高压纳秒脉冲电场的细胞器生物电效应综述*
2022-03-30郭雨怡石富坤王群季振宇庄杰
郭雨怡 石富坤 王群 季振宇 庄杰†
1) (中国科学技术大学生物医学工程学院(苏州)生命科学与医学部,苏州 215000)
2) (中国科学院苏州生物医学工程技术研究所,苏州 215000)
3) (空军军医大学军事生物医学工程学系,西安 710032)
1 引言
高压脉冲电场(pulsed electric fields,PEFs)能够在毫秒到皮秒的时间尺度内以极高的瞬时功率(可达百兆瓦量级)将电场能量施加到生物细胞和组织上,产生多种非线性生物效应,其中最典型的是细胞膜对亲水性大分子的通透性显著增加,即电穿孔(electroporation,EP).毫秒级和微秒级的PEFs 已经被广泛应用于污水处理[1,2]、食品保鲜[3,4]、生物育种[5,6]、肿瘤电化学治疗[7-10]和软组织非热消融[11,12]等领域.脉冲宽度更窄、场强更高的高压纳秒脉冲电场(nanosecond pulsed electric fields,nsPEFs)不仅能够导致细胞膜穿孔,还可以穿透细胞膜直接作用于细胞内部,造成细胞器膜穿孔[13]、细胞骨架解聚[14]、胞内钙离子释放[15]、DNA 损伤[16]等效应,在诱导癌细胞凋亡[17,18]、促进干细胞分化[19,20]以及神经调控[21-23]等领域展现出广阔的应用前景.
20 世纪50 年代,Stampfli[24]在实验中观察到高压电场作用后细胞膜对亲水性大分子的通透性显著增加.1982 年,Neumann 等[25]将PEFs 作用下细胞膜通透性增加的现象称为EP.EP 的产生机理是PEFs 对细胞膜充电,导致跨膜电压超过磷脂双分子层介电击穿的阈值(约1 V),细胞膜生成大量缺陷,缺陷处的磷脂分子发生倒转,从而在细胞膜上产生亲水性微孔[26,27].图1 为细胞膜EP 的结构示意图,其中磷脂分子亲水性的一端朝外,亲油性的一端朝内.

图1 PEFs 导致的细胞膜EP 示意图Fig.1.Schematic diagram of PEF-induced membrane electroporation.
依据细胞膜上的穿孔恢复与否可进一步将EP 划分为可逆电穿孔(reversible electroporation,RE)和不可逆电穿孔(irreversible electroporation,IRE)[12,28].RE 指PEFs 作用后细胞膜上的微孔在细胞活性未受影响前闭合,已用于药物分子和基因导入[29];IRE 指PEFs 作用后细胞膜上微孔无法闭合,导致细胞内物质泄漏,对细胞造成不可逆的伤害,已被应用于实体肿瘤消融等领域[30-32].
为了研究nsPEFs 与细胞膜和细胞内结构相互作用的电学机制,Schoenbach 等[13]针对处于电极之间的真核细胞建立了如图2 所示的等效电路模型.该等效电路模型包含了最主要的细胞结构,即细胞膜、细胞质和细胞核膜及细胞核质.细胞质和细胞核质含有丰富的离子从而具有明显的电阻特性,膜结构由于磷脂双分子层内部的疏水性而具有电容特性[33].由于核膜是双层膜而细胞膜为单层膜,可简化认为核膜电容量约为细胞膜电容的一半,相应的核膜充电时间常数较细胞膜充电时间常数更大[34].因此短于细胞膜充电时间常数的nsPEFs可能造成核膜[35]等细胞器膜产生电穿孔,导致线粒体[36-38]、内质网[39,40]、溶酶体[41,42]等细胞器的结构和功能改变,进一步引发钙离子浓度增加、活性氧产生、细胞凋亡机制激活等次级效应,以上细胞器生物电效应也被称为细胞内电处理效应(intracellular electromanipulation,IEM)[43-45].IEM是nsPEFs 与传统微秒、毫秒PEFs 生物电效应的显著区别.据本文作者调研,目前尚未有专门针对nsPEFs 的细胞器生物电效应的综述报道.因此本文就nsPEFs 影响细胞器的功能结构变化及作用机制的研究展开综述.

