矩镰荚苜蓿越冬期地下芽的组织化学特征与分化规律
2022-03-21杨小霞魏甲科夏建强方强恩
杨小霞,魏甲科,夏建强,方强恩,张 勃
(甘肃农业大学草业学院, 甘肃 兰州 730070)
芽休眠(bud dormancy)是植物芽端分生组织随环境变化在活跃生长和休眠状态之间转变的动态过程[1]。在冬季,植物的芽能以休眠的方式应对严酷的低温胁迫,是其经过长期演变而获得的一种环境适应能力[2]。多年生草本植物主要以地下芽休眠的方式进行越冬[3]。草本植物的越冬芽在休眠发生时,通常会表现出芽端分生组织生长分化停止、芽内部物质(如碳水化合物、脂类、蛋白质)代谢减慢等一系列生理特性的变化[2]。为适应特定的气候条件,一些植物的越冬芽还会形成特征各异的组织结构与生长节律。例如,茭白(Zizania latifolia)冬芽在越冬期间会形成特殊的维管结构,其芽生长锥和幼叶细胞分裂活动减弱,代谢活动减缓,同时细胞内的淀粉颗粒开始积累增多[4]。芍药(Paeonia lactiflora)的地下芽在越冬前(秋季)能快速发育,并完成其营养生长期;在临近土壤封冻时,地下芽进入休眠状态,随之芽内的可溶性糖含量开始增加[5-7]。目前,有关多年生草本植物地下芽休眠的研究主要集中在人工栽培植物,关于青藏高原高寒草地多年生野生草本植物地下芽休眠特性的研究鲜有报道。
矩镰荚苜蓿(Medicago archiducis-nicolai),又称青藏扁蓿豆,是豆科苜蓿属多年生野生优质牧草,分布于我国西部高寒地区,其自然生境多处于冬季少雪或无雪覆盖的极端寒冷区域,耐寒性极强[8-10]。研究表明,矩镰荚苜蓿在每年8 月 - 9 月会形成许多地下越冬芽,这些芽能在极端的低温条件下安全越冬,对寒冷胁迫具有极强的耐受性[9,11]。目前,关于矩镰荚苜蓿的研究主要涉及地理分布、栽培技术[12-13],营养价值的评价[14]、遗传多样性评价[9-10]等方面,对地下芽在自然越冬过程中的组织化学特征与分化及休眠的研究尚未见报道。本研究采用石蜡切片方法,通过组织化学染色,跟踪观测了矩镰荚苜蓿自然越冬过程中地下芽的组织结构与生长变化特征,以期为理解该植物的耐寒越冬机理提供基础依据。本研究拟回答两个问题:1)矩镰荚苜蓿越冬期地下芽的组织化学特征和芽尖分化特征会发生哪些变化?2)越冬前后地下芽的休眠态是如何转变的?
