退化程度对玛沁高寒草甸植物群落及土壤持水能力的影响
2022-03-21张宇恒张秀娟杨永胜王军邦徐兴良黄煜茹赵求东李英年周华坤
张宇恒,张 莉,张秀娟,杨永胜,王军邦,徐兴良,黄煜茹,赵求东,李英年,周华坤
(1. 长江大学园艺园林学院, 湖北 荆州 434025;2. 中国科学院西北高原生物研究所 / 青海省寒区恢复生态学重点实验室,青海 西宁 810001;3. 中国科学院高原生物适应与进化重点实验室,青海 西宁 810001;4. 中国科学院地理科学与资源研究所 / 生态系统网络观测与模拟重点实验室,北京 100101;5. 中国科学院西北生态环境资源研究院, 甘肃 兰州 730000)
三江源区位于我国青海省南部,平均海拔3 500~4 800 m,是长江、黄河、澜沧江的发源地,素有“中华水塔”之美誉[1],是我国重要的生态安全和水源涵养功能区。三江源区高寒草甸面积1.279 × 105km2,占该地区总土地面积的78.72%[2],其分布区域广阔,发育环境多样,在水源涵养、生物多样性保护和碳汇等方面起着不可替代的生态屏障作用[3]。然而,自20 世纪90 年代开始,受气候变暖、鼠虫危害、冰川缩减、冻土退化及人类活动加剧等诸多因素的综合影响,三江源高寒草甸区域发生了不同程度的退化,这导致该地区水土流失日趋严重、土地贫瘠化、物种多样性减少、土壤结构遭到破坏、草原鼠害猖獗及草地沙化面积不断扩大等一系列生态问题[4],不但危及区域生态安全,而且影响牧区经济的可持续发展,给当地牧民生产生活带来了巨大影响。随着全社会对生态环境问题的日益重视,三江源高寒草甸退化问题受到了生态学领域的广泛关注[5]。
目前,相关学者对草地退化的成因、演替过程、影响因素以及恢复措施等进行了较为广泛的研究[6-7]。研究发现,随着退化程度的增加,土壤抗侵蚀能力、土壤有机质、全氮及速效钾含量呈先增加后降低的趋势[8-9],植被地上、地下生物量及土壤微生物数量显著下降,在真菌数量和脲酶活性方面尤为显著[10]。在极度退化阶段呈现“黑土滩”地貌[11]。土壤持水能力是评价土壤水分调节能力和涵养水源的重要指标,与土壤结构等指标密不可分[12]。目前,有关高寒草甸退化过程中的研究多集中在土壤理化性质、土壤水文特征、土壤酶活性及微生物等方面[4,12-14],鲜有涉及高寒草甸植被群落结构与水源涵养方面的研究,综合分析不同退化程度高寒草甸的植物群落特征与土壤持水能力特征,对于了解高寒草甸退化演替的生态学过程、诊断草地生态健康具有重要意义,可便于管理部门采取合理的管理措施以防止高寒草地生态进一步恶化。