图2 PEFs 中哺乳动物细胞等效电路模型Fig.2.Equivalent circuit model for the mammalian cells with the exposure to PEFs,where individual subcellular components are described by a combination of the resistor or capacitor.
为了确保充足的参考文献,本文的搜索基于以下电子数据库:PubMed (美国国家医学图书馆,http://www.ncbi.nlm.nih.gov/PubMed); Scopus Elsevier (http://www.scopus.com/);科学网(Thomson Reuters,http://apps.webofknowledge.com/);Google Scholar (https://scholar.google.com/).本文参考文献主要包括从2000 年起至今所发表的nsPEFs对细胞器作用的相关文献.
图3 给出了收集到的nsPEFs 的细胞器生物电效应的发文量,自2000 年至今,发文量随着时间呈现明显的上升趋势,说明近年来nsPEFs 的细胞器生物电效应的研究吸引了学者们更多的关注.
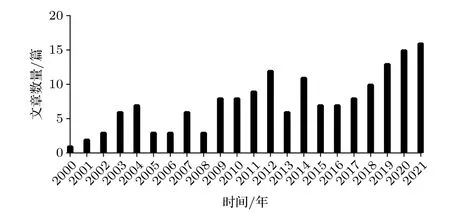
图3 NsPEFs 细胞器效应的文献发表情况Fig.3.Literature research about the intracellular effects for cells exposured to nsPEFs.
文章具体检索包括关键词及其组合的中英文,以及领域内重要研究者名单.中文平台搜索关键词为:(nsPEF OR 纳秒脉冲电场 OR 超短脉冲电场OR 陡脉冲电场 OR 亚微秒脉冲电场)AND(细胞器 OR 细胞内 OR 细胞骨架 OR 线粒体 OR 内质网 OR 囊泡 OR 液泡 OR 核膜 OR 细胞核 OR溶酶体 OR DNA OR 钙离子 OR 蛋白质);英文搜索关键词为:(nsPEF OR nanosecond OR ultrashort OR submicrosecond OR electrical pulse)AND (organelles OR intracellular OR cytoskeleton OR mitochondria OR endoplasmic reticulum OR vesicle OR vacuole OR nuclear OR lysosome OR nucleus OR DNA OR calcium OR proteins) OR nanoelectroporation OR supraelectroporation.为尽量减少遗漏业内重要研究者的研究,在关键词检索搜集到的文章中,找到本研究领域内引文量高的文章,并对作者及同机构其他学者的文章再进行检索筛选;最后,在Google Scholar搜索引擎中对上述关键词再进行检索,找到遗漏的文章.通过以上的文献检索方法尽可能确保文献收集工作的准确性和完整性.
2 高压纳秒脉冲电场的细胞器生物电效应
如上节所述,细胞膜、细胞核、细胞器等亚细胞结构有特定的介电性质,从而可以建立细胞的等效电路模型以描述nsPEFs 的细胞器生物电效应.除了基于充电时间常数的描述,米彦[46]基于等效电路和单细胞壳层介电模型仿真在频域分析细胞膜和核膜跨膜电位,得到细胞的频率响应特性,发现当PEFs 携带频率超过1 MHz 的分量时即可影响细胞核膜.
针对nsPEFs 的细胞器生物电效应的相关研究采用了脉宽从几纳秒到几百纳秒,场强从几kV/cm 到几百kV/cm 的PEFs 参数,作用对象包括来源于人和动物多种器官的癌细胞或正常细胞.该脉宽范围包含了几MHz 到几百MHz 的频率分量,从而可以影响细胞器.相关研究文献所用PEFs参数和观察到的细胞器变化[15-18,34,36,39-42,46-101]总结于表1,从表1 可以看出大多数研究关注细胞骨架、线粒体、内质网、细胞核等亚细胞结构.因此本文将依次分类阐述nsPEFs 与上述各细胞器的相互作用,最后综合讨论nsPEFs 对其他亚细胞结构的效应.

表1 nsPEFs 的细胞器生物电效应总结Table 1.Summary of effects of nsPEFs on cell organelles.