1 材料与方法
1.1 试验材料
本研究材料为青藏高原东缘自然种群中随机采集的单株个体(芽),采样地点位于天祝县甘肃农业大学高山草原试验站禁牧草地,该地区海拔2 071 m,年均温-8~14 ℃,土壤类型为亚高山草甸土,气候类型为大陆性高原季风气候、大陆性半干旱气候。
1.2 试验方法
试验在2018 年9 月至2019 年5 月进行,根据试验地日均气温的变化规律(图1),参考Pearce[15]、Taylor 等[16]对植物低温胁迫的划分方法,将矩镰荚苜蓿越冬过程分为4 个时期:秋末冷适应期(coldadaptation stage in late autumn, CA,日均气温在5 ℃以上);初冬冷胁迫期(cold stress stage in early winter,CS,日均气温0~5 ℃);隆冬冻胁迫期(freezing-stress stage in mid-winter, FS,日均气温降至0 ℃以下);返青期(spring growth stage, SS,日均气温回升至5 ℃以上)。根据试验期试验地日平均气温的变化,4 个时期共设8 次取样。由于低温胁迫期(冷胁迫期和冻胁迫期)是植物休眠态转变的关键期[17-18],为了揭示矩镰荚苜蓿越冬芽休眠态的转变规律,在初冬冷胁迫期设两次取样(2018 年10 月27 日、2018 年11 月10 日),在隆冬冻胁迫期设4 次取样(2018 年11 月24 日、2018 年12 月26 日、2019 年1 月12 日、2019 年3 月24 日),在秋末冷适应期和返青期各设1 次取样,分别是2018 年9 月8 日和2019 年5 月5 日。
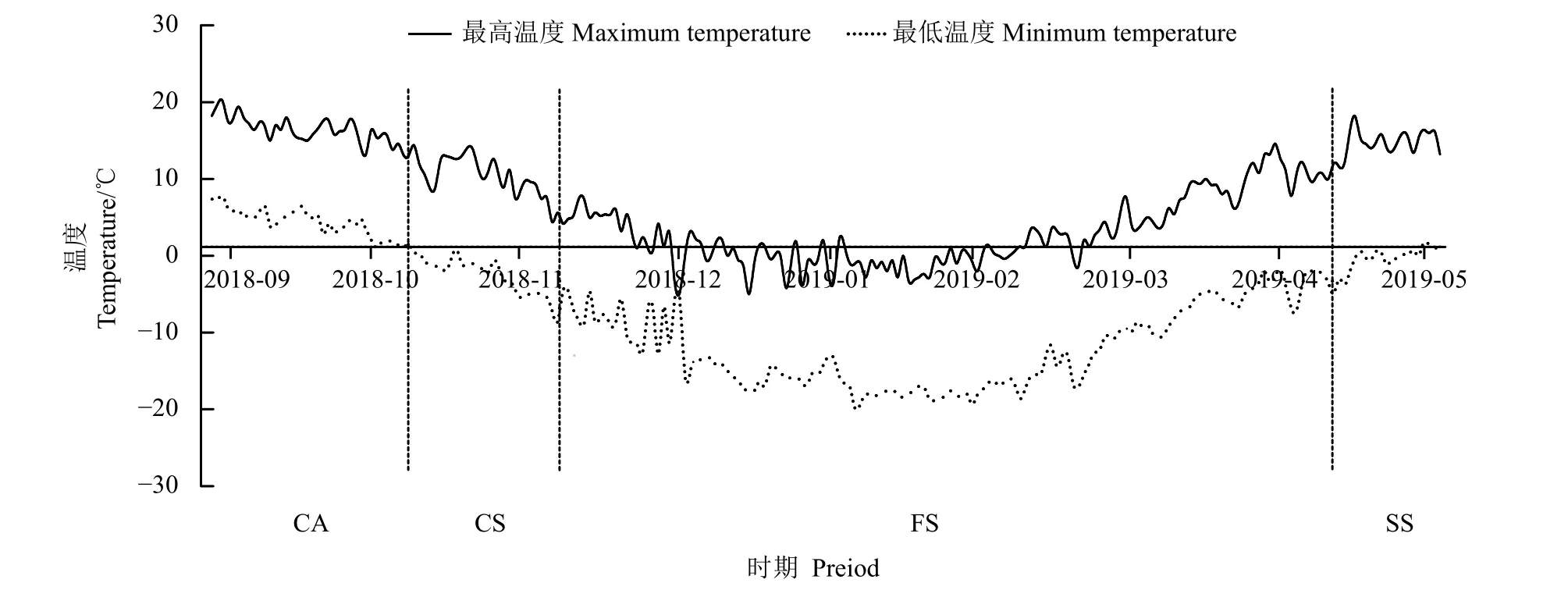
图1 试验期内(2018 年9 月至2019 年5 月)日最高温、日最低温Figure 1 Daily maximum temperature, daily minimum temperature (September 8, 2018, to the solstice on May 5, 2019)
