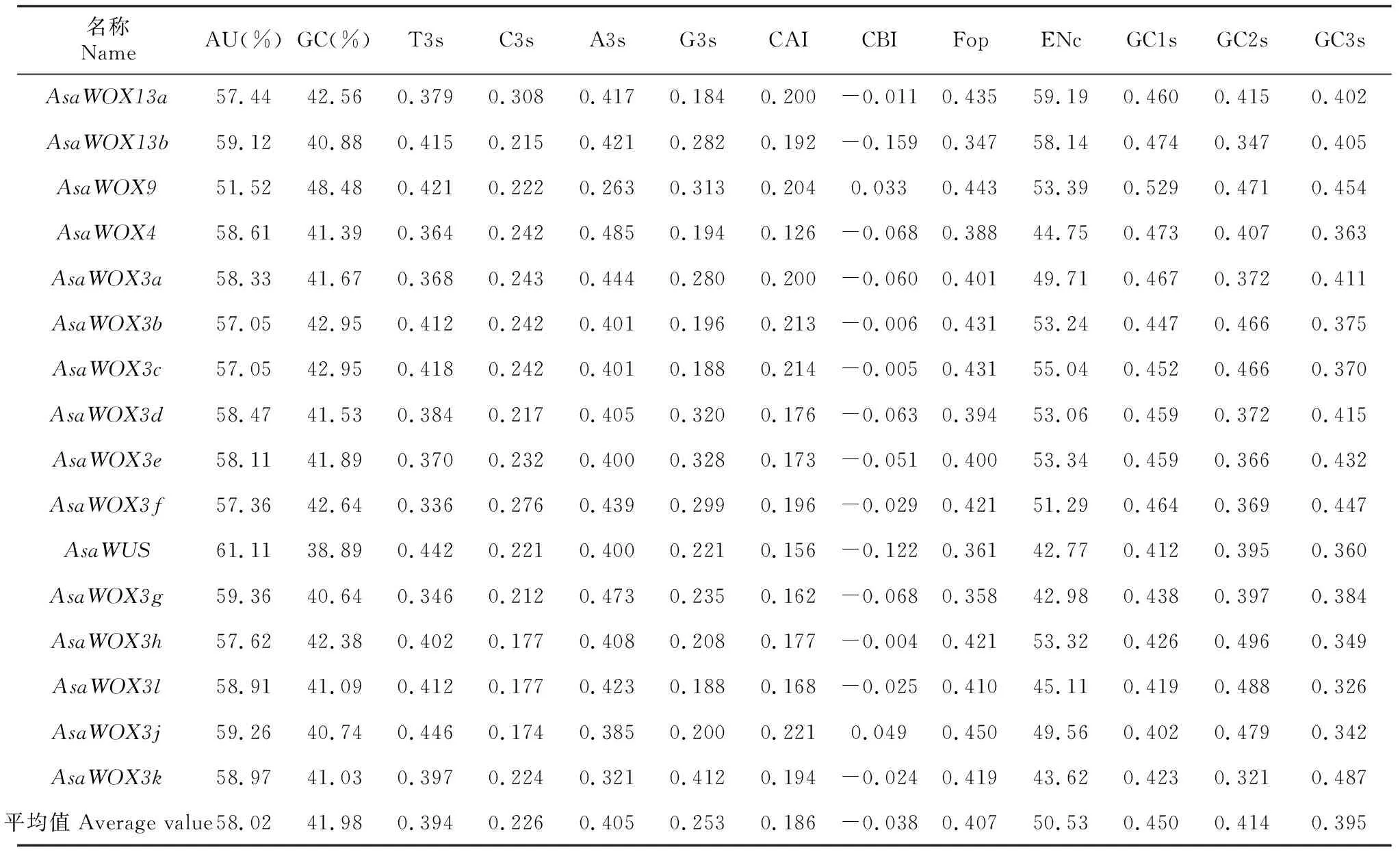
表1 大蒜WOX家族成员碱基组成及相关密码子偏性参数
2.3.2 大蒜WOX成员密码子与GC(GC3s)偏好性的关系 分析大蒜WOX家族成员密码子与GC(GC3s)偏好性的关系,结果(图3)表明,以G/C结尾的密码子与GC3s偏好性表现正相关,多数以A/U结尾的密码子与GC3s偏好性表现负相关,但CUU、UUU、ACU、AGU、AAU、CCA、GCA、GGA、AUA与GC3s偏好性表现正相关,表明它们受GC3s偏好性影响。
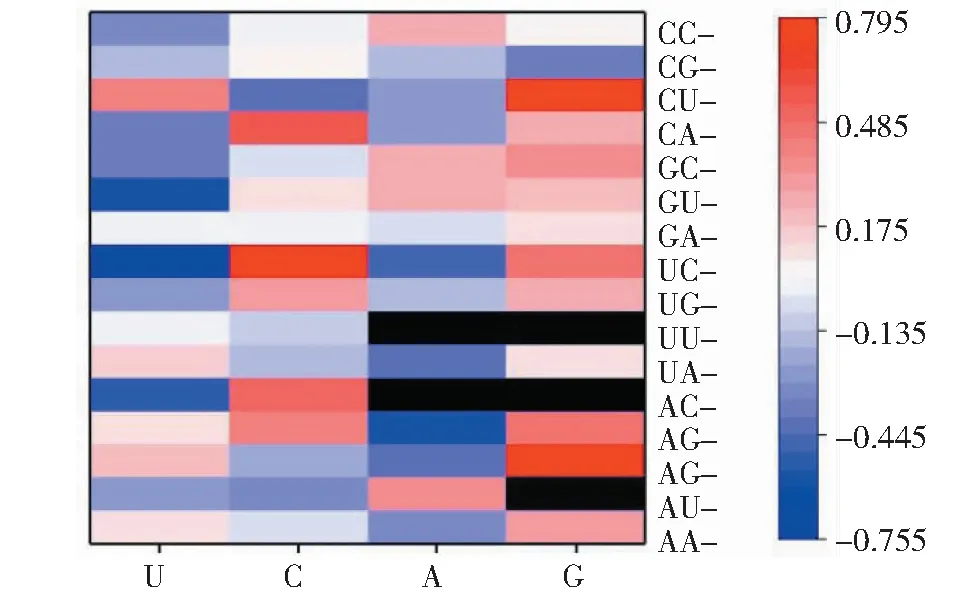
红色代表正相关,蓝色代表负相关,黑色表示终止密码子(UAA、UAG、UGA)和起始密码子(AUG、UGG)Red represents positive correlation, blue represents negative correlation, and black represents stop codons (UAA, UAG, UGA) and start codons (AUG, UGG)图3 蒜WOX家族密码子与GC3s相关性Fig.3 The correlation analysis between codons and GC3s of garlic WOX family
