锡兰钩虫SLP-1 基因的克隆表达与免疫学特性分析
2022-03-22庄婷婷朱诗兰陈晓雨李国清
何 龙,赵 琪,庄婷婷,朱诗兰,陈晓雨,李国清
(华南农业大学兽医学院,广东 广州 510642)
钩虫是一类寄生于人和犬、猫等动物肠道中的土源性线虫,其感染性幼虫可通过皮肤与口腔感染宿主,引发人畜共患的钩虫病[1]。该病是一种极易被忽视,但呈世界性流行的肠道寄生虫病,在热带及亚热带经济落后地区的感染尤为严重,尽管该病的死亡率相对较低,但在一些贫穷落后的国家仍可造成健康隐患与严重的经济损失[2-3]。锡兰钩虫(Ancylostoma ceylanicum)是一种能够在人体内发育为成虫的动物源性钩虫,既可感染犬、猫也可感染人,研究表明,锡兰钩虫已成为亚洲地区感染人的第二大钩虫,感染后可引起腹痛、腹泻、营养不良和缺铁性贫血等症状[4]。多项研究证实锡兰钩虫已在中国、尼日利亚、柬埔寨等地的人和伴侣动物之间传播,尤其是在犬猫中高度流行,使得人感染锡兰钩虫的风险显著上升[5-6]。因此,研究控制该病的新方法已成为当务之急。本研究室一直致力于锡兰钩虫的分泌蛋白候选抗原的研究,前期已对谷胱甘肽-S-转移酶、血小板抑制剂和天冬氨酸蛋白酶抑制剂进行了克隆表达,并对其生物学功能进行了探究[7-9]。
鞘脂激活蛋白样蛋白(Saposin-like proteins,SLPs)又称皂素样蛋白,是由鞘脂激活蛋白原(Prosaposin)衍生的一类蛋白家族,具有改变细胞膜通透性及裂解红细胞等的作用。该蛋白广泛分布于各种原生动物及哺乳动物体内,目前已发现235 个家族成员,它们具有溶血、抗菌等多种生物学功能[10]。Don 等在犬钩虫(Ancylostoma caninum)中克隆表达了一种可以致红细胞破裂的蛋白,通过与肝片吸虫(Fasciola hepatica)FhSAP 基因序列的比对,证实其属于SLPs[11]。随后Willis 等完成了对美洲钩虫(Necator americanus)和犬钩虫SLPs 晶体结构的测定与功能分析[12]。但迄今未见对锡兰钩虫SLPs 的报道。基于此,本研究克隆锡兰钩虫Ace-SLP-1 基因,并对其编码蛋白的氨基酸序列进行生物信息学分析;将该基因经原核表达,对其表达产物进行免疫学特性分析,为进一步研究该蛋白的生物学功能及其作为钩虫疫苗候选分子的研究奠定基础。
1 材料与方法
1.1 犬源锡兰钩虫的获得锡兰钩虫三期幼虫(L3幼虫)采用离心管“T”型滤纸法从犬粪便中分离所得,成虫通过L3 幼虫人工感染金黄仓鼠而获得;所有虫体样品均参照文献[13]方法鉴定为锡兰钩虫,置于RNA 样品保存液中-20 ℃保存备用;锡兰钩虫感染犬和健康犬均由华南农业大学兽医学院临床外科教研室提供。
1.2 主要试剂pMD-19T 与pET-28a 克隆载体、BamH I、Hind III 均购自TaKaRa 公司;改良BCA 蛋白浓度测定试剂盒、CCK-8 试剂盒、兔抗犬IgGHRP、兔抗鼠IgG-HPR 和鼠His 标签单克隆抗体(MAb)均购自生工生物工程(上海)股份有限公司;E. coliDH5α、BL21(DE3)感受态细胞购自广州擎科生物科技有限公司;蛋白Marker 购自美国Thermo Fisher Scientific 公司;MicroElute Total RNA Kit、MMLV First Strand cDNA Synthesis Kit 和Gel Extraction Kit D2500 均购自广州欧米伽生物工程有限公司;His 标签纯化试剂盒购自碧云天生物技术(上海)公司;外周血淋巴细胞分离试剂盒购自天津灏洋华科生物科技有限公司。