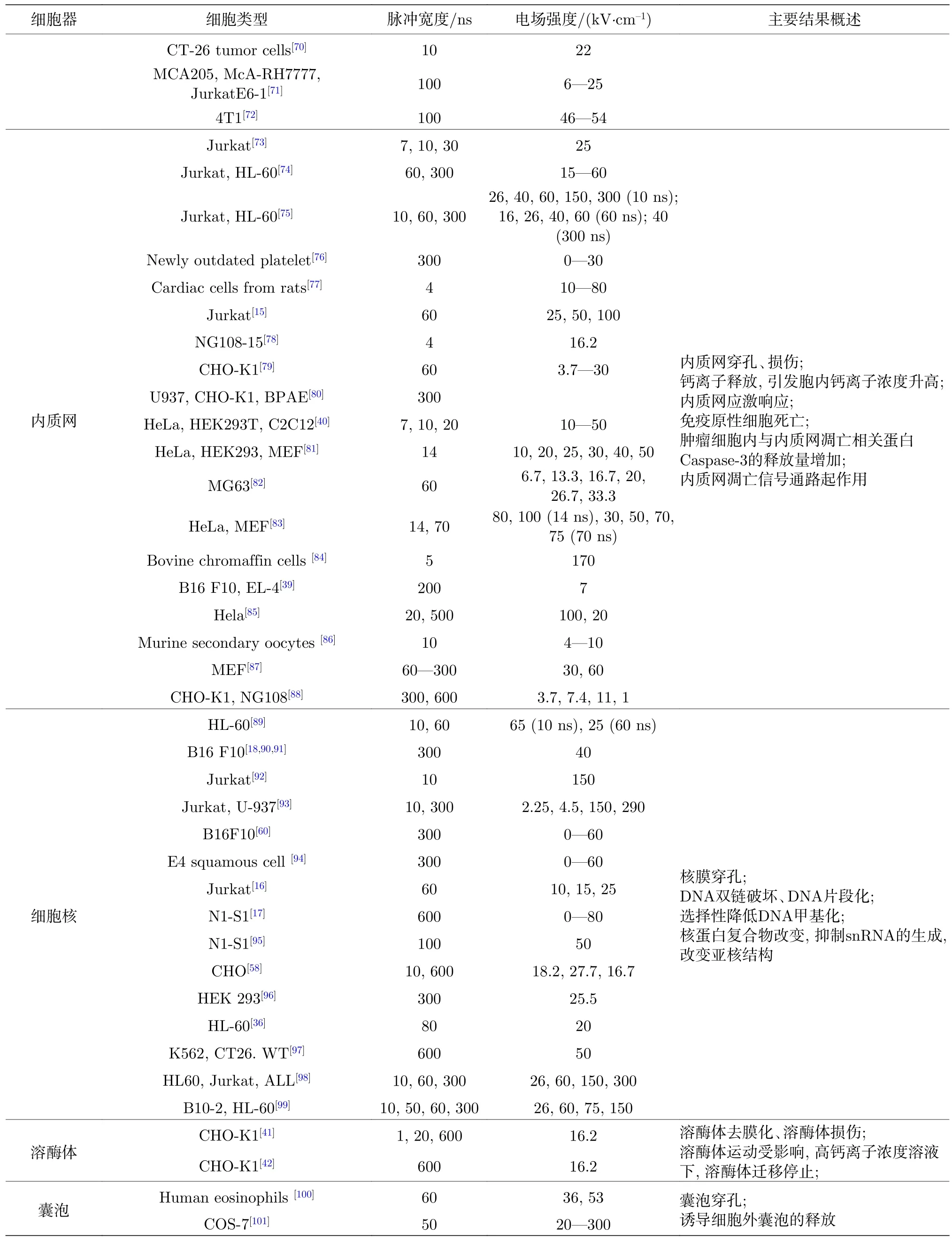
表1 (续)nsPEFs 的细胞器生物电效应总结Table 1 (continued).Summary of effects of nsPEFs on cell organelles.
3 细胞骨架相关变化
3.1 NsPEFs 影响细胞骨架完整性
细胞骨架是一种由蛋白纤维组成的网状结构,主要由肌动蛋白(actin,AT)、微管(microtubules,MT)、中间丝(intermediate filaments,IFs)等组成[102].细胞骨架作为细胞形貌和力学性能的支撑结构,与多种细胞结构和功能相关,在细胞贴壁、增殖分化、信号传导、迁移和细胞存活等方面发挥重要作用[103,104].