2.3.3 大蒜WOX基因家族整体RSCU值、最优密码子及聚类分析 大蒜WOX家族整体RSCU值分析发现,大蒜WOX家族59个密码子〔除终止密码子(UAA、UAG、UGA)和起始密码子(AUG、UGG)〕中,存在27个高频密码子(RSCU>1表示使用频率较高的密码子,表中以*表示)。其中,只有7个密码子以G/C结尾(AGG、UGC、CAC、UUG、UUC、AGC和GUG),20个密码子以A/U结尾,表明大蒜WOX家族成员偏好A/U结尾密码子。
大蒜WOX家族成员RSCU值聚类分析显示(图4),大蒜WOX家族成员被划分为2大分支,聚为同一支成员间对密码子的使用偏性具有相似性,聚为同一支上的密码子,在不同成员间,其RSCU值大小也基本相似,也说明成员之间对密码子使用偏性基本一致。
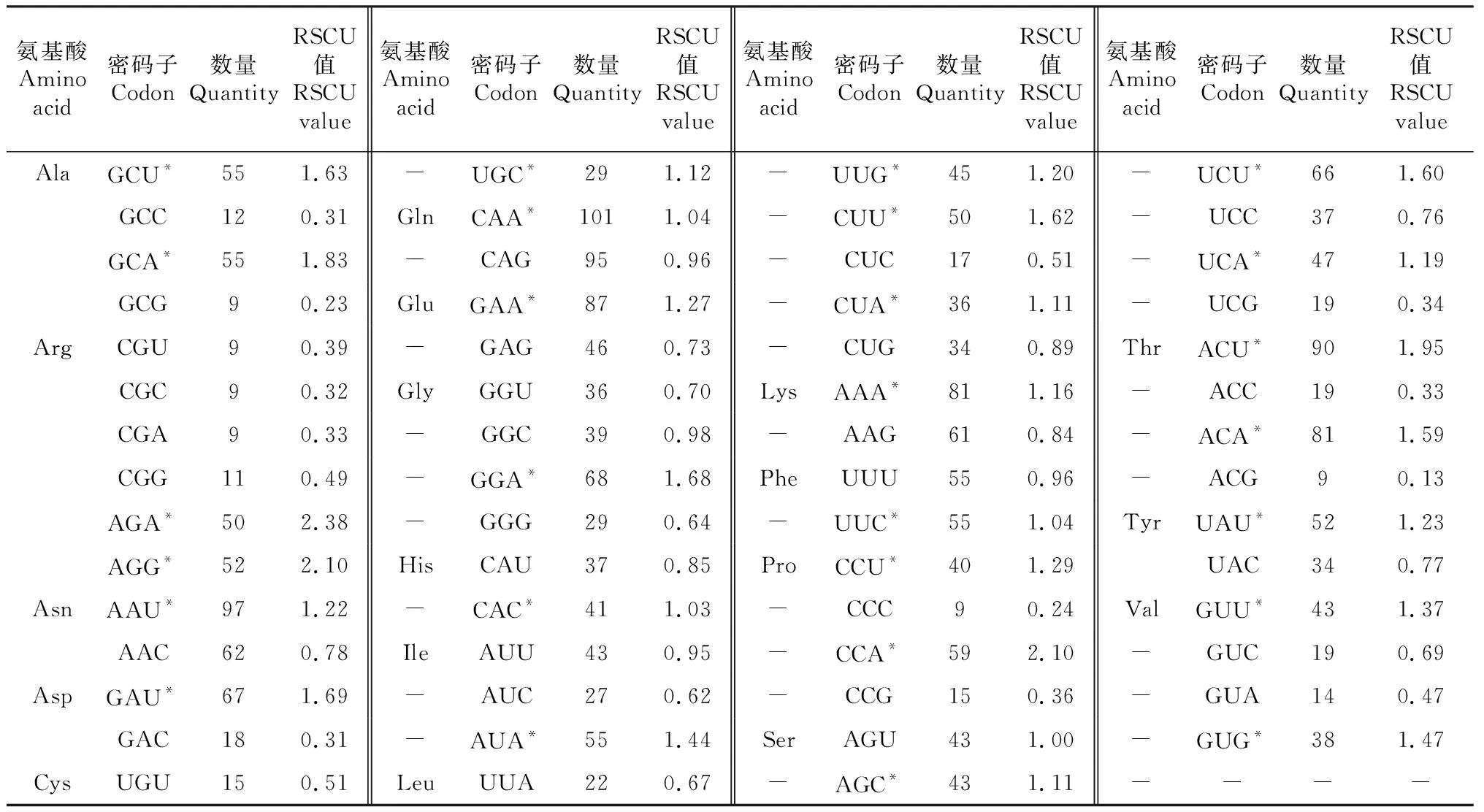
表2 大蒜WOX基因家族整体RSCU值

图4 大蒜WOX家族成员RSCU聚类分析Fig.4 RSCU cluster analysis of garlic WOX family members
RSCU>1的密码子确定为高频密码子,△RSCU值大于0.08即为高表达密码子,同时满足高频密码子与高表达密码子即为最优密码子[18]。最优密码子分析发现(图5),大蒜WOX家族存在14个最优密码子,只有UUC和GUG以G/C结尾,其余12个最优密码子均以A/U结尾,进一步表明大蒜WOX家族成员偏好A/U结尾密码子。
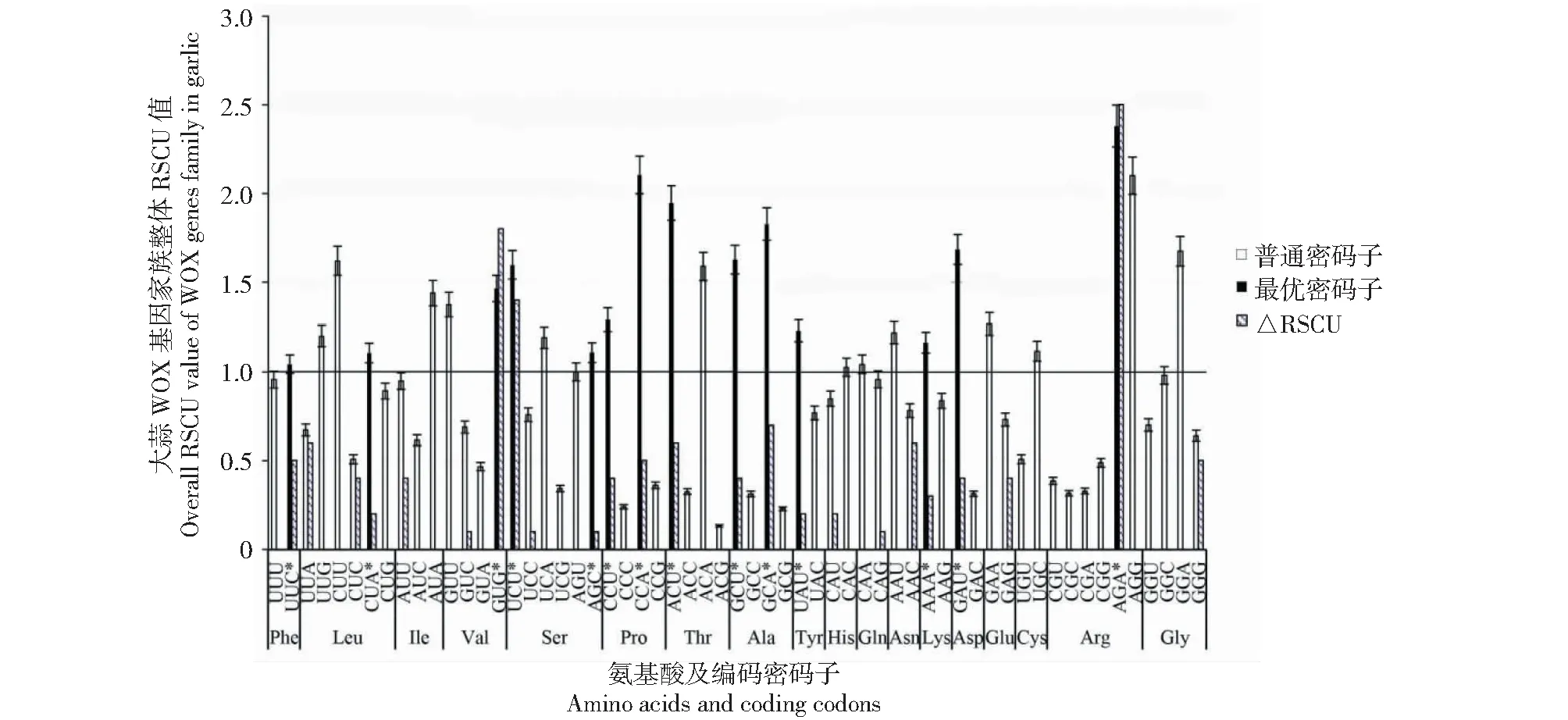
