基于RNA测序分析不同低氧暴露模式对大鼠胫骨前肌差异基因表达的影响
2022-03-24龚丽景贾杰付鹏宇
龚丽景贾杰付鹏宇
(1.北京体育大学 运动与体质健康教育部重点实验室,北京 100084;2.北京体育大学 运动人体科学学院,北京 100084;3.西北工业大学体育部,西安 710072)
骨骼肌是人体内最大的器官,是蛋白质储存和糖代谢的重要场所,在维持姿势、运动、呼吸、能量和代谢稳态、体温维持和调节及内分泌等方面都发挥着重要的作用[1]。同时,骨骼肌也是高耗能组织,氧气不足会明显影响骨骼肌代谢,促进肌肉蛋白质代谢率增加,诱导肌萎缩,常见于缺氧性疾病如慢性阻塞性肺疾病(chronic obstructive pulmonary disease,COPD)和阻塞性睡眠呼吸暂停低通气综合征(obstructive sleep apnea-hypopnea syndrome,OSAHS)等,或见于世居平原者进入高原环境旅游、训练或从事军事活动。骨骼肌萎缩会导致肌肉质量减少、功能下降,从而威胁缺氧性疾病患者健康,减弱运动员和士兵高原训练的效果,或影响高原旅游大众的生产生活[2]。
骨骼肌在缺氧时会经历3个时期:预警期、抵抗/适应期和失适应期[3]。预警期中骨骼肌突然受到低氧刺激,会出现一系列应激反应,导致肌肉蛋白分解增加和一系列肌肉功能相关蛋白表达的改变;而在适应期内骨骼肌会发生解剖和生理上的过度补偿,随着低氧暴露时间的延长,又会进入失适应阶段,肌萎缩程度进一步加重。提示不同模式的低氧暴露诱导肌萎缩可能经历不同的生理过程。数日的急性低氧暴露可模拟初上高原的情况。研究发现,8%氧浓度环境下暴露2 d和4 d均可导致小鼠骨骼肌蛋白质分解增加,诱导骨骼肌萎缩[4]。慢性间歇低氧暴露可模拟生理性缺氧疾病的病理过程或运动员的低住高练(living low-training high,LoHi)的训练环境。研究显示,慢性间歇低氧暴露可降低大鼠的骨骼肌指数(骨骼四肢骨骼肌质量之和/身高2),促进大鼠骨骼肌组织和成肌细胞的衰老[5]。上述研究证实急性低氧和慢性间歇低氧可诱导骨骼肌萎缩。
不同类型的肌纤维在形态、代谢和收缩特性等方面存在较大差异[6]。低氧暴露所致的肌萎缩具有肌纤维选择性。本实验室前期研究对比了慢肌、快肌和混合肌对4周连续低氧(氧浓度12.4%)的适应情况,发现快肌对低氧更为敏感[7],因此本研究选择了快肌——胫骨前肌作为研究对象。有关低氧诱导肌萎缩的分子作用机制尚存在争议,多数研究以骨骼肌蛋白的合成和分解相关蛋白的表达入手,难以深入探究该过程的机制,且不同低氧暴露模式可能诱导不同的基因信号以参与肌萎缩的发生,因此本研究拟通过采用RNA测序的研究方法,探究急性低氧暴露和慢性间歇低氧暴露对大鼠胫骨前肌的差异基因表达影响及其富集通路,为治疗缺氧性疾病和防治高原肌萎缩提供可能的分子靶点。
1 材料与方法
1.1 材料
1.1.1 实验动物
24只10周龄SPF级雄性SD大鼠,初始体重为(333.56±6.35)g,购于北京维通利华实验动物技术有限公司【SCXK(京)2015-0001】,随机分为常氧对照组(C组,n=16)和慢性间歇低氧组(IH组,n=8),IH组大鼠每天在氧浓度为12.4%的低氧房内暴露8 h,共4周。在干预的第25天(干预结束前3 d),将C组大鼠按体重随机均分为2组,一组继续维持C组常氧环境饲喂,另一组命名为急性低氧组(AH组,n=8),AH组大鼠置于低氧房,干预方案为每天24 h,氧浓度为12.4%,共干预3 d,分组情况如图1。动物饲养在北京体育大学动物实验室【SYXK(京)2016-0033】内进行。本项目获得北京体育大学运动科学伦理审查委员会的批准(批准号:2019038A)。每天记录大鼠体重和摄食量。