在每个时期取样时,用铁锹、锄头挖取试验材料,迅速清洗干净,并用剪刀随机剪取生长健康、长势一致的地下芽10 个,并采集切取顶端长约5 mm片段,立刻用FAA 固定液(70%乙醇 : 冰乙酸 : 甲醛 =18 : 1 : 1)固定,带回实验室4 ℃保存,以备后期石蜡切片。
石蜡切片采用常规法[19],厚度8 μm。淀粉和蛋白质染色用高碘酸 - 锡夫反应法(Periodic acid -Schiff reaction, PAS)[20],淀粉在碱性品红染色下呈玫红色,蛋白质在萘酚黄染色下呈黄色。用Motic 生物显微镜(型号:Panthera U)观察地下芽各组织结构以及淀粉粒和蛋白质分布情况,拍照。淀粉颗粒的观察,每次取3 个视野,每个视野5 个细胞。不同时期地下芽顶端生长锥、芽尖、芽体的高度和宽度值,采用Motic 生物显微镜自带测量工具测量。
1.3 数据处理
采用SPSS 20.0 软件对所测数据进行统计分析,用平均值 ± 标准误表示测定结果,用最小极差法(LSD)进行差异显著性分析。并采用Sigmaplot 10.0和Office Excel 2013 制图。
2 结果与分析
2.1 矩镰荚苜蓿地下芽结构组成
根据地下芽解剖结构(图2),把芽最顶端长为1~2 mm 的区段划分为芽尖和芽体两个功能区。芽尖是分生分化区,由生长锥(分生区)和芽尖基层组织(分化区)组成。在芽尖基层组织内部看不到形成层,外部分化有2~3 个叶原基,没有腋芽原基。芽体是伸长区,从第一个腋芽原基分化节开始到第一个伸长节,在这一功能区段,内部组织已经分化,可以明显看到皮层、髓与形成层等维管组织;叶原基分化为幼叶,伸长至芽尖顶端,顶部向内收拢;节间伸长,腋芽原基分化为腋芽。比较不同越冬期芽组织后发现,在冬季,生长锥、芽尖和芽体的高、宽均发生了变化(图3、图4)。
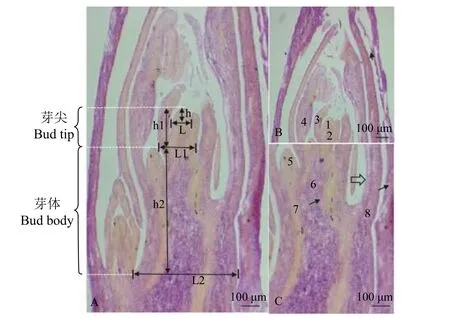
图2 矩镰荚苜蓿地下芽解剖结构(纵切)Figure 2 Anatomical structures of the belowground buds of Medicago archiducis-nicolai (longitudinal sections)