△RSCU<0在图中未显示;*代表最优密码子△RSCU<0 is not shown in the figure; * represents the best codon图5 大蒜WOX家族最优密码子分析Fig.5 Optimal codon analysis of garlic WOX family
2.3.4 ENC-Plot和PR2-plot分析 GC3s分布范围反映了植物所受的选择压力,GC3s分布范围越小,表明密码子使用偏性受自然选择压力越大,同时当4种碱基使用频率一致时,密码子偏性仅受突变压力的影响[20]。此外,绘制理论ENC值标准曲线可反映密码子受碱基突变或选择压力的影响,实际值在曲线上,表示密码子仅受碱基突变的影响,在标准曲线下方,表示密码子受选择压力的影响[18]。ENC和GC3s关联分析显示(图6-A),大蒜WOX家族成员GC3s分布范围在0.326~0.487,分布较为集中,说明该家族成员密码子使用偏性受选择压力影响较大。此外,该家族成员基本分布在标准曲线下方,有2个成员分布在标准曲线上,说明该家族成员在进化过程中大多受选择压力的影响,少数成员受碱基突变压力的影响。PR2-plot分析显示(图6-B),大蒜WOX家族成员的A3/(A3+G3)或G3/(G3+C3)值基本都偏离0.5,进一步表明该家族成员密码子使用偏性主要受选择压力的影响。
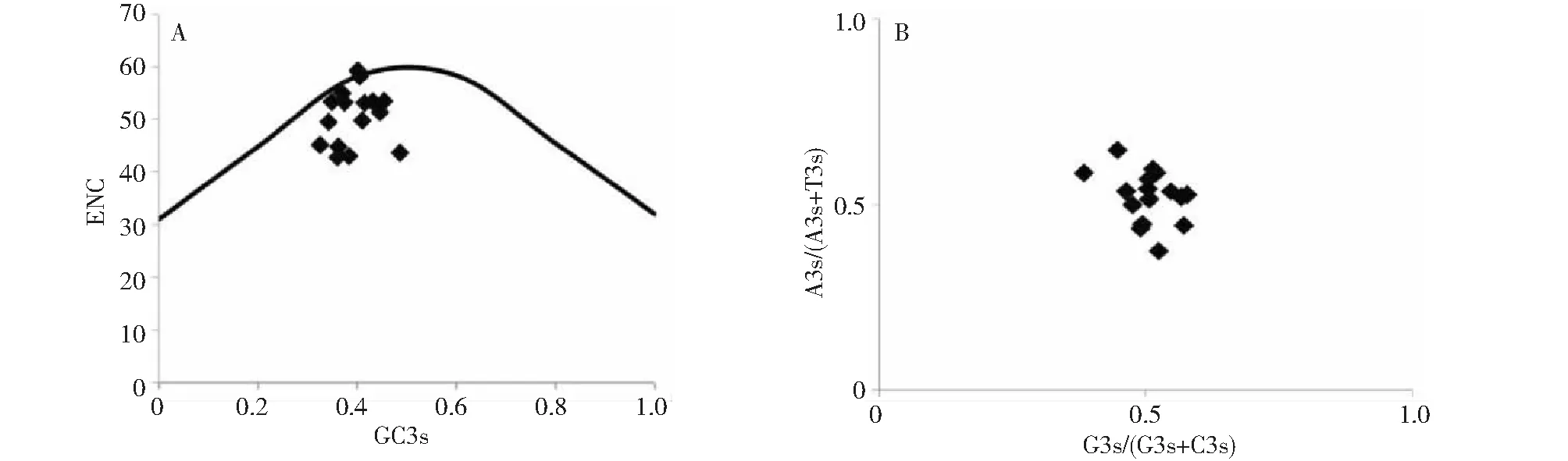