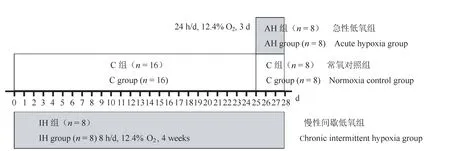
图1 大鼠分组及干预示意图Figure 1 Diagram of rats grouping and intervention
1.1.2 主要试剂与仪器
戊巴比妥钠(Merck,P8410),Anti-Atrogin1/Fbx32(Abcam,ab74023),Anti-MuRF1(Abcam,ab172479),Anti-α-tubulin(Sigma,T6074),山羊抗兔二抗(LI-COR,926-68071),RNA保存液等为国产试剂。
小动物抓力仪(DB-YLS-13 A,北京智鼠多宝),双能 X射线骨密度仪(dual energy X-ray absorptiometer,DEXA;Lunar Idxa,美国),电子天平(Sartorius,美国),倒置光学显微镜(Leica,DMI3000,德国),电泳槽和干转转膜系统(Life Technologies,美国),近红外光谱(Near Infrared)检测系统(LI-COR,美国)。
1.2 方法
1.2.1 抓力仪测试抓力
每周训练大鼠练习1次抓力仪,干预结束即刻测试抓力,测量3次,取最大值,并计算相对抓力(最大抓力/体重)。
1.2.2 DEXA法测量瘦体重
低氧干预结束后,所有大鼠禁食12 h,腹腔注射2%戊巴比妥钠(40 mg/kg体重)麻醉,DEXA测量体成分,计算瘦体重百分比(瘦体重/体重)。
1.2.3 取材
干预4周后,麻醉取材,心脏取血处死,取胫骨前肌(musculi tibialis anterior,TA),分成3份,1份置于RNA保存液中,用于提取RNA和测序;1份置于液氮冷冻后转移至-80℃保存,用于蛋白检测;第3份置于4%多聚甲醛中固定,后续石蜡包埋、切片和HE染色。
1.2.4 胫骨前肌肌纤维形态和面积计算
TA组织HE染色后,在10×40倍镜下观察并拍照。肌纤维横截面积(fiber cross sectional area,FCSA)用软件Image-Pro Plus(IPP)6.0计算。
1.2.5 Western Blot法检测蛋白质表达
TA总蛋白质提取后,用聚丙稀酰胺梯度胶电泳分离蛋白,转膜至NC膜上,明胶溶液封闭,孵育Atrogin1和MuRF1抗体,4℃过夜,室温孵育山羊抗兔IgG抗体,最后使用Image Studio Ver 5.2软件曝光,对条带的信号值进行相对定量分析,以α-tubulin为内参。
1.2.6 骨骼肌RNA测序和功能分析
TA的RNA提取后进行片段化处理,然后进行cDNA的合成,PCR富集文库片段,基于华大基因BGISEQ平台进行RNA测序。对测序数据进行过滤、质量评估和比对结果评估,然后进行表达量计算,分析差异表达基因,并使用基因本体论数据库(Gene Ontology,GO)分析、基因和基因组百科数据库(Kyoto Encyclopedia of Genes and Genomes,KEGG)对差异基因进行富集分析和Pathway分析。
1.3 统计学分析
数据以平均值±标准差(±s)表示,采用SPSS 22.0软件进行分析,各组间比较采用单因素方差分析,显著性差异以P<0.05或P<0.01表示;采用DESeq对基因表达进行差异分析,筛选差异倍数Fold Change>2,显著性Pvalue<0.05被认为富集分析具有统计学意义。
2 结果
2.1 干预期间大鼠的体重、摄食量和瘦体重
