养殖水体1株好氧反硝化菌的分离鉴定及脱氮特征研究
2022-03-22张艳昕章晓栋沈文英
曲 寅 张 涵 张艳昕 章晓栋 沈文英
(绍兴文理学院 生命科学学院,浙江 绍兴 312000)
随着规模化高密度水产养殖的发展,养殖水体中的有机物、氨氮、亚硝酸盐氮等污染物的浓度不断累积升高,导致水质恶化,养殖动物病害频繁发生,养殖水体生态遭到严重破坏,水产品品质下降[1].亚硝酸盐含量升高,说明池塘底部缺氧,养殖水质恶化,同时也是水生动物发病的前期征兆[2].亚硝酸盐是三态氮(氨氮-亚硝酸盐-硝酸盐)的中间形态,受微生物的作用而活化,是一种不稳定的形式,在溶解氧充足时可转化为无毒的硝酸盐,而在缺氧的条件下转化为毒性更强的氨[3].与物理去氮、化学去氮相比,生物去氮被认为是一种经济、有效和最有发展前途的方法,具有工艺简单、成本低廉、较易推广等特点[4].
中华鳖(Trionyxsinensis),俗称甲鱼,是我国重点水产养殖动物之一[5],主要采用高密度池塘养殖模式.在中华鳖养殖后期,养殖水体中的氨氮、亚硝酸盐氮等污染物的浓度增加[6-7],水产养殖动物疾病的发生率也增加[1].本试验从高密度中华鳖养殖水体中筛选出降亚硝酸盐氮的好氧反硝化菌种[8],分析了其降亚硝酸盐氮效果及其最适培养条件,为进一步研制降亚硝酸盐氮微生物制剂提供试验基础.
1 材料与方法
1.1 实验材料
本实验所用中华鳖高密度养殖池水样取自绍兴大畈水产合作社(浙江,绍兴).
1.2 实验方法
1.2.1 好氧反硝化菌的筛选
菌落的分离与培养:水样用PBS缓冲液进行梯度稀释至原液的1、10、100和1 000倍,取200 μL均匀涂布于营养琼脂平板上,于37 ℃恒温培养24 h后,随机挑取100个生长良好的单菌落接种于液体营养液培养基中,于180 rpm,37 ℃培养24 h.按照1%的接种量,将菌种接种到以亚硝酸钠为氮源的液体培养基中,于180 r/min,37 ℃培养24 h,通过定性显色反应检测不同菌株利用亚硝酸盐氮的情况.
定性显色反应:培养液用亚硝酸盐显色剂点样,若显示红色说明培养液中存在亚硝酸盐氮,颜色越深浓度越大.若不显示红色说明培养基中不存在亚硝酸盐氮,定性地鉴定该菌种可以消除培养基中亚硝酸盐氮.显色初筛之后通过N-(1-萘基)-乙二胺光度法测定亚硝酸盐含量,计算亚硝酸盐氮的消除率.
1.2.2 好氧反硝化菌的鉴定
采用16S rDNA序列鉴定方法鉴定菌种.用试剂盒法提取筛选获得的菌株DNA,采用16S rDNA通用引物进行PCR扩增,PCR反应体系(50 μL)为:10×Buffer5.0 μL,上下游引物各1.0 mL,dNTP4.0 μL,Taq酶0.5 μL,模板1.0 μL,dd H2O37.5 μL;反应条件为94 ℃预变性5 min,94 ℃,变性50 s,52 ℃退火60 S,72 ℃延伸1 min30 s,共30个循环.对阳性克隆的片段进行测序(上海,生工生物有限公司),测序结果与GenBank中的16S rDNA序列进行比对分析.