标准曲线绘制公式为ENC=2+GC3s+29/(GC3s2+(1-GC3s)2)[18]The standard curve drawing formula is ENC=2+GC3s+29/ (GC3s2+ (1-GC3s) 2)[18]图6 大蒜WOX家族成员ENC-Plot和PR2-plot分析Fig.6 ENC-Plot and PR2-plot analysis of garlic WOX family members
2.3.5 大蒜WOX家族成员RSCU值和CDS序列聚类分析 一般功能相似性高的基因,往往表现出类似的密码子使用模式[21]。RSCU值和CDS序列聚类分析结果表明,基于RSCU值和CDS序列聚类均将大蒜WOX家族16个成员划分为2大类,但是2种聚类存在较大差异。基于RSCU值聚类(图7-A),表明同一分支上的成员间,具有类似的密码子使用模式;而基于CDS序列聚类(图7-B),表明同一分支上的成员间,同源性高、亲缘关系更近。也说明基于CDS序列的聚类分析相比较基于RSCU值的聚类分析要更加准确,更能说明WOX家族成员之间的进化关系,而基于RSCU值的聚类分析可作为前者的补充。
A为RSCU值聚类;B为CDS序列聚类A is RSCU values cluster analysis; B is CDS sequences cluster analysis图7 大蒜WOX家族成员RSCU值和CDS序列聚类分析Fig.7 RSCU values and CDS sequences cluster analysis of garlic WOX family members