干预期间,IH组体重在各时间点均低于C组,但无显著性差异;AH组干预3 d期间呈下降趋势,且随着干预时间的延长,AH组较C组的体重差值逐渐增加(各时间点下降率分别为2.99%、3.76%和5.88%),但各时间点较C组无显著差异,如图2A。IH组摄食量在干预前3 d低于C组,但在干预后期逐渐与C组接近;AH组摄食量干预期间持续低于C组,如图2B。干预后测试各组大鼠瘦体重,AH组大鼠瘦体重显著低于C组(P<0.05),IH较C组也有下降趋势,但无显著性差异,如图2C;各组间瘦体重百分比无显著性差异,如图2D。

图2 各组大鼠体重、摄食量变化和瘦体重Figure 2 Changes of body weight,food intake and body mass in the rats
2.2 干预后大鼠抓力
干预末次测试大鼠抓力,与C组相比,IH组和AH组大鼠抓力均有下降,且AH组下降更为明显,但各组间无显著性差异,如图3A;用抓力/体重表示相对抓力,比较各组大鼠的相对抓力结果显示,AH组抓力较C组显著下降(P<0.01),如图3B。

图3 干预后各组大鼠抓力和相对抓力Figure 3 Grasping force and relative grasping force of the rats after intervention
2.3 干预后大鼠胫骨前肌FCSA
HE染色观察大鼠TA肌纤维形态发现,AH组和IH组较C组肌纤维形态呈现不规则状,肌纤维面积大小不一,肌间隔增加,肌纤维出现裂痕,IH组较AH组更加明显,如图4A;统计FCSA结果显示,IH组和AH组FCSA均低于C组,但均无显著性差异,如图4B。

图4 干预后各组大鼠肌纤维形态和FCSAFigure 4 Muscle fiber morphology and FCSA of TA in the rats after intervention
2.4 干预后大鼠肌萎缩蛋白Atrogin1和MuRF1含量
干预后,IH组大鼠TA中肌萎缩相关蛋白肌萎缩 F-box蛋白(muscle-specific F-box protein,Atrogin1)和肌肉特异性环指蛋白1(muscle-specific ring finger 1,MuRF1)[8]相对含量显著高于C组(P<0.05),AH组大鼠肌萎缩蛋白Atrogin1相对含量显著高于C组(P<0.05),如图5。

图5 干预后各组大鼠TA中Atrogin1和MuRF1蛋白条带和相对含量Figure 5 Atrogin1 and MuRF1 protein bands and relative content of TA in the rats after intervention
2.5 干预后大鼠TA测序结果
2.5.1 IH/C组和AH/C组的差异基因筛选
IH/C组差异基因共有228个,其中上调基因128个,下调基因100个;AH/C组差异基因共有380个,其中上调基因289个,下调基因91个。
在IH/C组和AH/C组中均上调的基因有59个;均下调的基因有29个;在AH/C组中上调而在IH/C组中下调的基因有6个,分别为:慢肌肌钙蛋白I1(troponin I type 1,slow skeletal,TNNI1),肌球蛋白轻链2(myosin light chain 2,MYL2),慢肌肌钙蛋白C1(troponin C type 1,slow,TNNC1),慢肌肌钙蛋白T1(troponin T type 1,slow skeletal,TNNT1),锌指蛋白385B(zinc finger protein 385B,ZFP385B)和LOC501116;在AH/C组中下调而在IH/C组中上调的基因有1个,为磷脂酶A2 IIA(phospholipase A2,group IIA,PLA2G2A),如图6。