1.2.3 亚硝酸还原酶鉴定
以菌株DNA为模板,根据Genbank上亚硝酸还原酶基因nirS、nirK全长序列设计引物,进行PCR扩增.nirS F:5-ATGAGCAATGT(T/C)GGTAAACCTA-3,nirS R:5-TCAACACCATG(A/C)ACGACGTGTACTAA-3;nirK1 F:5-GGMATGGTKCCSTGGCA-3,nirK5 R:5-GCCTCGATCAGRTTRTGG-3.PCR反应体系(50 μL)为:10×Buffer5.0 μL,正反向引物各2.0 μL,dNTP4.0 μL,Taq酶0.5 μL,模板1.0 μL,ddH2O35.5 μL;PCR反应条件:95 ℃预变性2 min;94 ℃变性40 s,50 ℃退火30 s,72 ℃延伸2 min,反应10个循环;94 ℃变性40 s,55 ℃退火30 s,72 ℃延伸2 min,反应22个循环;重组克隆菌进行序列双向测定(上海,生工生物有限公司),测序结果采用BLAST程序进行同源性检索.
1.2.4 最适培养条件筛选
取-80 ℃保存的菌株活化扩大培养,按1%的接种量接种至不同培养基.以亚硝酸盐为氮源、柠檬酸钠为碳源,设置不同初始亚硝酸盐氮浓度、碳氮比、温度和pH值筛选好氧反硝化菌的最适生长和降亚硝酸盐氮效果的培养条件.初始亚硝酸盐氮浓度分别为10、20、40、60、80、100、120、140 mg/L,C/N分别为2、4、6、8、10、12,培养温度分别为20、25、30、35 ℃,pH分别为5、6、7、8、9.每个实验组设置3个重复.培养24 h,取0 h和24 h菌液各10 mL,测定OD600,计算菌种生长值.其余菌液于8 000 r/min离心5 min,取上清测亚硝酸盐的浓度,计算亚硝酸盐氮消除率.
1.2.5 养殖污水试验
取NO2-N浓度为5 mg/L的水产养殖污水,将菌株活化,按照最适培养条件培养获得浓度为108CFU/mL的菌液,以1∶1 000的比例投入模拟污水中,设置对照组,水体不添加任何菌液,每组设置3个重复.分别于实验第2 d、5 d,用N-(1-萘基)-乙二胺光度法测定水体中亚硝酸盐氮浓度.
1.3 数据处理
实验数据用平均数±标准差表示,采用SPSS统计软件进行单因子多重分析,P<0.05表示存在显著性差异.
2 实验结果
2.1 养殖水体好氧反硝化菌的筛选
显色鉴定结果表明,有8个菌株具有转化亚硝酸盐氮的能力,培养24 h后,培养基中亚硝酸盐氮消除率均达到98%以上,显著高于对照组消除率(65.50%).
2.2 好氧反硝化菌的鉴定
通过16S rDNA序列鉴定分析,具有降解亚硝酸盐氮菌株为肺炎克雷伯菌(Klebsiellapneumoniae).核酸序列相似性均为99%.
2.3 亚硝酸还原酶基因的鉴定
通过对肺炎克雷伯菌(K.pneumoniae)的亚硝酸还原酶基因的nirS、nirK进行扩增,结果表明该菌为nirS+、nirK-菌.对扩增得到的nirS片段部分测序,并与GenBank其他细菌的nirS序列进行比对发现,基因同源性达99%.
2.4 最适生长和降亚硝酸盐氮效果的培养条件筛选
以NaNO2为氮源、以柠檬酸钠为碳源,对肺炎克雷伯菌进行最适培养条件优化.结果表明:肺炎克雷伯菌最适培养条件:初始氮浓度为80 mg/L,碳氮比为10,温度为35 ℃,pH为6-7(见图1、图2).
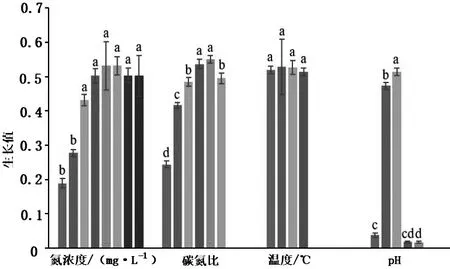
图1 不同培养条件肺炎克雷伯菌的生长值

图2 不同培养条件肺炎克雷伯菌对亚硝酸盐氮的消除率
2.5 好氧反硝化菌对模拟污水亚硝酸盐氮含量的影响
利用肺炎克雷伯菌菌液处理水产养殖污水,第2 d、5 d添加菌液的实验组亚硝酸盐氮的含量明显降低(图3),且实验组和对照组之间均存在显著性差异(P<0.05);结果表明筛选得到的肺炎克雷伯菌能有效降低水体中亚硝酸盐氮浓度.