2.3.6 大蒜WOX基因家族外源转化受体系统选择 选择最佳受体表达系统可通过物种间密码子使用偏性差异来表示。而物种间密码子使用偏性差异则通过密码子使用频率比值来进行衡量。2个物种密码子使用偏性较一致,说明它们之间密码子使用频率比值介于0.5~2,相反,表示2个物种密码子使用偏性差异较大[16,22]。大蒜WOX基因家族最佳外源转化受体系统选择分析显示(表3),不同物种间密码子使用偏好性差异不同(表中下划线标出)。大蒜与大肠杆菌存在密码子使用偏好性差异较大的密码子有17个,与酵母菌有11个,与拟南芥有6个。表明拟南芥较适用于大蒜WOX基因家族的外源表达系统。
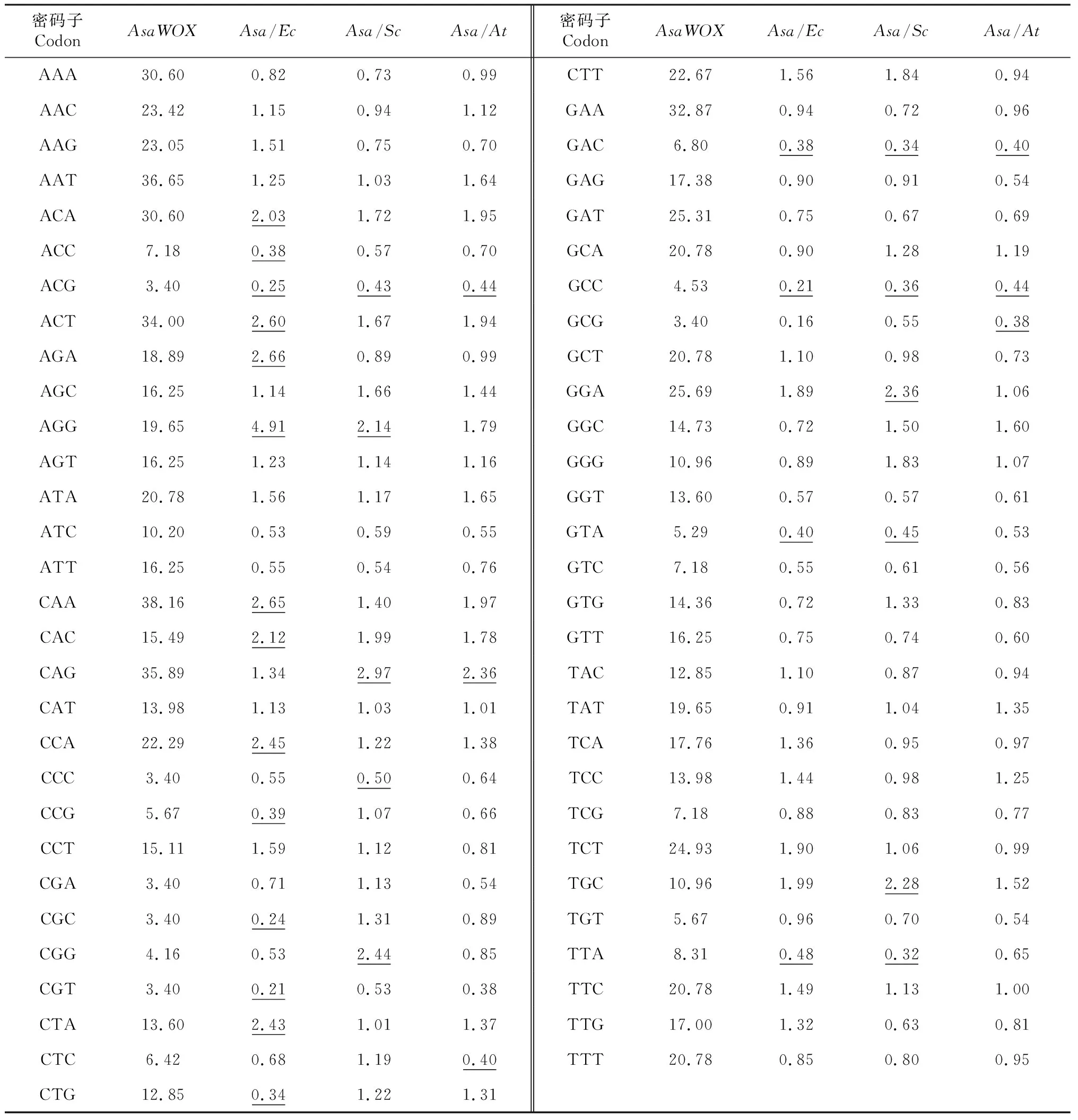
表3 大蒜WOX基因家族最佳外源转化受体系统选择
2.4 大蒜WOX基因家族表达分析
从GEO数据库中下载获取大蒜全基因组的表达数据,并抽取大蒜WOX基因家族在鳞茎发育8个时期(LJ-1~LJ-8,鳞茎发育8个时期分别为:从鳞茎未膨大至鳞茎未膨大完成,每个时期间隔5 d)、叶片(YP)、假茎(JJ)和根(G)中的表达数据做表达热图(AsaWUS、AsaWOX3a、AsaWOX3b、AsaWOX9、AsaWOX13a表达数据为0,未在图中展示),结果(图8)表明,AsaWOX13b在LJ-1、LJ-5、LJ-8和LJ-7中表达,以LJ-5中表达量最高;AsaWOX4在LJ-1和叶片中表达,以叶片中表达量最高;AsaWOX3c在LJ-7、叶片、假茎和根中