为此,本研究选取玛沁高寒草甸不同退化程度样地,结合野外调查和室内分析,量化退化程度对该区域植被群落结构和土壤持水特征的影响,以期为揭示三江源高寒草甸退化过程中植被演替及生态功能变化提供基础数据和理论依据。
1 研究区概况与研究方法
1.1 研究区域概况
研究区位于玛沁县大武乡德尔尼铜矿附近(34°23′27.6″ N, 100°15′28.8″ E),海拔3 780 m,气候为典型高原大陆性气候,无四季之分,有冷暖季之别。年平均温度为-0.4 ℃,其中最低气温在1 月,为-12.4 ℃;最高气温在7 月,为9.8 ℃[15]。年均降水量为508.5 mm,全年60%的降水量集中在6 月 - 8月。土壤类型为高山草甸土和高山灌丛土。该区常见的优势种有矮生嵩草(Kobresia humilis)、垂穗披碱草(Elymus nutans)、异针茅(Stipa aliena)。伴生种有高山嵩草(K. pygmaea)、细叶亚菊(Ajania tenuifolia)、西伯利亚蓼(Polygonum sibiricum)、附地菜(Trigonotis peduncularis)、委陵菜(Potentilla)等[16]。该区域植株低矮,生态系统脆弱,易受扰动。
1.2 研究方法
1.2.1 试验设计
试验于2020 年8 月开展,依据退化草地评价等级标准(原生植被盖度 > 80%,轻度退化盖度70%~85%,重度退化盖度30%~50%)[15],选取3 个退化梯度的样地,依次为原生植被、轻度退化、重度退化,样地间距约20 m。每个样地设置0.5 m × 0.5 m 群落样方和分种样方各3 个,即每个处理3 个重复,样方间距为5 m。
1.2.2 植被群落调查和样品采集
植物群落调查:在各样方内,用直尺测量地上植物群落的平均高度(在每个样方内测定10 次记录,取平均值)和单种的植株高度,用针刺法测定植被群落的总盖度和单种植被的分盖度。
地上生物量:首先进行群落样品采集,在样方内先分拣出凋落物,再用剪刀齐地面剪去植被地上部分,分别装入信封并标记。同时,在分种样方内进行分种样品采集,将刈割采集的每个植物种分别装袋,标记。将群落生物量和分种生物量样品均带回实验室,置于65 ℃烘箱中烘至恒重,称重,得到群落和分种地上生物量。