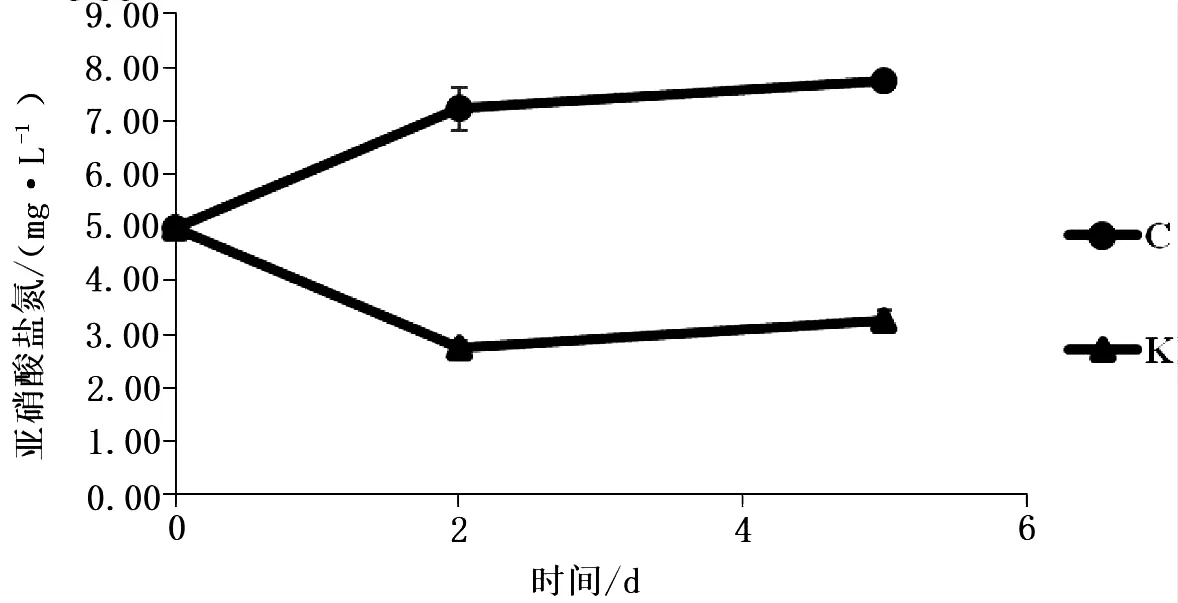
图3 降亚硝酸盐菌对养殖污水亚硝酸盐氮含量的影响
3 讨论
多年来,我国的水产养殖中养殖对象的活动、摄食、排泄都在同一池塘中进行,切断了自然生态系统的食物链[5],导致水产养殖的生态环境日趋恶化,氨氮和亚硝酸盐超标,对水产动物产生毒害作用[6].1972年,首次有报道得到1株具有异养硝化能力的节细菌属细菌(Arthrobactersp)[7].后来又发现粪产碱菌(Alcaligenesfaecalis)[9]、施氏假单胞菌(Pseudomonasstutzeri)[10]、铜绿假单胞菌(Pseudomonasaeruginosa)[11]等多个具有异养硝化-好氧反硝化的细菌.孙庆花等[12]从海底沉积物中分离筛选得到的1株耐盐异养硝化-好氧反硝化细菌被鉴定为肺炎克雷伯菌,研究表明,肺炎克雷伯菌具有良好的氮转化功效[13],在好氧条件下均能直接以NO3-N和NO2-N为底物进行反硝化,并且亚硝酸盐脱氮率高于硝酸盐脱氮率[14].本实验从中华鳖养殖水体筛选出一株肺炎克雷伯菌,能以亚硝酸盐为唯一氮源,利用亚硝酸还原酶,将亚硝酸盐转化为NO或NO2.亚硝酸还原酶分为两种,由nirS编码的血红素cd1型亚硝酸还原酶(cd1-NiR)和由nirK编码的铜型亚硝酸还原酶(Cu-NiR),在进行反硝化的分子生态学研究中主要以nirS或nirK基因作为分子标记.本实验从中华鳖养殖池筛选得到肺炎克雷伯菌,对菌种的亚硝酸还原酶基因进行分析,nirS基因为阳性、nirK基因为阴性,表明该为nirS+,nirK-菌.