图6 IH/C组和AH/C组上/下调差异基因UpSet图Figure 6 UpSet diagram of differential genes in groups IH/Cand AH/C
2.5.2 GO分析差异基因所富集的生物过程
根据Pvalue值大小排序,列出各组排名前20的上/下调差异基因所富集的生物过程(biological process,BP)(从上至下Pvalue值依次增加)。
IH/C组上调差异基因所富集的BP主要为:谷胱甘肽代谢,细胞死亡的负性调节,低氧应答,胰岛素应答,葡萄糖稳态状态,对糖皮质激素的反应,对氧化还原状态的反应和葡萄糖代谢过程;IH/C组下调差异基因BP主要为:快慢肌之间的转化,调节骨骼肌收缩,对过度缺氧的反应,对低氧的反应和对过氧化氢的反应。根据Pvalue值大小排序,列出各组差异基因所富集的通路(从上至下Pvalue值依次增加)。IH/C组上调差异基因所富集的通路主要为过氧化物酶体增殖物激活受体(peroxisome proliferator activated receptor,PPAR)信号通路和谷胱甘肽代谢通路(glutathione metabolism),如图7A。其中,PPAR信号通路所富集的差异基因包括:脂联素(adiponectin,ADIPOQ),血管生成素样蛋白4(angiopoietin-like factor 4,ANGPTL4),外周脂蛋白1(perilipin 1,PLIN1),磷酸烯醇式丙酮酸羧激酶1(phosphoenolpyruvate carboxykinase 1,PCK1)。
AH/C组上调差异基因所富集的BP主要为:氧气运输,快肌和慢肌之间的转化,骨骼肌收缩,ATP代谢,低氧应答,神经元动作电位,肌肉收缩,对过氧化氢的反应,炎症应答,骨骼肌纤维发育和细胞死亡的调节;AH/C组下调差异基因BP主要为:磷脂代谢过程,smad蛋白信号转导,肌管分化的正向调节,泛素蛋白连接酶结合,脂质磷酸酶活性,如图7B。

图7 IH/C组和AH/C组差异基因所富集的BP及PathwayFigure 7 Enriched BP and pathway of differential genes in groups IH/Cand AH/C
3 讨论
生理性缺氧疾病和高原/低氧环境造成的骨骼肌萎缩都会给机体健康带来巨大风险。许多研究采用不同的低氧暴露时长和低氧浓度(海拔高度)均观察到了骨骼肌萎缩的现象,其中在海拔3700~4300 m时,瘦体重的减少最为明显[9],因此本研究选择了海拔4000 m所对应的氧浓度作为低氧干预浓度。但有关不同低氧暴露方式导致肌萎缩的作用机制尚无定论。
本研究显示,慢性间歇低氧暴露后,大鼠体重在干预期间持续低于常氧组,4周干预结束后,虽然大鼠骨骼肌已对低氧产生一定的适应(表现为瘦体重、抓力、FCSA无显著改变),但HE染色可见TA肌间隔增加,肌纤维呈现不规则状,且肌萎缩相关蛋白Atrogin1和MuRF1表达均显著增加,提示慢性间歇低氧暴露仍对肌纤维产生了一定程度的损伤[10];急性低氧暴露后,大鼠体重在干预期间持续降低,且降低率随低氧暴露时间的延长而增加,干预3 d后,瘦体重和相对抓力较常氧组显著下降。其中,急进低氧环境造成的食欲抑制及胃肠功能障碍造成的摄食量下降可能是诱因之一;此外,TA肌间隔也有所增加,但肌纤维损伤程度不及间歇低氧干预,且肌萎缩蛋白MuRF1表达较常氧组增加。此研究结果提示,慢性间歇和急性低氧暴露均可导致大鼠骨骼肌出现一定程度的萎缩,但不同的低氧模式对大鼠骨骼肌的影响不同,慢性间歇低氧主要从微观结构和分子层面影响TA,而急性低氧主要从质量和功能层面影响TA,说明不同的低氧模式可能通过不同途径影响骨骼肌。