研究表明,nsPEFs 对AT,MT 和IFs 都会造成影响[14,105].Berghöfer 等[48]对烟草细胞BY-2 施加10 ns,约33 kV/cm 电脉冲后,观察到肌动蛋白丝向细胞核收缩从而导致细胞核形态发生变化.Steuer 等[61]对大鼠肝上皮细胞(WB-F344)施加20 个100 ns,20 kV/cm 的脉冲,发现肌动蛋白纤维的完整性和有序性在脉冲电场刺激完5—30 min内遭到破坏,随后逐步恢复.该项研究同时观察到细胞间连接直接通信功能被抑制,但由于使用的荧光标记方法耗时较长,并且对低剂量刺激不敏感,难以判断肌动蛋白变化的明确机理.Chafai 等[56,57,106]在2019 和2020 年的研究中指出,使用超分辨平台观察到,nsPEFs 会改变MT 蛋白的瞬时构象,通过控制nsPEFs 参数,能够控制MT 蛋白自组装的进行.这种控制策略不仅对功能和结构生物学过程至关重要,也为合成纳米材料提供了新的思路.
NsPEFs 导致的细胞骨架变化的机理目前尚未得到充分理解.有研究显示在nsPEFs 作用后,细胞形态会发生变化,表现为细胞肿胀、变圆、起泡、起皱、出现足小体等,这些可能是导致细胞骨架破坏的原因[107,108].Pakhomov 等[50]对比了4 个600 ns,19.2 kV/cm 的脉冲作用于不同电击液环境下的中国仓鼠卵巢细胞CHO-K1,结果表明细胞骨架的变化与EP 导致的细胞肿胀有关.该研究使用等渗透压蔗糖溶液抑制细胞在EP 后的肿胀,对比正常电击液下存在细胞肿胀的情况,发现前者AT 解体受到抑制,表明细胞膜EP 导致的细胞膜内外渗透压变化及细胞肿胀是细胞骨架完整性受到破坏的重要原因.这项研究说明nsPEFs 导致的细胞次级效应在nsPEFs 与细胞器相互作用中扮演了重要角色.
NsPEFs 对细胞骨架破坏的机制包括:直接破坏机制和由次级效应引起的细胞骨架破坏间接机制.直接机制包括nsPEFs 引起AT 和MT 等的构象变化,使细胞弹性降低,细胞膜通透性增加.对于参与细胞内运输的MT 的直接破坏,会影响细胞内细胞器位置.如溶酶体的运动受抑制.细胞骨架破坏的间接机制存在多种情况:细胞骨架参与维持细胞形态,nsPEFs 作用后,细胞肿胀,体积增大,诱导细胞骨架破坏;钙离子是调节AT 和MT的信号分子,nsPEFs 作用后的钙离子浓度增加使MT 解聚,破坏细胞骨架;nsPEFs 作用后,细胞内离子浓度变化,为了恢复正常的胞内离子浓度会导致严重的ATP 耗竭,抑制相关蛋白的合成从而使细胞骨架破坏;此外,nsPEFs 引起的脂质信号分子PIP2 的耗竭和磷脂酶C (phospholipase C,PLC)活化也可能是导致细胞骨架破坏的间接机制之一.
3.2 细胞骨架相关亚细胞结构的变化
细胞骨架与细胞膜的连接决定了细胞形状,与细胞间连接、细胞与基底间连接、溶酶体等相关.因此nsPEFs 不仅可以导致细胞骨架解聚,也可能影响与细胞骨架相关的其他亚细胞结构.Thompson等[41]在研究20 个600 ns,16.2 kV/cm 的脉冲对中国仓鼠卵巢细胞(CHO-K1)的影响中,观察到nsPEFs 作用完数秒钟后细胞膜起泡,发生MT 解聚、钙离子流入、溶酶体裂解等一系列细胞内变化.
细胞骨架的组成部分AT 和IFs 等可以跨膜建立起细胞-细胞、细胞-细胞外基质之间的连接,这种连接是维持细胞及组织机械稳定性和实现相邻细胞间的信号传递和分子运输等生化功能的重要结构[109].
紧密连接(tight junction,TJs)可以封闭细胞间缝隙,进而调控分子从顶端向基底面移动,维持组织内不同功能区域,紧密连接还与细胞骨架紧密相连.Shi 等[110-112]利用电阻抗谱结合免疫荧光染色等实验方法观察到不同个数和场强的100 ns脉冲电场对老鼠肝上皮细胞WB-F344 的紧密连接会产生影响,并且观察到这种影响存在脉冲剂量关系.图4 给出了紧密生长的单层细胞在nsPEFs 作用下通透性增加的3 个主要通道,即细胞膜穿孔、细胞间连接的破坏及细胞死亡后脱离导致的空隙.