1.3 Ace-SLP-1 基因的PCR 扩增与测序根据GenBank 中犬钩虫Ac-SLP-1 基因序列(DQ855414.1),设计特异性引物SLP-1-F:5'-ATGACTCGCTTGAT CTTCGTCGT-3'/SLP-1-R:5'-TCAACAAGCATGCA GAGTTGTGC-3'。采用Total RNA Kit 提取成虫总RNA,反转录合成cDNA 第一链作为模板,利用上述引物经PCR 扩增Ace-SLP-1 基因,反应条件:94 ℃5 min;94 ℃30 s、56.5 ℃30 s、72 ℃80 s,共35个循环; 72 ℃8 min。采用BamHI、Hind III 分别双酶切pMD19-T 和目的基因PCR 产物,经T4 连接酶连接,构建重组质粒pMD19-T-Ace-SLP-1,并经双酶切鉴定,阳性克隆由生工生物工程(上海)股份有限公司测序。
1.4 Ace-SLP-1 编码蛋白的生物信息学及同源性分析根据Ace-SLP-1 基因测序结果推导其氨基酸序列,利用DNAMAN 软件将其与11 种不同虫种SLPs 的氨基酸序列进行同源性比对。利用在线软件(https://web.expasy.org/protparam/)分析该蛋白的分子量大小、氨基酸组成、等电点(PI)等性质;分别利用在线软件SignalP(http://www.cbs.dtu.dk/services/SignalP/)、TMHMMServer2.0(http://www.cbs.dtu.dk/services/TMHMM/)和ProtScale(http://web. expasy.org/cgi-bin/protscale/protscale.pl)对该蛋白的信号肽序列、跨膜结构域与疏/亲水性进行分析;利用SWISS-MODEL预测软件(http://swissmodel.expasy.org/)预测并构建该蛋白的三维结构模型。
1.5 Ace-SLP-1 成熟肽基因(Ace-XL-SLP-1)重组质粒的构建及其重组蛋白(rAce-XL-SLP-1)的表达、纯化及鉴定根据1.4 的分析结果,选择锡兰钩虫Ace-XL-SLP-1 的成熟肽基因序列进行蛋白表达:设计扩增其成熟肽基因序列Ace-XL-SLP-1 的上下游引物XL-SLP-1-F:5'-AAGCTTTCAACAAGCAT GCAGAG-3'/XL-SLP-1-R:5'-GGATCCACTCCAGT AGTGATCAAC-3'。参照1.3 方法经PCR 扩增Ace-XL-SLP-1 基因,采用BamH I、Hind III 双酶切质粒pET-28a 和目的基因PCR 产物,并利用T4 连接酶连接,构建锡兰钩虫Ace-XL-SLP-1 成熟肽重组质粒pET28a-Ace-XL-SLP-1 并经双酶切鉴定,阳性克隆由生工生物工程(上海)股份有限公司测序。