表达,以假茎中的表达量最高;AsaWOX3d在LJ-3和LJ-8中表达,以LJ-8中表达量最高;AsaWOX3e和AsaWOX3f在叶片和假茎中表达,以叶片中表达量最高;AsaWOX3g在LJ-2和根中表达,以LJ-2中表达量最高;AsaWOX3h在LJ-6和假茎中表达,以假茎中表达量最高;AsaWOX3l只在叶片中表达;AsaWOX3j只在根中表达;AsaWOX3k只在LJ-8表达。
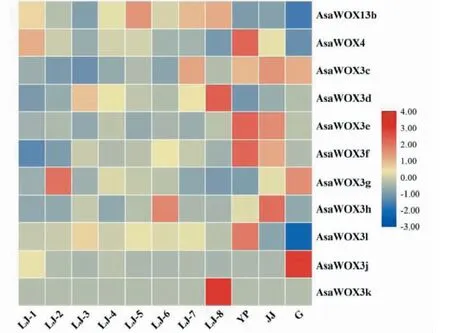
LJ-1~LJ-8表示鳞茎发育8个时期;YP代表叶片;JJ代表假茎;G代表根LJ-1 to LJ-8 represent 8 stages of bulb development; YP represents leaf; JJ represents pseudostem; G represents root图8 大蒜WOX家族成员在不同组织中的表达Fig.8 The expression of garlic WOX family members in different tissues
3 讨 论
3.1 大蒜WOX基因家族鉴定与分析
植物WOX家族在植物生长发育、干细胞维持、激素信号转导、初生和次生物质代谢及抗逆响应等过程中发挥重要调节作用[5]。大蒜基因组数据的公布,为研究大蒜WOX家族提供了重要参考。