图4 NsPEFs 对贴壁细胞的影响.1,破坏细胞与细胞间连接;2,细胞膜电穿孔;3,细胞脱落Fig.4.Effect of nsPEFs on adherent cells.1,Disruption of the intercallular connections;2,electroporation;3,cell necrosis or shedding.
间隙连接(gap junctions,GJs)是细胞间膜上六簇连接蛋白组成的供亲水性小分子直接通过的通道,存在于大多数正常细胞中,负责细胞间直接通信,与细胞的增殖、分化和凋亡等相关[113].Steuer等对大鼠肝上皮细胞施加20 个20 kV/cm,100 ns脉冲电场,发现nsPEFs 除了会导致细胞骨架解聚,还观察到细胞膜上的连接蛋白堆积,减少了与膜结合的连接蛋白,从而干扰了间隙连接细胞间直接通信[52].此外,对单层细胞施加不同场强(10,15,20 kV/cm)、不同脉冲数的脉冲电场,结果表明nsPEFs 对GJs 的影响是一个剂量和场强相关的过程.如表1 所列,脉宽从10 ns 到600 ns,场强在10 kV/cm 量级的脉冲电场都能对细胞骨架及相关亚细胞结构产生影响.
细胞骨架及其相关联的细胞连接也是导致nsPEFs 对悬浮细胞和贴壁细胞不同影响程度的主要因素.Stacey 等[98]对4 种不同细胞进行了不同参数的nsPEFs 处理,发现悬浮液中细胞活力大幅下降而贴壁细胞存活率下降相对较小.
4 内质网和线粒体相关变化
4.1 NsPEFs 引起的内质网膜电穿孔及相关效应
钙离子是细胞内的第二信使,对于机体的各项生理活动不可缺少,它可以维持细胞膜两侧的电位差,参与细胞内的信号传导,对细胞内蛋白质调控、细胞凋亡等都起到重要的作用.钙离子广泛分布于内质网和线粒体内,尤其内质网被称为细胞内的钙库[114,115].胞内钙离子增加的途径主要是以下两个:一是胞内释放,即内质网等储存钙离子的细胞器释放钙离子到细胞质中,因此内质网膜穿孔可以显著提高胞内钙离子水平;二是胞外流入,即细胞外微环境中的钙离子通过细胞膜上的离子通道或者是EP 后形成的“纳米孔”进入细胞内[99].
Tolstykh 等[116]研究发现nsPEFs 能够导致内质网膜穿孔,导致钙离子释放到细胞质中,从而增加细胞内钙离子的浓度.Beebe 等[75]探索了10—300 ns 范围的脉冲电场对人类白血病细胞(HL-60)的结构和功能的影响.他们发现在不引起细胞膜EP 的PEFs 作用下,细胞内钙离子水平提高,表明nsPEFs 可以直接作用于内质网使其释放钙离子.
内质网在细胞内负责合成或折叠蛋白质,这项功能受外界刺激而发生变化的现象叫做内质网应激.内质网应激过强或者持续时间过久会导致细胞凋亡,与糖尿病、肥胖症等有关[117].而应激反应是生物体避免外界刺激出现的防卫措施.德岛大学Oyadomari 课题组[40,81,83]发现nsPEFs 会激活内质网应激反应,可能与内质网膜穿孔相关.Furumoto 等[83]对Hela 细胞和鼠类胚胎成纤维细胞(MEF)施加14 ns,80,100 kV/cm;70 ns,20,30,50 kV/cm 两种脉宽的脉冲电场,发现内质网膜穿孔及胞内钙离子释放.另外,nsPEFs 直接诱导真核起始因子(eukaryotic initiation factor-2α,eIF2α)磷酸化,显示nsPEFs 激活了内质网应激反应,并且70 ns 脉冲引起的应激反应强度较14 ns 强.这项研究表明携带有更高频率信息的短脉冲对内质网作用更加明显.
NsPEFs 导致的内质网应激可进一步诱导肿瘤细胞凋亡[118].Rossi 等[39]利用200 ns,7 kV/cm的脉冲电场处理小鼠黑色素瘤细胞(B16-F10)和淋巴瘤细胞(EL-4),发现nsPEFs 引起内质网应激,并伴有免疫原性细胞死亡.这些研究表明脉宽、电场强度是控制nsPEFs 引起应激现象及连锁反应的重要因素.