地下生物量和土壤:在群落样品采集样方内,用直径为7 cm 的土钻分层(0 - 5、5 - 10、10 - 20、20 -30、30 - 50、50 - 70 cm)采集植物根系和土壤,每个土层采集3 钻样品合为一体,然后用2 mm 筛子将土壤和植物根系分离标记,将取得的土样带回实验室备用。根系经清洗后装入信封,在65 ℃的烘箱内烘至恒重,称重。
土壤容重:采用环刀法,在分种群落样方内,挖取1 m 深的土壤剖面,用直径为5 cm、高5 cm 的环刀分层(0 - 5、5 - 10、10 - 20、20 - 30、30 - 50、50 -70 cm)采集原状土壤,带回实验室,测定土壤容重、持水能力及饱和导水率。
1.2.3 群落物种多样性
物种多样性指数(species diversity index, H′)、优势度(Simpson, C)、均匀度(eveness, J)、重要值(important value, IV)的计算公式如下:

1.2.4 土壤持水能力及容重
土壤持水量:首先将采集装有原状土的环刀揭去上、下底盖,仅留一垫有滤纸的带孔底盖,放置平底盆内注入并保持盆中水层高度至环刀上沿为止,使其吸水达12 h,此时环刀土壤中所有非毛管孔隙及毛管孔隙均充满了水分,盖上上、下底盖,水平取出,擦干环刀外沾的水,称取重量(M1);然后去掉底盖,放置在铺有干砂的平底盘中2 h,此时环刀中土壤的非毛管水分已经全部流出,但环刀中土壤的毛细管仍充满水分,盖上底盖,立即称重(M2);再将上述称量M2的环刀,揭去上、下底盖,继续放置于铺有干砂的平底盘中48 h,盖上上、下底盖称重(M3)。按照下列公式分别计算土壤饱和持水量、毛管持水量及田间持水量:

式中:W1为土壤饱和持水量(%);W2为土壤毛管持水量(%);W3为土壤田间持水量(%);M为环刀重量(g);B为环刀内烘干的土壤重量(g);A为滤纸重量(g);M1为吸水12 h 的土壤和环刀的重量(g);M2代表放在平盘中2 h 的土壤和环刀的重量(g);M3代表放在平盘中48 h 的土壤和环刀的重量(g)。
土壤容重:将采集的环刀放置于105 ℃的烘箱内,烘干至恒重称重,按照下列公式计算:
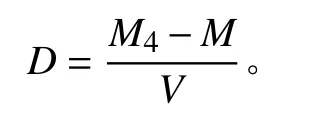
式中:D为土壤容重(g·cm-3);M4为烘干的土壤和环刀总重量(g);M为环刀重量(g);V为环刀体积(cm3)。