通过研究从大蒜基因组数据中鉴定得到16个大蒜WOX家族成员,前人研究也表明不同物种间WOX家族成员个数存在差异[拟南芥(15个)、辣椒(11个)、番茄(10个)、猕猴桃(16个)、水稻(17个)和棉花(37个)[4-5,10]],这可能是不同物种在长期进化过程中,为适应生存环境的变化,形成适应自身发展基因组而导致的。进化分析进一步发现,大蒜和水稻亲缘关系更近,拟南芥、番茄、辣椒和猕猴桃亲缘关系更近,且不同植物WOX家族成员在3个分支中均有分布,说明WOX家族成员分化时间要早于这些物种的分化时间,这与在其他植物上的研究结果类似[11,23]。保守结构域分析发现,亲缘关系近的序列,它们之间具有相似的保守结构域,说明鉴定和进化分析结果是可靠的。此外,大蒜WOX家族成员保守结构域由68个氨基酸组成,而星油藤、猕猴桃由60个氨基酸组成[10-11],拟南芥、桂花和乌梅等则由61个氨基酸组成[24],存在这种差异,说明WOX家族在不同物种之间可能经历了不同的分化过程。
3.2 大蒜WOX家族成员密码子使用偏性分析
大蒜WOX家族成员密码子偏性参数、整体RSCU值、最优密码子分析发现,大蒜WOX家族成员偏好使用A/U结尾的密码子,与GC含量偏性相关性分析发现,该家族成员密码子使用偏性与GC含量呈正相关,而该家族成员GC含量低于AU含量,说明大蒜WOX家族成员密码子使用偏性较弱,这与单子叶植物偏爱使用G/C结尾的密码子,双子叶植物偏爱使用A/U结尾的密码子报道不符[25],但是与在菠萝和马尾松等单子叶植物上的研究结果类似[26-27],说明大蒜在进化上可能较其他单子叶植物更接近双子叶植物,进化程度可能较其他单子叶植物高。