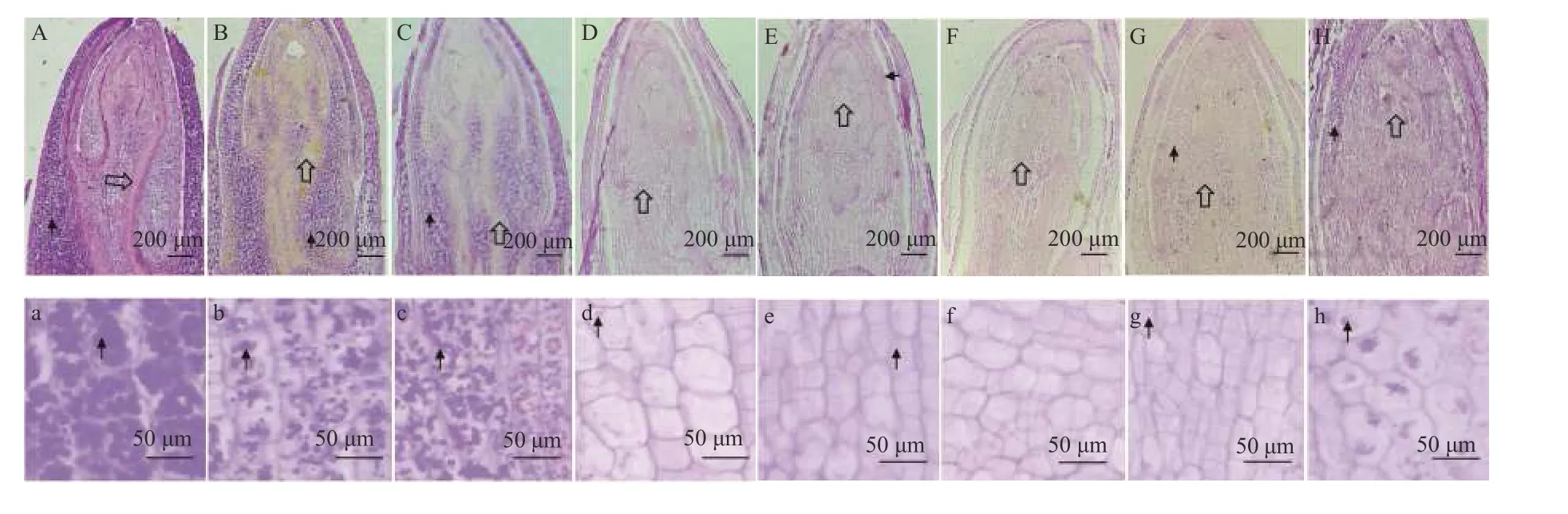
图3 矩镰荚苜蓿地下芽细胞内淀粉粒和蛋白质的分布Figure 3 The distribution of starch and protein in cells of the belowground buds of Medicago archiducis-nicolai
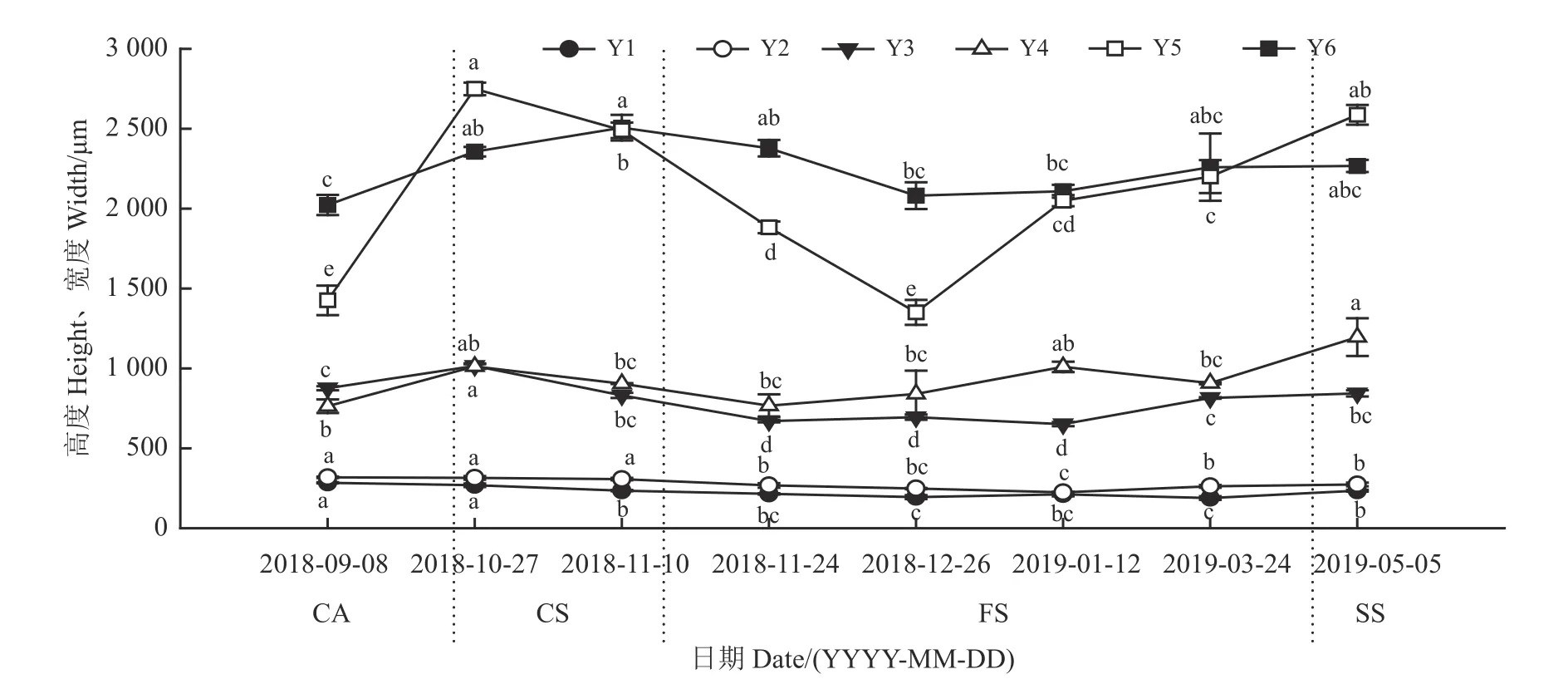
图4 越冬期矩镰荚苜蓿地下芽芽端组织生长变化Figure 4 Development of the terminal tissues of belowground buds of Medicago archiducis-nicolai during overwintering
2.2 矩镰荚苜蓿地下芽组织内淀粉粒和蛋白质的分布及数量变化
越冬期间,矩镰荚苜蓿芽尖、芽体细胞内淀粉粒和蛋白质分布发生变化(图3、图5)。淀粉粒主要分布在芽尖基层组织、芽尖叶原基底部、芽尖外围幼叶、芽体髓部和芽体皮层等部位。蛋白质主要分布在芽尖生长锥、芽尖叶原基和芽形成层等部位。冷适应期和冷胁迫期,随日均气温的降低,矩镰荚苜蓿地下芽组织细胞内的淀粉粒和蛋白质均逐渐减少。不同的是,淀粉粒集中排列在薄壁组织细胞中心,而蛋白质的排列没有明显的规律(图3A-C)。冻胁迫期,随日均气温的降低,淀粉粒几乎消失殆尽,随后不再随气温的降低而变化,零散出现在组织细胞中,该时期蛋白质的积累极少(图3D-G)。返青期,随日均气温的逐渐回升,淀粉粒开始逐渐增多。相较于冷适应期和冷胁迫期,其排列不集中。该时期蛋白质的含量变化不显著(P> 0.05) (图3H)。
矩镰荚苜蓿地下芽在越冬过程中,淀粉粒的数量在冻胁迫期较其他时期低。在越冬期的某一特定时期,地下芽不同组织部位储存淀粉粒的数量存在差异。同一时期,芽尖基层组织淀粉粒较其他组织细胞内少,芽体髓部和皮层(图3a - h)居多,芽尖叶原基细胞内淀粉粒始终少于芽尖外围幼叶细胞内淀粉粒(图5)。不同组织部位细胞内淀粉粒数量在越冬期的变化趋势是一致的。冷适应期、冷胁迫期和冻胁迫前期,组织内淀粉粒数量显著降低(P<0.05);冻胁迫中期,淀粉粒数量变化平缓;冻胁迫后期和返青期,淀粉粒数量开始缓慢增加。冷适应期,蛋白质的含量较高,越冬其他时期的蛋白质含量变化不明显。