已报道的好氧反硝化菌中,能利用亚硝酸盐氮为唯一氮源的很少.Wan等[15]研究表明,假单胞菌yy7随着培养基中初始亚硝酸盐氮浓度的增加,对亚硝酸盐的去除率降低;当初始亚硝酸盐氮浓度为50 mg/L,细菌生长缓慢,亚硝酸盐氮去除率小于50%.本实验中,在初始氮浓度从10 μg/L到80 μg/L时,肺炎克雷伯菌的生长速率随浓度的升高而增大,初始氮浓度在80 μg/L与140 μg/L时,菌株生长速率无显著差异.而在初始氮浓度为10 μg/L至120 μg/L时,肺炎克雷伯菌对亚硝酸盐氮的利用率在47%左右,当亚硝酸盐浓度达到140 μg/L时,菌株对其的利用率提高了大约20%左右.说明氮浓度太低时,菌体没有足够的氮源,不能很好地生长繁殖,脱氮能力低.因此,氮浓度在一定的范围内,菌株才能保持最高的脱氮能力.
C/N在反硝化过程中同样起着很重要的作用,Cervantes等[16]认为C/N是获得高效反硝化效率的主要控制参数.Kumar等[17]研究表明,当生物反应器的C/N为9∶3时,工业废水反硝化率和总氮去除率达到最大值.本实验中碳氮比对肺炎克雷伯菌的生长和脱氮效率的影响几乎一致,在一定范围内,随着碳氮比的增大,菌株的生长速率增大,且脱氮效果变好,当碳氮比达到10时,肺炎克雷伯菌的生长和脱氮效率达到最佳.碳源缺乏会导致菌株的生长繁殖受限,反硝化过程酶系合成不完全,使中间产物累积造成反硝化不彻底[18].同时反硝化关键酶硝酸盐还原酶的表达受到碳源的影响,碳源消耗越多,其酶活性越高.因此作为能源的有机碳浓度越高,反硝化速率越快;碳源不足,易引起不完全反硝化,导致高浓度的硝酸盐和亚硝酸盐残留[19].
阎胜利等[20]研究发现细菌生长及反硝化活性最适温度范围是25 ℃~35 ℃,当温度超过这一范围时,均会抑制细菌快速生长及反硝化性能的正常发挥.施氏假单胞菌(Pseudomonusstutzeri)在4 ℃~45 ℃的范围内均能正常生长,但在30 ℃时亚硝酸盐利用率最高,达5.53 mg/(L·h)[10];托拉假单胞菌(Pseudomonastolaasii)Y-11在15 ℃下可以在初始亚硝酸盐氮质量浓度高达209.62 mg/L的情况下以2.04 mg/(L·h)的速率去除93.6%氨氮[21].本研究结果表明,肺炎克雷伯菌在22 ℃至35 ℃的培养条件下,其生长速率无显著差异,但随着温度的升高,肺炎克雷伯菌对亚硝酸盐氮的利用率相应提高.可见,温度对细菌生长及反硝化性能发挥有重要的影响,这可能与菌株生长及反硝化酶的机制有关,需进一步研究.
王宏宇等[22]对好氧反硝化菌株C3研究表明,pH为中性条件最有利于菌株生长及反硝化性能的发挥.已有研究表明,细菌生长及反硝化酶活性的最适pH为中性或微碱性,pH过高或过低均会对菌株生长及反硝化性能的发挥产生影响[23].本实验中肺炎克雷伯菌在pH为6、7时,菌株生长状况良好且对亚硝酸盐氮的利用率也高,在偏酸和偏碱的环境中,酶活性受到巨大影响,不利于其正常进行反硝化作用[24].
4 结论
本研究筛选出具有降解亚硝酸盐氮的肺炎克雷伯菌,在以NaNO2为唯一氮源的培养基中,肺炎克雷伯菌最适生长条件为初始氮浓度80 mg/L,碳氮比10,温度35 ℃,pH 6~7.实验结果为进一步将肺炎克雷伯菌开发成降亚硝酸盐氮微生态制剂提供了实验基础.