将pET28a-Ace-XL-SLP-1 转化大肠杆菌E. coliBL21(DE3),37 ℃、180 r/min 培养至OD600nm值为0.6 时,加入1 mmol/L IPTG 诱导表达6 h~8 h,4 ℃离心收集菌体,超声破碎后分别取上清和沉淀经SDS-PAGE电泳,分析重组蛋白的表达。将离心收集的包涵体置于8 mol/L尿素缓冲液中溶解过夜,离心收集上清,用0.22 μm滤膜过滤后,上样于Ni-NTA亲和层析柱,分别用10 mmol/L、20 mmol/L、40 mmol/L、50 mmol/L、80 mmol/L、100 mmol/L、200 mmol/L、500 mmol/L 的咪唑缓冲液洗脱,收集洗脱液及纯化后的蛋白,以鼠His 标签MAb(1∶3 000)为一抗,兔抗鼠HRP-IgG(1∶20 000)为二抗,同时以诱导的pET-28a/BL21(DE3)菌液与鼠His 标签MAb 孵育后作为阴性对照。经western blot 进一步分析rAce-XL-SLP-1 的纯化效果。
1.6 rAce-XL-SLP-1 的反应原性分析分别从锡兰钩虫感染犬和健康犬经前肢静脉无菌采血,分离血清。对纯化的rAce-XL-SLP-1 分别以感染犬血清(1∶4 000)和健康犬血清(1∶4 000)为一抗,兔抗犬IgG-HRP(1∶20 000)为二抗,通过western blot 分析rAce-XL-SLP-1 的反应原性。
1.7 rAce-XL-SLP-1 对犬外周血淋巴细胞(PBMC)增殖影响的检测从前肢静脉无菌采集健康犬EDTA抗凝血5 mL,采用外周血淋巴细胞分离液试剂盒分离健康犬PBMC,用台盼蓝染色计算活细胞数量,活细胞比率大于95%即可用于后续试验。取96孔细胞培养板,每孔加入100 μL PBMC悬液,再加入Ace-XL-SLP-1重组蛋白至终浓度分别为0、10 μg/mL、20 μg/mL、和40 μg/mL,同时设置ConA(刀豆球蛋白A)阳性对照和空白对照(细胞培养液),用细胞培养液将酶标仪调零,每个浓度3 个重复,置37 ℃、5% CO2培养48 h后,每孔加入10 μL CCK-8 溶液,继续培养4 h 后利用酶标仪测定其OD450nm值,并按公式计算PBMC 的增殖指数(SI),SI=实验组OD450nm值/对照组OD450nm值,SI>1.5 时,判为有效刺激。所得数据采用SPSS软件分析,两组间比较采用Studentt检验。
2 结 果
2.1 Ace-SLP-1 基因的PCR 扩增及其编码蛋白的生物信息学分析以锡兰钩虫cDNA 为模板经PCR扩增Ace-SLP-1 基因。结果显示,获得了全长为315 bp 的Ace-SLP-1 基因序列(图1)。测序结果显示锡兰钩虫Ace-SLP-1 基因全长315 bp,共编码104 个氨基酸。将犬源Ace-SLP-1 基因编码的氨基酸序列与其他虫种的SLPs 进行比对,结果显示,Ace-SLP-1 含有SLP 家族特有的6 个Cys-X-Cys 特征性序列,其与犬钩虫Ac-SLP-1 和Ac-SLP-2 编码氨基酸序列的同源性分别为95.28%和19.81%。利用在线软件对其氨基酸序列进行生物信息学分析,结果显示该蛋白分子质量和PI 分别为11.4 ku 和7.0;疏水性平均系数(GRAVY)大于0,推测其为疏水性蛋白(图1B);有跨膜结构域(图1C),且该蛋白前17 个氨基酸为信号肽序列(图1D);其三维结构模型(图1E)结果显示,Ace-SLP-1 编码蛋白的全球性模型质量估值(GMQE 值)为0.64(可信度范围为0~1,值越大表明质量越好),表明该蛋白三维结构模型建模质量良好。