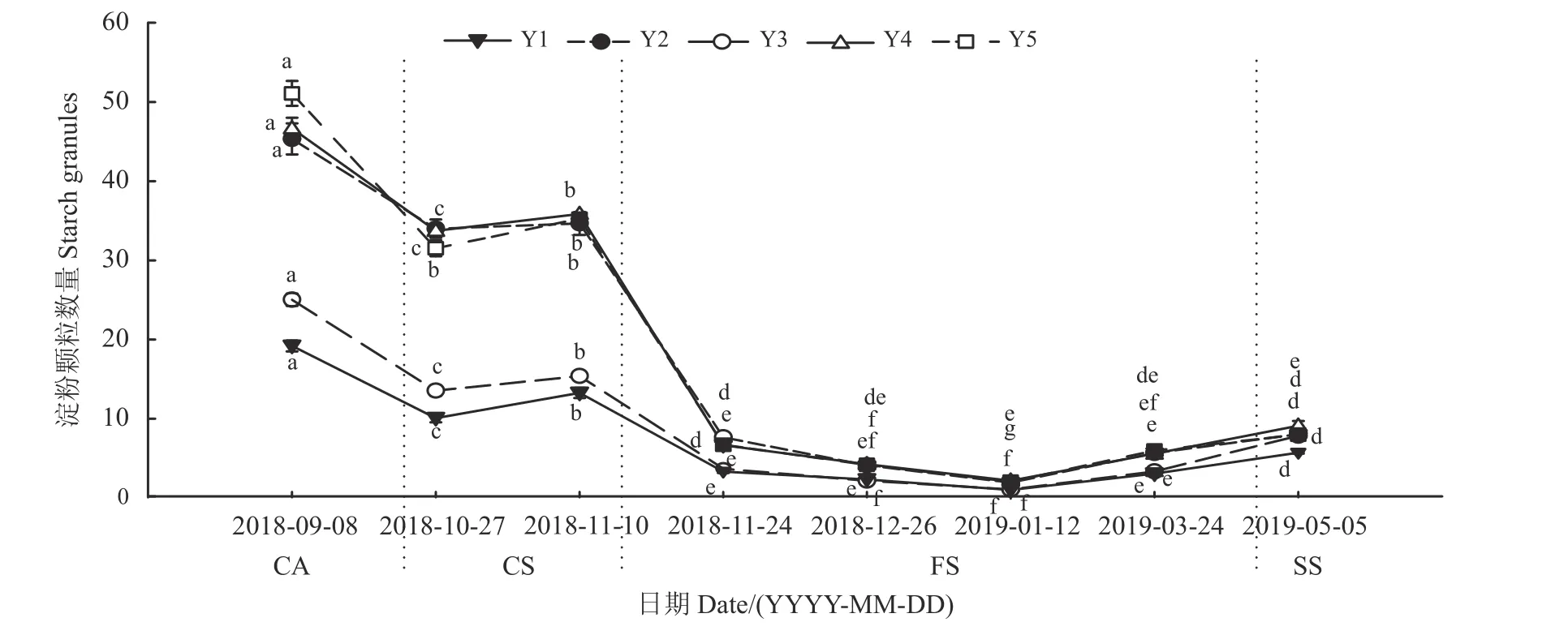
图5 越冬期矩镰荚苜蓿地下芽细胞内淀粉粒数量的变化Figure 5 Variation in the number of starch grains in cells of the belowground buds of Medicago archiducis-nicolai during overwintering
2.3 矩镰荚苜蓿地下芽芽端组织分化特征
矩镰荚苜蓿地下芽在越冬期的生长发生变化(图4)。冷适应期,随日均气温降低,芽尖和芽体的伸长和增粗显著增加(P< 0.05)。然而,生长锥的伸长和增粗变化不明显。冷胁迫期,随日均气温的降低,生长锥和芽体的增粗变化不显著(P >0.05)。冻胁迫期,日均气温降到最低并保持稳定状态,生长锥、芽尖的伸长和增粗以及芽体的增粗变化平缓,只有芽体的伸长显著增加(P< 0.05)。返青期,随日均气温的回升,地下芽各部位的增粗和伸长开始缓慢增加。
矩镰荚苜蓿地下芽在越冬前后,芽体部位会形成腋芽或腋芽原基。越冬不同时期,腋芽和腋芽原基的发育进度不同(图6)。冷适应期和冷胁迫期,芽体部位形成腋芽,且腋芽已分化出芽尖生长锥和叶原基(图6A-C)。冻胁迫前期和中期,在芽体部位分化出腋芽原基(图6D-F),但腋芽原基未进一步分化出芽尖生长锥和叶原基。冻胁迫后期和返青期,芽体部位又逐渐恢复分化生长,形成了腋芽并分化出叶原基(图6G、H)。

图6 越冬期矩镰荚苜蓿地下芽芽体部位腋芽原基的发育变化Figure 6 Developmental changes in the axillary primordia of the belowground buds of Medicago archiducis-nicolai during overwintering
3 讨论
3.1 矩镰荚苜蓿越冬期地下芽组织化学特征变化