土壤饱和导水率:采用定水头法测定土壤饱和导水率。将原状环刀土样有孔盖一端朝下,覆上滤纸,盖上环刀孔盖,浸入水面与环刀上口接近平齐的水中,浸泡8 h 之后将环刀取出,揭去环刀盖,用胶带将其与另外一个空环刀连接,并放置在已架设好铁架台的三角漏斗上,马氏瓶出水口距环刀内土壤表面的高度为3 cm。安装完成后,马氏瓶开始供水,使土壤表层形成3 cm 的稳定水层,持续供水4 h 后测定,每隔5 min 记录渗出水量,同时记录水温。当最终测定的渗出水量基本不变或者循环出现时,停止测定。取最终稳定值计算饱和导水率(Ks)。
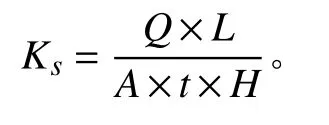
式中:Q为一段时间内的流量(cm3);L为土柱高度(cm);A为土柱截面积(cm2);t为时间(s);H为水头高度(cm)。
为方便计算,取t= 10,得出:
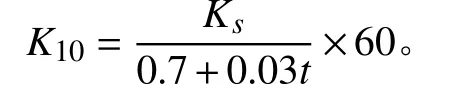
式中:K10为温度等于10 ℃时的饱和导水率(mm·min-1);Ks为t℃时的饱和导水率(mm·s-1);t为测量时的水温(℃)。
1.3 数据分析
试验数据采用Excel 2016 进行数据整理;采用SPSS 23.0 软件进行各指标的One-way ANOVA 统计分析,显著性水平为0.05;采用Origin 2018 作图。
2 结果与分析
2.1 植物群落结构
2.1.1 植被盖度和高度
随着退化程度的加剧,高寒草甸植被的总盖度和植物高度逐渐降低(图1)。从原生植被至重度退化阶段,植被群落总盖度和高度分别显著下降了58.5%和41.1% (P< 0.05)。
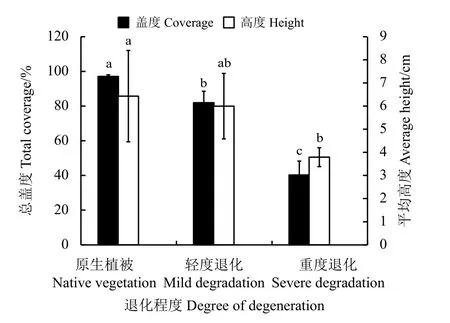
图1 三江源高寒草甸不同退化程度下植物平均高度及总盖度Figure 1 Average height and total coverage of alpine meadow under different degradation levels in the Three Rivers Source Region
2.1.2 物种丰富度与重要值
随着退化程度的加剧,高寒草甸植物群落物种丰富度明显降低,植物群落的物种组成也发生了显著变化(表1)。其中,原生植被样地植被群落共计36 种,分属16 科,其中,菊科7 种,莎草科4 种;轻度退化样地植被群落共25 种,分属14 科,其中,菊科3 种,莎草科3 种;重度退化样地植被群落共21 种,其中菊科2 种,莎草科2 种。