4.2 nsPEFs 引起的线粒体膜穿孔及相关效应
图5 给出了nsPEFs 诱导细胞凋亡的一种主要途径.线粒体包含触发细胞凋亡的关键调节因子,是诱导肿瘤细胞凋亡的主要靶标[68].细胞凋亡是指细胞为维持内环境稳定,由基因控制的细胞自主有序的死亡,是细胞的主动死亡过程.在凋亡信号触发后,一些凋亡因子,如细胞色素C 从线粒体释放进入细胞质中,然后激活细胞凋亡蛋白酶caspase,诱导细胞凋亡.线粒体凋亡途径中的很多关键事件,包括线粒体信号传导途径的启动和完成,都是由细胞中钙离子信号触发的.因此钙离子不仅可以影响线粒体膜的通透性,还可以通过触发细胞色素C 的进一步释放来放大凋亡信号[98,119].
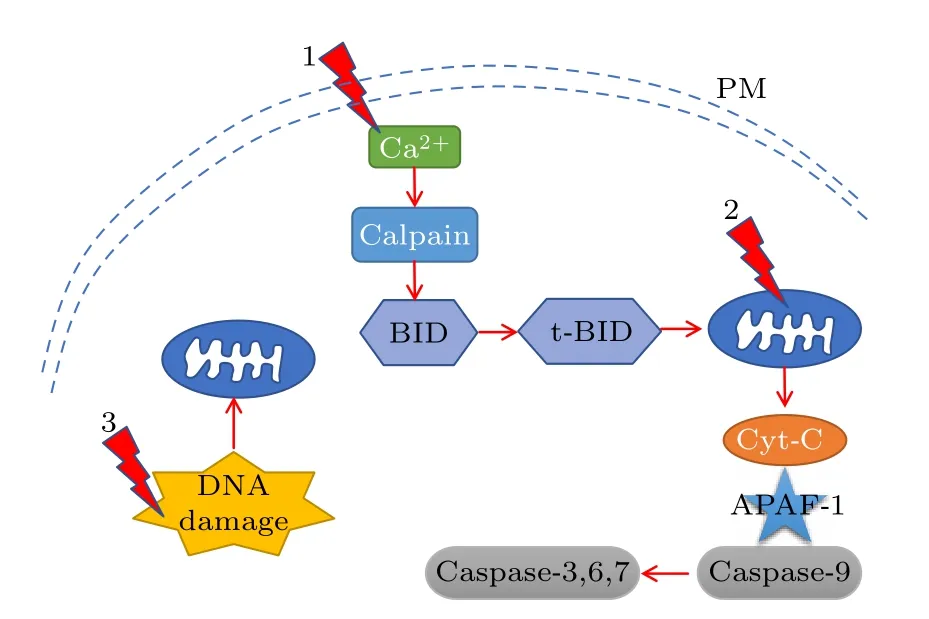
图5 NsPEFs 诱导的细胞内凋亡途径Fig.5.Apoptosis pathways for cells after exposure to nsPEFs.
研究发现,nsPEFs 可引起线粒体膜电位耗散、线粒体肿胀等变化[70,120,121].线粒体膜电位的耗散与细胞活力丧失之间存在密切的关系,并且与细胞质中钙离子相关.Guerrero-Hernandez 等[36]对肝癌细胞施加场强最高达80 kV/cm,脉宽600 ns,上升下降沿时间分别为15 和150 ns 的多种强度脉冲电场.发现nsPEFs 作用后的肝癌细胞产生明显的线粒体膜电位耗散.其中nsPEFs 的上下沿,即高频成分,发挥了主要作用.nsPEFs 使线粒体电穿孔同内质网一样也会导致胞内钙离子释放,并且往往伴随着caspase 蛋白酶激活,细胞活力丧失.熊本大学的Morotomi-Yano 等[65]对HeLa S3 细胞施加80 ns,20 kV/cm,1 Hz 的电脉冲,结果显示nsPEFs 诱导释放的钙离子参与了细胞内信号传导,并且这些信号传导事件与细胞死亡和存活的调节相关.Muratori 等[96]对HEK 293 细胞施加了300 ns,25.5 kV/cm 的脉冲电场,发现nsPEFs激活了TMEM16F 蛋白酶(或Anoctamin 6),其中TMEM16F 蛋白酶与钙离子和细胞凋亡有关.如表1 所列,通过改变nsPEFs 参数,可以针对不同细胞调控其胞内钙离子浓度从而调节细胞功能,如细胞凋亡[122,123].Nuccitelli 等[71]和Guo 等[72]报道免疫原性细胞死亡的特征是释放危险相关分子模式(danger-associated molecular patterns,DAMPs),ATP 是其中一种关键性DAMPs.ATP 耗竭会迅速导致细胞坏死,研究表明nsPEFs 处理后,细胞内ATP 被消耗.nsPEFs 还会引起细胞内的ROS增加,众多研究发现,nsPEFs 引起胞内ROS 水平显著增加,产生的ROS 可能导致细胞刺激和氧化性细胞损伤,并通过凋亡、坏死或焦亡等信号通路导致细胞死亡[60,94,124,125].