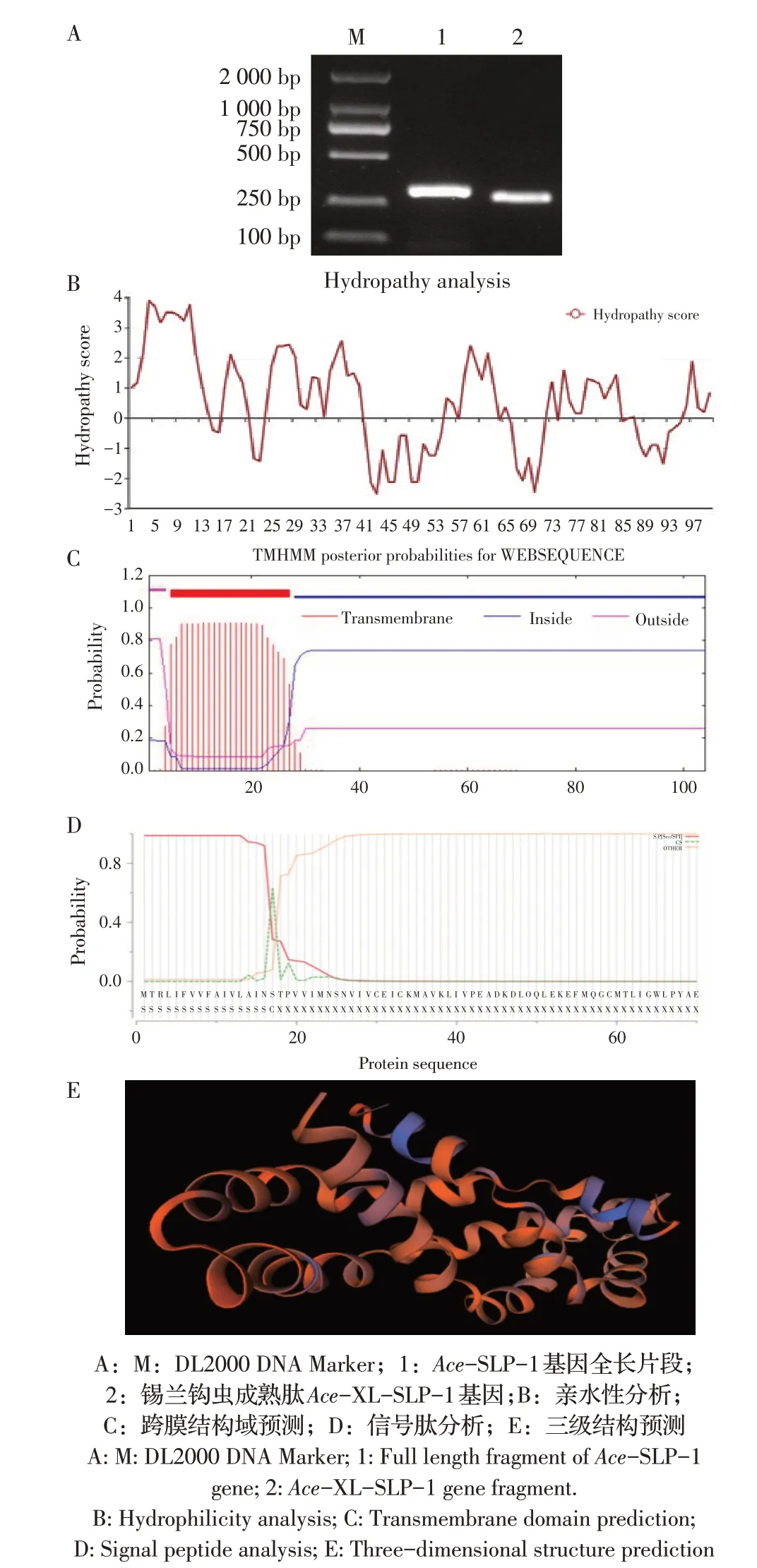
图1 Ace-SLP-1基因的PCR扩增及其编码蛋白的生物信息学分析Fig.1 PCR amplification of Ace-SLP-1 gene and bioinformatics analysis of Ace-SLP-1 protein
2.2 Ace-XL-SLP-1 重组质粒的构建与鉴定以锡兰钩虫cDNA 为模板经PCR 扩增Ace-XL-SLP-1,并克隆至pET-28a 中,构建重组质粒pET28a-Ace-XLSLP-1,经Hind III/BamH I酶切鉴定,结果显示获得了约260 bp 的目的条带,与预期目的片段相符(图2)。测序结果表明,正确构建重组质粒pET-28a-Ace-XL-SLP-1。

图2 pET-28a-Ace-XL-SLP-1重组粒双酶切鉴定结果Fig.2 Identification of recombinant plasmid by double enzyme digestion pET-28a-Ace-XL-SLP-1
2.3 锡兰钩虫Ace-XL-SLP-1 的表达、纯化及鉴定结果将重组质粒pET-28a-Ace-XL-SLP-1 转化大肠杆菌后,经IPTG 诱导表达并经Ni-NTA 亲和层析法纯化,将诱导前后菌液、破碎后沉淀、上清以及纯化蛋白经SDS-PAGE 检测,结果显示,在14.4 ku处出现目的条带,且该目的蛋白主要在包涵体中表达(图3A);western blot 结果显示,该重组蛋白能被鼠His 标签MAb 识别,在14.4 ku 处出现特异性条带(图3B);通过western blot 分析锡兰钩虫rAce-XL-SLP-1 的反应原性,结果显示该重组蛋白能够被感染锡兰钩虫的犬阳性血清特异性识别,在14.4 ku 处出现特异性条带,而健康犬血清与该重组蛋白反应后未出现该条带(图3C)。上述结果表明,锡兰钩虫重组成熟肽蛋白rAce-XL-SLP-1 具有良好的反应原性。

图3 rAce-XL-SLP-1表达、纯化的SDS-PAGE(A)及western blot(B、C)鉴定结果Fig.3 SDS-PAGE(A)and western blot(B,C)identification results of rAce-XL-SLP-1 expression and purification
2.4 Ace-XL-SLP-1 对PBMC 增殖影响的检测结果将不同浓度的rAce-XL-SLP-1与PBMC共孵育4 h后利用酶标仪测定其OD450nm值。结果显示,与阴性对照相比,20 μg/mL 和40 μg/mL 的rAce-XL-SLP-1 对PBMC 均具有明显的刺激增殖作用(P<0.05),且与其浓度有关,当蛋白浓度为40 μg/mL 时,SI 最大(2.15),与阳性对照组的SI 相当。当蛋白浓度低至10 μg/mL 时,刺激效果不明显(表1)。表明,获得的rAce-XL-SLP-1 具有一定的免疫原性。