原生植被样地植被群落主要由矮生嵩草、高山嵩草及麻花艽(Gentiana straminea)等构成,其重要值分别为8.99、8.35 和6.98;轻度退化样地植被群落主要由鹅绒委陵菜(Potentilla anserina)、细叶亚菊及高山嵩草等构成,其重要值分别为22.66、10.25 和8.86;重度退化样地植被群落主要由鹅绒委陵菜、西伯利亚蓼及附地菜等构成,其重要值分别为29.90、25.11 和10.33。随着高寒草甸退化程度的加剧,高山嵩草等物种消失,毒杂草成为优势种,导致草地生产功能减弱。

表1 玛沁高寒草甸不同退化程度下植物群落参数及重要值Table 1 Plant community parameters and important values under different degradation degrees in Maqin alpine meadow
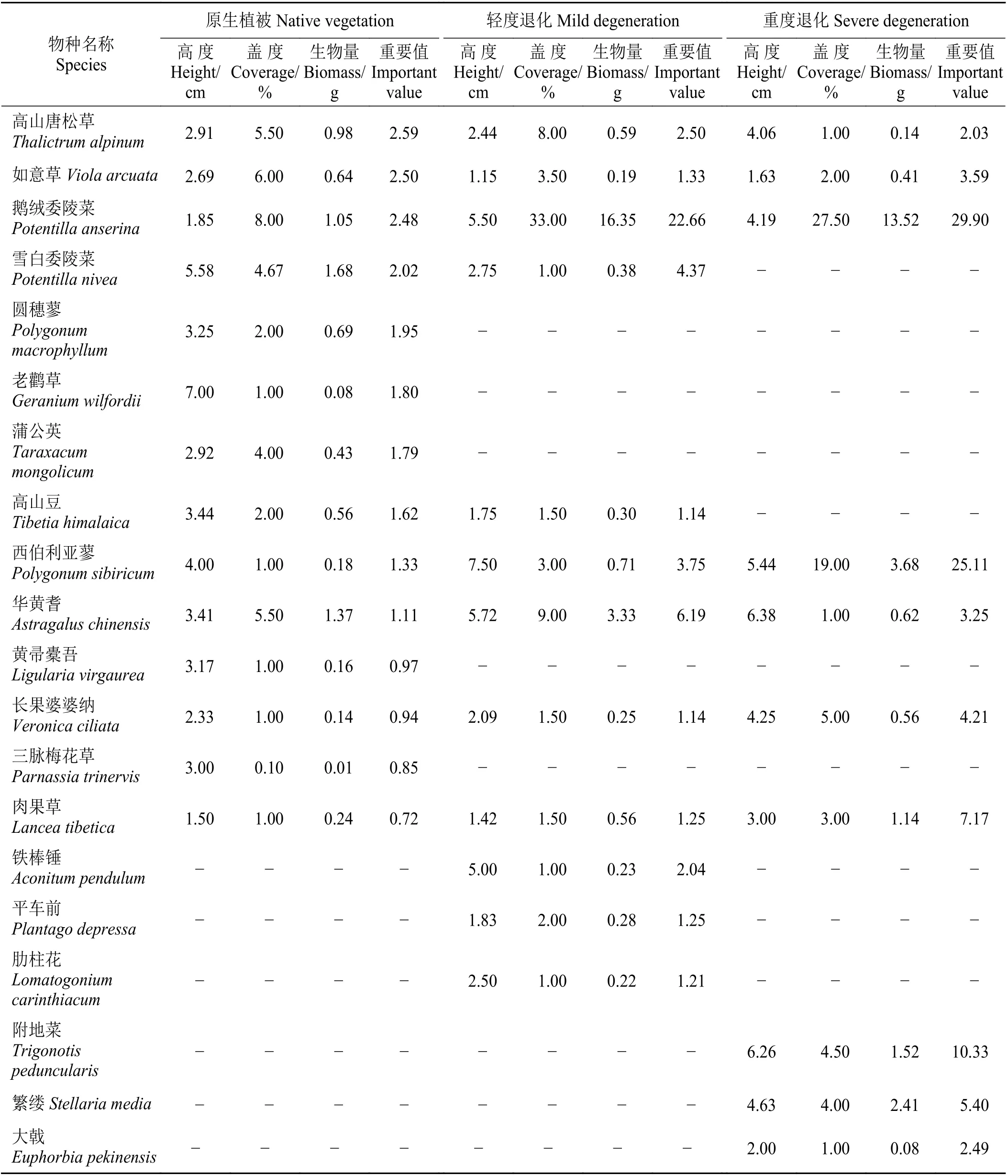
续表1Table 1 (Continued)
2.1.3 物种多样性
退化程度加剧降低了高寒草甸植被群落的物种多样性、丰富度、均匀度和优势度(图2)。重度退化样地的多样性指数较原生植被显著降低了25.6%(P< 0.05),均匀度指数和优势度指数分别下降了5.4%和3.9%,但均未达到显著水平(P> 0.05)。
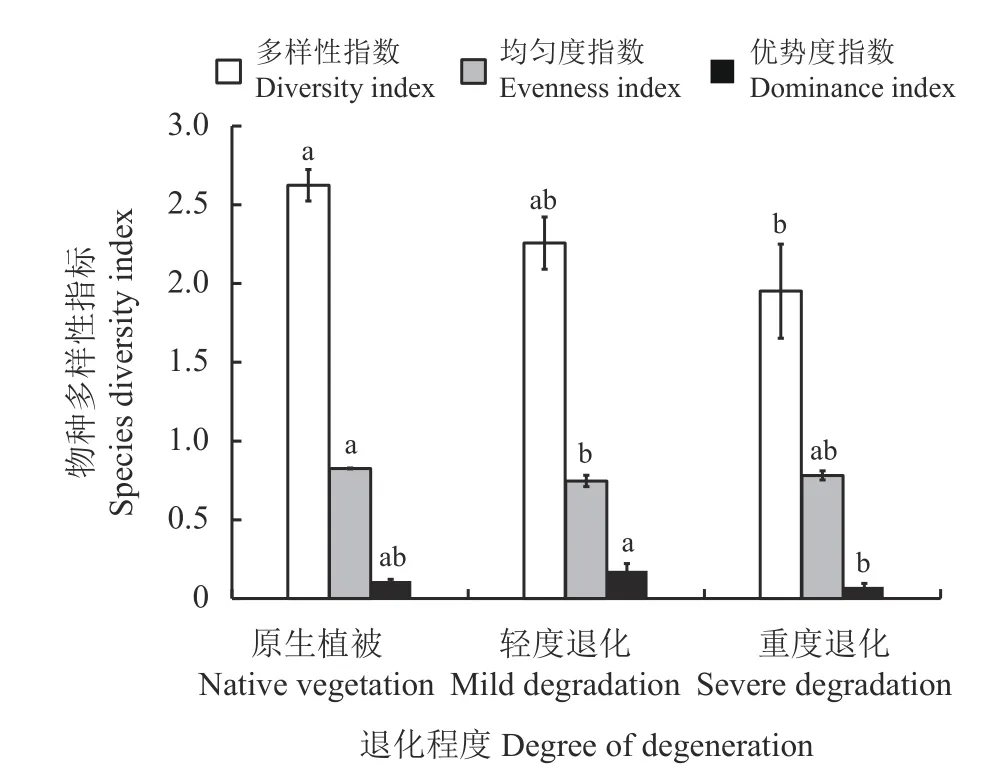
图2 玛沁高寒草甸不同退化程度下植物物种多样性指数Figure 2 Plant species diversity index under different degradation levels in Maqin alpine meadow
2.2 植物生物量
随着退化程度的加剧,高寒草甸的地上、地下及总生物量均呈逐渐减少趋势(图3A)。与原生植被样地相比,重度退化样地地上、地下及总生物量分别下降了49.8%、71.1%和70.0%,差异均达到显著水平(P< 0.05)。高寒草甸地下生物量主要集中在0 - 5 cm,占据地下总生物量的81% (图3B)。同时,不同土层地下生物量对退化程度的响应差异较大。其中,随着退化程度的加剧,表层(0 - 5 cm)地下生物量逐步降低,重度退化较原生植被显著下降了82.2% (P< 0.05);轻度退化显著提高了5 - 10 cm地下生物量(P< 0.05)。退化程度对高寒草甸深层(50 - 70 cm)地下生物量则无显著影响(P> 0.05)。