5 细胞核相关变化
细胞核是细胞遗传与代谢的调控中心,nsPEFs引起的细胞核变化是多样的[126].Zhuang 等[127]利用单细胞介电壳层模型对比100 ns 和100 µs 脉冲电场对Jurkat 细胞膜、细胞质、细胞核等亚细胞结构的介电变化,发现nsPEFs 除了与µsPEFs 一样导致细胞膜EP,还会导致细胞核膜电导率增加,表明nsPEFs 可以导致细胞核膜电穿孔.Chen 等[89]采用实时荧光标记成像方法研究10 ns,65 kV/cm和60 ns,25 kV/cm 两种脉宽、能量相同的脉冲电场对HL-60 细胞核的影响,结果表明nsPEFs 可以导致细胞核电穿孔并影响核质.Chen 等[92]在2007年报道了60 ns,60 kV/cm 的脉冲电场会损伤Jurkat 细胞和HL-60 细胞的DNA.他们发现对核膜进行充电会导致核膜中形成纳米孔,并且nsPEFs会通过抑制snRNA 的生成改变亚核结构.此研究首次表明nsPEFs 改变了核蛋白复合物.Chen 等[18]对黑色素瘤细胞(B16-F10)施加100 个300 ns,40 kV/cm 的电脉冲后,发现nsPEFs 诱导DNA 双链断裂,降低了血管内皮生长因子(VEGF)和血小板衍生的内皮细胞生长因子(PD-ECGF)蛋白的表达,促进了细胞凋亡.
nsPEFs 引起的细胞核变化包括:1) nsPEFs可以导致核膜的电穿孔[97];2) nsPEFs 可以影响核RNA-蛋白质复合物及核斑点的变化[92],主要表现为核内亚结构数量和大小的变化.核RNA-蛋白质复合物的变化反映了细胞转录机制的破坏.此外,由于DNA 是带电物质因此易受电场的影响,nsPEFs 可以直接诱导DNA 损伤从而使细胞死亡.另一方面,核被膜的穿孔也可以促进DNA 质粒进入核内,增强细胞的基因表达[75].
6 其他亚细胞结构
除了上述提到的细胞器生物电效应,nsPEFs影响溶酶体的运动,使溶酶体去膜化、损伤溶酶体等[128].溶酶体可以降解损伤的细胞膜和细胞器,参与细胞自噬过程.2014 年,Thompson 等[41]发现了溶酶体、钙离子和细胞骨架三者之间存在联系,研究发现在含钙离子的培养基中生长的细胞在施加高阈值剂量的nsPEFs 后,MT 破坏,溶酶体迁移停止.2018 年,Thompson 等[42]为了探究细胞内钙离子浓度是如何影响溶酶体运动,对中国仓鼠卵巢细胞(CHO-K1)施加了600 ns,16.2 kV/cm的纳秒脉冲电场,研究发现在含钙离子的溶液中,施加nsPEFs 会破坏细胞骨架结构,从而阻止溶酶体运动.他们分别拍摄了高钙离子浓度的溶液和不含钙离子的溶液中溶酶体运动图像,发现溶酶体运动情况也受细胞内钙离子浓度影响.高钙离子浓度的溶液中,细胞内溶酶体迁移停止,而在缺乏钙离子溶液中,一些溶酶体发生移位.Kielbik 等[129]研究发现钙离子与细胞骨架破坏之间存在相关性,通过共聚焦显微镜观察,钙离子浓度增加所引起细胞骨架破坏程度更加显著,这与Thompson 等的研究一致.由于细胞骨架受到破坏,因此受细胞骨架牵引的溶酶体运动受到抑制.