表1 rAce-XL-SLP-1对犬PBMC增殖效应的检测结果Table 1 Detection results of the effect of recombinant protein Ace-XL-SLP-1 on canine PBMC proliferation
3 讨 论
钩虫被世界卫生组织列为易被忽视的主要土源性线虫之一[14]。我国多次对家养犬进行的蠕虫流行病学调查时发现,锡兰钩虫是寄生在犬体内的优势虫种,严重危害犬的健康[15]。目前主要采取药物治疗和环境卫生措施防治钩虫病,但由于钩虫抗药性的产生和重复感染使其疗效不佳。近年来,研究人员试图从钩虫致病机理角度来探究该病新的防治方法,其中能够改变细胞膜通透性并使红细胞破裂的SLPs 成为研究热点。
本研究首次从锡兰钩虫成虫克隆了Ace-SLP-1的cDNA 序列。通过对其氨基酸序列的生物信息学分析发现,该蛋白前17 个氨基酸为信号肽序列,不利于该重组蛋白的表达。为了提高其表达量,本研究设计了一对引物扩增Ace-SLP-1 基因的成熟肽序列,并在E. coliBL21(DE3)菌株中表达出了该蛋白。同源性分析发现该蛋白的氨基酸序列含有6 个保守的半胱氨酸残基,可参与二硫键的形成,对高热下多肽的稳定性具有重要作用[16]。可溶性分析发现该蛋白主要在大肠杆菌中以包涵体的形式表达,可能由于其与二硫键配对的半胱氨酸含量较多,所以易形成包涵体。
SLPs 是一类作用于细胞膜的蛋白,具有作为疫苗候选分子或诊断分子的潜力。据报道采用肝片吸虫的重组SLP 蛋白(rFh-SAP-2)免疫小鼠,可获得81.2%的减虫率,显示其是一种潜在的疫苗候选分子[17];而曼氏血吸虫和日本血吸虫的SLPs 具有良好的反应原性,可以被感染动物(小鼠和兔)的血清和人血清所识别[10],具有潜在的血清学诊断意义[18]。本研究结果显示经原核表达的rAce-XL-SLP-1 能够被锡兰钩虫感染的犬阳性血清所识别,显示其具有作为锡兰钩虫血清学诊断的潜在候选抗原。
近年来,CCK-8 试剂盒因检测灵敏度高,操作简单,无需有机溶剂,对细胞无毒性,已广泛用于细胞增殖的快速检测。通过该试剂检测经不同浓度rAce-XL-SLP-1 刺激后的犬PBMC 的增殖效应能够反映该重组蛋白的细胞免疫功能,在有效刺激(SI>1.5)的情况下,SI 越大,淋巴细胞群体的反应能力和免疫功能越强[19]。Huang 等通过该试剂盒的检测结果证实锡兰钩虫血小板抑制剂(Ace-HPI)能够刺激犬PBMC 的增殖和细胞因子的分泌,有助于钩虫在宿主体内建立慢性感染[20]。本研究分别将不同浓度的rAce-XL-SLP-1 与PBMC 共孵育时,当该重组蛋白浓度为20 μg/mL 和40 μg/mL 时,PBMC 的SI 明显高于对照组(P<0.05)。表明rAce-XL-SLP-1 能够诱导犬PBMC 的增殖,具有良好的免疫原性,提示rAce-XL-SLP-1 具有作为钩虫疫苗候选抗原分子的潜力。
综上所述,本研究克隆了锡兰钩虫Ace-SLP-1基因,对其编码的氨基酸序列进行了生物信息学分析后,构建了该基因成熟肽的重组质粒pET-28a-Ace-XL-SLP-1 并诱导表达与纯化了rAce-XL-SLP-1,对其免疫学特性进行了初步分析,为钩虫病的血清学诊断及钩虫疫苗候选分子的筛选与功能学等研究奠定了基础。