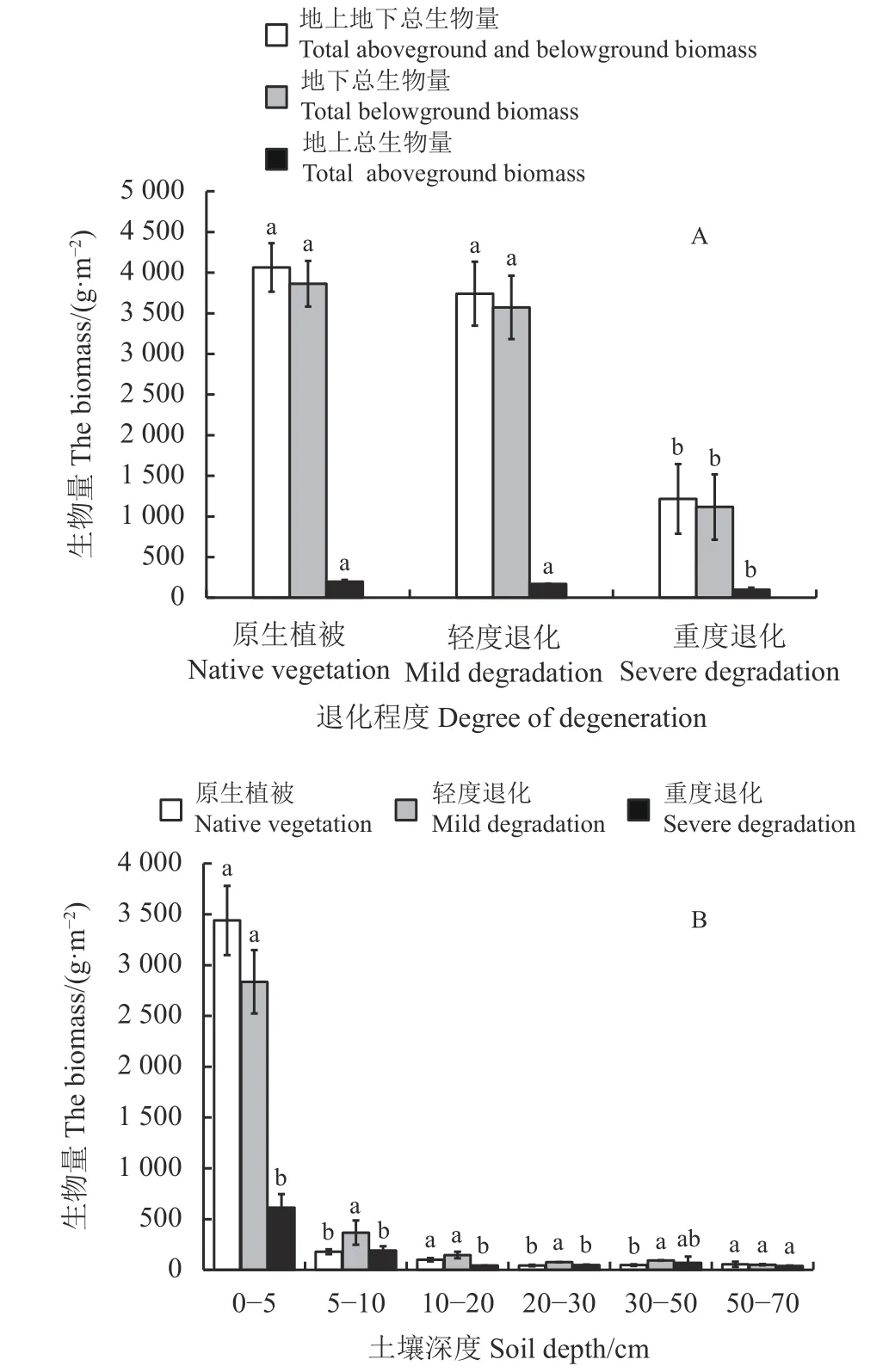
图3 玛沁高寒草甸不同退化程度下地上、地下总生物量(A)及不同土层的地下生物量(B)Figure 3 Total aboveground and belowground biomass (A)and belowground biomass of different soil layers (B) of Maqin alpine meadow at different degradation levels
2.3 土壤容重与持水能力
2.3.1 土壤容重
退化程度对高寒草甸不同土层土壤容重的影响差异明显,在0 - 20 cm 土层影响最大(表2)。随着退化程度加剧,0 - 5 cm 土层土壤容重显著增加(P<0.05),与原生植被相比,重度退化样地0 - 5 cm 土壤容重显著提高了41.0% (P< 0.05);5 - 10 cm 土壤容重随退化程度加剧呈先增加后降低趋势,差异未达到显著水平(P> 0.05);10 - 20 cm 土层土壤容重随退化程度加剧呈逐步降低趋势,重度退化样地10 -20 cm 土壤容重比原生植被显著降低了18.9% (P<0.05)。随着土壤深度的增加,退化程度对土壤容重的影响逐渐减弱,20 - 70 cm 土层的土壤容重无显著变化(P> 0.05)。

表2 玛沁高寒草甸不同退化程度下土壤容重及持水特征参数Table 2 Characteristics parameters of soil bulk density and water holding capacity at different degradation levels in Maqin alpine meadow
2.3.2 土壤饱和导水率与持水能力特征
退化程度对高寒草甸土壤饱和导水率与持水能力的影响主要集中在0 - 20 cm 土层,退化程度对二者的影响程度受土壤深度的影响(表2)。随着退化程度加剧,高寒草甸0 - 5 cm 土层土壤饱和导水率、饱和持水量、毛管持水量及田间持水量逐步降低,与原生植被相比,重度退化样地0 - 5 cm 土壤饱和导水率、饱和持水量、毛管持水量及田间持水量分别显著降低了84.4%、41.7%、41.4%和42.9% (P< 0.05)。随着退化程度加剧,5 - 10 cm 土层土壤饱和导水率、饱和持水量及毛管持水量无显著变化(P> 0.05),田间持水量呈先降低后稳定的趋势。