NsPEFs 作用下,囊泡的脂质双分子层形成瞬时孔隙,囊泡膜的通透性增加[130].Qian 等[131]在研究nsPEFs 介导的抗肿瘤免疫反应机制时发现,nsPEFs 诱导了细胞外囊泡的释放.
细胞各亚结构相互联系紧密,因此nsPEFs 与细胞的相互作用往往会存在多种细胞器的物理结构和生化功能的变化,及其他次级效应.除了上述主要的细胞器结构及相关的细胞间连接、溶酶体、囊泡等,有文章报道nsPEFs 对细胞内的酶也会产生影响[132].Della Valle 等[133]通过分子动力学仿真100 ns 脉冲电场对超氧化物歧化酶结构的影响,表明103kV/cm 以上的nsPEFs 可以直接影响酶活性部位的静电场,从而导致酶结构发生变化.Guo 等[134]测量了电场5—30 kV/cm,100 和300 ns脉冲电场作用于小鼠口腔癌后,口腔癌细胞内NOS的变化,预示一氧化氮合成酶可能受到影响.
7 结论
本文基于nsPEFs 的细胞器生物电效应的研究发展简史及不同细胞器结构和功能变化等展开叙述,总结了2000 年以来nsPEFs 的细胞器生物电效应相关文献,尤其是近五年的研究.通过表1所列的总结数据可以看出,对于施加同一种脉冲参数,不同细胞类型在nsPEFs 处理后的细胞器生物电效应存在显著差异.对于同一种细胞,仅仅改变nsPEFs 参数的其中一项指标,比如脉宽、电场强度、脉冲个数、脉冲极性等,细胞器效应的表现也会产生明显变化.因此在研究nsPEFs 的细胞器效应时,需要结合特定的脉冲参数及细胞类型进行讨论.根据现有的研究综述,作者提出若干未来研究中值得加强关切的问题:
1) 已有的研究多聚焦于nsPEFs 对悬浮细胞的细胞器生物电效应,悬浮状态下的细胞以单一个体的形式存在,而实际应用往往在组织层面进行,未来的研究应更多考虑在多细胞体系或生物组织中发挥重要作用的细胞间连接,细胞与外基质间连接的影响.
2) 传统的基于荧光等标记物的观察手段,以及扫描电子显微镜等都会对被测物产生不可逆伤害,而且往往难以观察到脉冲电场生物电效应的快速响应.一些物理技术,比如生物介电谱方法,具有非侵入性、实时、原位观测的优势,因此可以用生物介电谱技术研究nsPEFs 的细胞器生物电效应.
3) 超分辨光学显微成像技术除了具有非侵入性、实时原位观察的优势,还具有较高的空间分辨率,可达几十纳米.超分辨成像可以为进一步深入理解nsPEFs 的细胞器生物电效应及相应机理提供更多新信息,促进nsPEFs 技术在生物医学领域的进一步发展与应用.
4) 大量研究表明nsPEFs 可以抑制肿瘤生长和诱导抗肿瘤免疫反应,nsPEFs 诱导癌细胞凋亡涉及的靶点较多,且基本都与细胞器相关.因此,加强nsPEFs 的细胞器生物电效应的研究有助于发现癌细胞凋亡靶点,开发新的肿瘤免疫疗法.
5) 近年的研究和临床试验表明,µsPEFs 可作为一种新型物理能量用于心肌组织消融,治疗房颤等心脏疾病.相比µsPEFs,nsPEFs 引起的肌肉收缩更弱、气泡更少、消融更均匀,有望成为更好的脉冲电场心肌消融技术.此外,房颤发生与心肌细胞间连接等细胞器紧密相关,未来可加强nsPEFs对心肌细胞及其细胞器的相关研究.
现有诊断技术的局限和人们对细胞和组织等生物体系本身理解的不足,使得nsPEFs 的生物电效应尚未得到充分理解,本文聚焦nsPEFs 的细胞器生物电效应,旨在为nsPEFs 肿瘤治疗、软组织消融等生物医学领域的研究和应用提供参考.