淀粉是植物体内碳水化合物贮藏的主要形式,主要存在于植物的种子和根茎中。研究表明,淀粉的积累贮藏是植物越冬的先决条件。植物可通过淀粉的水解,增加细胞内的可溶性糖含量,以此提高细胞液浓度,降低植物细胞的冰点,从而提高其抗寒力[21-22]。本研究结果显示,矩镰荚苜蓿地下芽在秋末冷适应期,随气温的下降,其淀粉粒数量显著降低,在隆冬时期芽内的淀粉粒几乎消失;在返青期,芽内的淀粉粒又开始缓慢积累。这与前人研究结果相类似,例如,刘芳等[23]对百合(Lilium pumilum)鳞茎花芽分化的研究发现,在温度较高时,百合鳞茎中淀粉含量较高且变化平缓,随着温度的降低,百合鳞茎中淀粉含量急速下降。同样,Nagao 等[24]对小立碗藓(Physcomitrella patens)的研究发现,该植物随低温胁迫,其原丝体细胞内的淀粉含量也呈现出降低趋势。研究表明,受低温诱导后,植物体内淀粉含量的水解与淀粉酶活性的提高密切相关,因为温度降低能激活植物体内的淀粉酶活性,从而加快了淀粉水解成葡萄糖的速率;反过来,糖的增加可以提高细胞液浓度,从而降低植物组织结冰的冰点,以此抵御寒冷环境的胁迫[23,25]。
可溶性蛋白质是植物体内另一种重要的渗透调节物质,能在逆境下大量积累,以此提高细胞浓度、增强细胞保水能力,从而提高植物的抗逆性[26]。例如,王颖等[27]对彩叶芋(Caladium bicolor)叶片以及魏娜等[28]对宿根花卉根系低温胁迫的研究均表明,随着温度降低,这些植物体中均会积累大量的可溶性蛋白质。本研究中,矩镰荚苜蓿在秋末冷适应期,其地下芽内积累了较多蛋白质,但是,在整个越冬期芽内的蛋白质含量无明显变化;也就是说,该植物的地下芽在越冬期并未通过改变可溶性蛋白质含量应对低温胁迫。同样,在拟南芥(Arabidopsis thaliana)和樟子松(Pinus sylvestris)幼苗中也发现,可溶性蛋白质的含量不会因为低温胁迫而发生显著的变化[29-30]。有研究认为,在低温胁迫下,植物体内的蛋白质可能参与了多个代谢过程,从而导致其积累少、含量相对较低[31]。由此可见,不同植物在应对低温胁迫时,其蛋白质代谢响应机制不尽相同。
3.2 矩镰荚苜蓿越冬期地下芽休眠态的变化