10 - 20 cm 土层饱和持水量、毛管持水量及田间持水量随退化程度加剧呈逐步增加趋势,饱和导水率则呈先增高后降低趋势,重度退化样地10 - 20 cm 土壤饱和导水率、饱和持水量、毛管持水量及田间持水量较原生植被分别显著增加了100.0%、37.4%、35.8%和42.2%(P< 0.05)。随着土层的加深,退化程度对土壤饱和导水率与持水能力的影响逐渐减弱,50 - 70 cm 土层下四者均无显著变化(P> 0.05)。
3 讨论
3.1 退化程度对玛沁高寒草甸植被群落结构的影响
高寒草地生态系统逐渐退化主要表现在生物多样性的受损与生态系统循环的丧失。本研究结果表明,随着退化程度的加剧,高寒草甸植被盖度、植被群落高度及物种数量等特征均呈逐渐降低趋势;轻度和重度退化样地的多样性指数及均匀度指数低于原生植被样地,重度退化样地的优势度指数比轻度退化降低了57.9%,这与前人的研究结果基本一致[17-18]。出现这种结果的原因主要有:1)随着退化程度加剧,土壤肥力下降,持水能力降低,而相较杂类草,以禾本科、豆科和莎草科为主的优良牧草对水分和养分变化敏感,导致草地植被种间竞争格局改变,从而使退化样地的群落多样性指数与均匀性指数均低于原生植被样地[19]。2)过度放牧往往导致草地退化,过量的放牧家畜选择性地采食营养价值高、适口性好的优良牧草,导致草地植被优势种逐渐被毒杂草所取代,使其生活力与竞争能力减弱,最终导致群落物种多样性指数随着植被退化程度的加剧而降低[17]。
3.2 退化程度对玛沁高寒草甸生物量的影响
生物量是目前衡量草地植物生产力和植物生长发育的重要指标之一,也是生态系统健康评价的重要依据[20]。本研究结果显示,随着草地退化程度的加剧,地上、地下及总生物量均呈下降趋势,原生植被样地地上、地下生物量显著高于重度退化样地,这与前人在天祝高寒草甸的研究结果相似[21-22]。这主要是因为在高寒草甸退化演替过程中,植被物种多样性、植被盖度和高度逐步降低,导致地上植被部分和土壤中活根系的数量逐渐减少,土壤中的死根部分在微生物作用[23]与高原鼠兔(Ochotona curzoniae)等啮齿类动物活动影响下逐步降低[17,24-25]。与0 - 5 cm地下生物量变化趋势不同,5 - 30 cm 地下生物量在轻度退化下最高,这主要是因为:1)高寒草甸退化过程中,根系致密的禾本科和莎草科植物逐步被深根性的杂类草代替,造成轻度退化样地深层土壤地下生物量高于原生植被样地,且随着退化程度的进一步加剧,样地物种数显著降低,造成其地上、地下生物量的降低[26-27]。