因此,本研究通过骨骼肌RNA测序探究不同低氧模式对TA差异基因表达的富集通路,发现慢性间歇低氧可上调骨骼肌的抗氧化过程(谷胱甘肽代谢)、糖代谢(胰岛素应答、葡萄糖稳态状态和葡萄糖代谢过程)和PPAR通路。研究发现,葡萄糖利用障碍和脂代谢紊乱可损害骨骼肌质量和功能,而谷胱甘肽(glutaredoxin,Grx)是氧化还原稳态的重要代谢调节因子,骨骼肌中含有丰富的Grx,Grx缺失小鼠可出现肌萎缩,并伴随有骨骼肌葡萄糖利用障碍和脂质沉积[11]。本研究中,IH/C组差异通路——PPAR通路中所富集的脂代谢基因全部为上调基因(ADIPOQ、PLIN1、PCK1),且在IH/C组中上调而在AH/C组下调的差异基因为PLA2G2A,这些基因均与糖脂代谢密切相关[12]。研究发现,间歇低氧可影响骨骼肌中脂质的含量。中重度OSAHS患者循环系统中棕榈酸(palmitic acid,PA)和游离脂肪酸(free fatty acids,FFAs)在过夜后增肌,并可异位至股外侧肌中并导致胰岛素抵抗(insulin resistance,IR),导致肌肉中脂质增加,且增加程度与呼吸暂停低通气指数(apnea-hypopnea index,AHI)呈正相关[13]。提示慢性间歇低氧可通过损害TA的糖脂代谢功能而导致骨骼肌功能受损,而此过程中伴随着以Grx为主的抗氧化能力增强,说明骨骼肌在损伤同时也逐步对低氧环境产生习服,并表现为骨骼肌质量的恢复。
急性低氧可上调骨骼肌的肌纤维类型的转化(快慢肌之间的转化)、氧化应激反应(对过氧化氢的反应)和炎症反应过程。急性低氧主要对肌纤维类型的转化作用主要表现为促进快肌向慢肌的转化,表现为调节慢肌结构的相关基因(TNNI1、TNNC1、TNNT1和ZFP385B[14])表达增加。研究发现,急性低氧干预(氧浓度6.5%,暴露4 h)可降低甘肃鼢鼠骨骼肌中糖酵解相关酶的活性,增加肌红蛋白的含量,说明在急性低氧的应激刺激下,机体可通过向慢肌转变,利用慢肌特征以保持肌肉较充分的氧供应,从而减少或避免低氧对骨骼肌造成的损伤[15]。而急性低氧还可造成骨骼肌炎症和氧化应激水平增加,从而造成肌萎缩的发生。研究发现急性低氧可增加脯氨酰羟化酶3(prolyl hydroxylase 3,PHD3)的表达,进而增加低氧诱导因子-1α(hypoxia inducible factor-1α,HIF-1α)的转录[16]。而PHD3高表达可以促进肌萎缩的发生,其发挥作用的机制与介导炎症信号通路而影响骨骼肌中蛋白质的转运密切相关[17]。此外,急性低氧暴露还可增加骨骼肌的氧化应激水平,该过程可能受到核因子E2相关因子2(nuclear factor erythroid 2-related factor 2,Nrf2)-过氧化氢酶通路的调控[18-20]。提示急性低氧可促进快肌纤维向慢肌纤维转变,以适应低氧应激对肌肉的损伤,但仍不敌该过程中伴随着的炎症反应和氧化应激水平的增加,最终导致肌萎缩的发生,并降低肌肉力量[21]。
4 结论
不同低氧模式对大鼠胫骨前肌基因表达产生不同的影响。慢性间歇低氧可能通过影响骨骼肌的糖脂代谢基因水平以促进肌肉的损伤,但该过程伴随着抗氧化能力的提高,可能由此促进肌肉对低氧的适应;急性低氧虽可促进胫骨前肌肌纤维向慢肌的转化,但不敌急性低氧激活氧化应激和炎症反应,这可能是肌萎缩的诱因。