休眠是指植物外表停止生长,而体内仍然进行着最低限度的生理代谢活动的现象[32]。Rohde 和Bhalerao[33]从植物组织发育的角度认为,休眠是植物在不良环境或季节来临之时,其分生组织结构极度减缓或者暂时停止的一种状态,以此适应恶劣的外界环境。根据Rohde 和Bhalerao[33]的观点,研究者可通过越冬期跟踪观测芽内分生组织的生长变化判断出植物的休眠状态。本研究发现,矩镰荚苜蓿地下芽的生长锥、芽尖、芽体以及芽体腋芽的生长分化在越冬前后发生明显变化。在秋末冷适应期,地下芽的伸长和增粗显著增加,芽体部位腋芽分化程度高;冷胁迫期,地下芽的伸长和增粗显著减小,芽体部位腋芽分化程度高;冻胁迫期,地下芽增粗和伸长变化平缓,地下芽生长几乎停滞,芽体部位只分化出腋芽原基;返青期,地下芽增粗和伸长显著增加,地下芽又开始生长,芽体部位又分化出了腋芽。综合分析表明,矩镰荚苜蓿地下芽在越冬前后,共经历了快速生长期(10 月下旬之前)、休眠进入期(11 月)、休眠保持期(12 月至1 月中旬)、休眠解除期(1 月 - 4 月) 4 个阶段,从休眠进入到休眠解除长达6 个月。另外,有研究表明,植物休眠的起始与体内淀粉含量的变化密切相关[34-35]。本研究中,有关矩镰荚苜蓿地下芽内淀粉含量在不同时期的变化结果也支持了该观点。矩镰荚苜蓿芽组织细胞内的淀粉含量在越冬前快速生长期处于较高水平,进入休眠后随休眠态的转变,淀粉含量显著下降,在休眠保持期接近于零,随休眠解除,淀粉粒数量又开始逐渐积累上升。赵长山等[36]对苣荬菜(Sonchus brachyotus)芽根休眠过程的研究也发现了类似的结果。一方面,如前面所讨论,越冬期间地下芽内淀粉含量的这种变化可能与细胞调节渗透压、降低组织冰点以抵御低温胁迫有关;另一方面,芽休眠发生后,淀粉能通过水解过程为休眠解除提供物质基础和能量需要[37-38]。由此表明,高寒草地矩镰荚苜蓿地下芽在越冬期间能通过进入休眠状态抵御低温胁迫,同时通过水解芽内淀粉以满足其维持生命的物质和能量需要。
4 结论
矩镰荚苜蓿在越冬前地下芽基本组织内会积累大量淀粉,芽生长快速;进入冬季,芽尖分化与伸长生长开始减慢,随低温胁迫,地下芽内储存的淀粉粒快速降解,芽生长停止;翌年返青期(5 月),芽开始恢复生长。矩镰荚苜蓿越冬期间地下芽生长停滞和芽内淀粉含量降低是其休眠态转变的重要标志,休眠芽通过淀粉粒水解代谢,增加细胞可溶性糖浓度、降低细胞冰点是其抵御冬季低温胁迫的适应机制。