2)在轻度退化草地上存在一定强度的放牧活动,家畜的适度啃食去除了植物的顶端生长点与衰老组织,刺激了植被的补偿性生长;同时践踏促进了植物侧分生组织的生长;家畜排泄物为土壤供给养分,提高土壤肥力,为植物生长提供养分,因此轻度退化样地植物出现 “超补偿性”生长现象[28-29]。
3.3 退化程度对玛沁高寒草甸水源涵养功能的影响
土壤饱和导水率、土壤饱和持水量、毛管持水量、田间持水量是衡量土壤导水和持水能力的重要参数,用以衡量土壤通透性和贮水能力。同时,容重可表征土壤结构状况与土壤的松紧程度,可用于评价土壤的水源涵养功能[23]。本研究表明,高寒草甸0 -5 cm 土层土壤容重随退化程度的加剧逐步增大,饱和持水量、毛管持水量及田间持水量呈下降趋势,这与岳广阳等[30]和徐翠等[31]的研究结果一致,因为高寒草甸植被的根系主要集中在0 - 10 cm 土层,莎草科植物拥有庞大的地下根系,使得高寒草甸形成相对稳定的草毡层,随着退化程度的加剧,高寒草甸草毡层消失,植被物种数、盖度、生物量逐步降低,植被群落结构发生变化,土壤有机质和腐殖质含量逐渐减少,土壤逐渐贫瘠化,导致植被根系减少,土壤孔隙度和水分渗透能力减弱[8,11,32-33],土壤容重逐步增加,土壤持水能力及饱和导水率显著降低。这也说明草毡层(浅层土壤)是高寒草甸保持该区域水源涵养功能及通透性的关键所在。值得注意的是,本研究中随着退化程度的加剧,高寒草甸10 - 20 cm 土层出现容重降低、土壤持水量等指标增加的现象,这可能与高寒草甸退化过程中深根性的杂类草逐渐替代须根系的禾本科和莎草科植物有关[33]。
4 结论
1) 随着退化程度的加剧,高寒草甸植被群落的总盖度、平均高度、物种多样性、物种数、丰富度、多样性指数及均匀度指数均逐步下降,其中重度退化样地的多样性指数、均匀度指数和优势度指数较原生植被分别降低了25.6%、5.4%和33.9%。
2) 地上、地下生物量均随退化程度加剧而减少,与原生植被样地相比,重度退化样地地上、地下及总生物量分别显著下降了49.8%、71.1%和70.0%,同时研究发现轻度退化的高寒草甸出现“超补偿性”生长现象。
3) 退化程度对土壤持水能力的影响主要集中在0 - 5 cm,随着高寒草甸退化程度加剧,莎草物种逐渐被杂类草取代,土壤持水能力大幅度降低。在0 - 5 cm 土层土壤饱和导水率、饱和持水量、田间持水量、毛管持水量均逐渐降低。