此外,RSCU值聚类分析发现,具有类似RSCU值的成员聚为一支,暗示它们在功能上可能存在一定的相似性。ENC-Plot和PR2-plot分析表明大蒜WOX家族成员多数受选择压力的影响,少数受突变压力的影响,这与在谷子、月季上的研究结果类似[13,18];RSCU值和CDS序列聚类分析发现,2种聚类存在差异,这与在其他物种上的研究结果类似[12,16,28];说明基于CDS序列聚类更能反应基因间进化,而基于RSCU值一定程度上可作为前者的补充。最佳外源表达系统分析发现,拟南芥更适合作为大蒜WOX家族成员的外源表达系统,这与在苋菜上的研究结果类似,和忍冬、大麦上的研究结果不同[12,28-29],说明不同物种密码子使用偏性不同,在选择外源表达系统时也存在差异,同时说明大蒜WOX家族成员在外源表达时,要高效率异源表达,需对其部分密码子进行优化改造。表达谱分析发现,大蒜WOX家族成员在大蒜鳞茎发育过程、叶片、假茎和根中均有表达,说明WOX家族成员可能参与鳞茎、叶片、假茎和根的生长发育调控, 建议后期进行差异基因的克隆与遗传转化分析,以确定其在特定发育时期的功能作用。
4 结 论
利用生物信息学方法从大蒜基因组中鉴定获得16个WOX基因家族,大部分WOX基因家族含68个氨基酸组成的保守结构域,密码子使用偏性均较弱,且拟南芥更适合用于该基因家族的外源表达受体。此外,大蒜WOX基因家族在大蒜鳞茎不同发育时期及其他组织中差异性表达,这为进一步研究大蒜WOX基因家族在大蒜生长发育过程中的功能奠定了基础。
