基于组合物种分布模型的黄河源区鹅绒委陵菜适宜生境及其对气候变化的响应
2022-03-21张欣雨朱泽群袁雅欣姬文翔宋岑雨陆洪伟赵茜蕾卞庆瑶孙子墨宜树华
张欣雨,朱泽群,袁雅欣,姬文翔,宋岑雨,陆洪伟,赵茜蕾,卞庆瑶,孙子墨,宜树华,孙 义
(南通大学脆弱生态环境研究所 / 地理科学学院, 江苏 南通 226007)
全球气候变化引起自然地理环境格局的变化是影响物种地理分布的主要环境因素[1]。高海拔地区对全球气候变化更为敏感,黄河源区位于青藏高原的东北边缘,是我国重要的水源涵养地和生态安全屏障[2],也是最主要的畜牧业产业基地之一。高寒草地约占源区总面积的80%,是源区主要的自然资源和生态环境载体[3]。自身脆弱的生态环境、气候变化和不合理的人类活动是黄河源区植物群落结构持续退化的主要原因[4]。预测高寒草地植物物种的生境分布及其对气候变化的响应对草地的合理开发与生物多样性的保护至关重要。
鹅绒委陵菜(Potentilla anserina)是蔷薇科(Rosaceae)委陵菜属(Potentilla)多年生草本植物,主要生长于海拔500~4 100 m 的河岸、路边、山坡草地及草甸,是青藏高原主要的野生经济植物之一[5]。鹅绒委陵菜富含维生素、蛋白质、脂肪、淀粉、氨基酸以及各类矿质元素等[6],块根可药食两用;同时,也是国家级畜禽遗传保护品种蕨麻猪的主要饲料,对当地的农牧业经济发展和特种畜禽种质资源保护具有重要意义[7]。鹅绒委陵菜具有抗氧化及抗衰老、保护肝损伤、保护心肌细胞、增强免疫力、抑菌等药理作用[8],具备良好的药用价值。鹅绒委陵菜可进行有性和无性繁殖,具有广泛的生态适应性[9],耐瘠薄、耐寒、耐旱、抗盐碱、根系发达,有较强的蓄水保墒固沙能力[10];也被视作草地重度退化的优势指示植物[11],对高寒草地生态系统的管理具有不可忽视的经济和生态价值。虽然鹅绒委陵菜分布广泛,但其根系只有在海拔较高地区才会形成肥厚并具滋补作用的块根[12]。因此,研究鹅绒委陵菜在黄河源区的空间分布对其合理的开发与利用具有重要作用。
物种分布模型(species distribution models, SDMs)是利用物种的已知分布数据和相关环境变量推算物种的生态需求,将其运算结果投射至不同的空间和时间以预测物种的潜在分布[13],能很好地揭示物种存在记录和空间预测因子之间的复杂关系[14]。近年来物种分布模型被广泛应用于入侵生物学、保护生物学、进化生物学以及物种关键生境的识别[15]和全球气候变化对物种分布的影响等研究领域[16]。BIOMOD (BIOdiversity MODeling)是基于R 语言的物种分布集成预测的计算机平台,包含10 个不同的物种分布模型[17]。由于不同物种之间生态位和生境特征的差异,研究者们通常针对不同的应用场景而选择不同的物种分布模型。例如,使用便捷的最大熵模型(maximum entropy model, MaxEnt)在该领域得到了广泛的应用。然而,Tessarolo 等[18]认为仅靠单一的物种分布模型往往难以准确地确定物种的适宜生境,Aguirre-Gutiérrez 等[19]的研究表明多个模型的组合比单个模型预测精确度高。因此,以多模型组合为特征的BIOMOD 可实现择优构建组合模型,能够更好地模拟物种的空间分布并推断其主要影响因子。物种基础分布数据是准确模拟和预估物种空间分布特征的基础。传统地面调查方法由于效率低下、标准不统一、人力成本高和观测范围小等限制因素而难以开展大区域调查[20]。尤其在特殊环境研究区(如青藏高原高寒缺氧、植物生长季短暂、生境脆弱),传统观测方法难以在有限的时间内完成大范围的样本采集工作,而且可能对植物生境造成破坏[21]。此外,目前普遍使用的网络标本馆的物种分布数据通常缺乏时效性及准确全面的地理定位信息[22],属被动获取数据方式,数据量偏少、代表性受限。更为关键的是该类数据集无法提供物种不存在的样本,从而对模拟结果的准确性产生直接影响[23]。随着无人机(unmanned aerial vehicle,UAV)技术日益成熟[22],其所具备的高时效性、高分辨率、低成本、标准统一等优点为克服取样环境限制、获取高时效性数据提供了新的方法。随着无人机路径规划和信息提取技术的不断完善,如Yi[24]自主开发的无人机路径规划和信息提取系统(fragmentation monitoring and analysis with aerial photography, FragMAP)为植被覆盖度、高原鼠兔总洞口和种群密度、地上生物量、砂砾石分布和植物种类等生态学领域研究提供大量、标准统一的数据,为鹅绒委陵菜基础空间分布格局数据的获取和分析奠定了基础。
本研究采用无人机获取黄河源区鹅绒委陵菜的基础分布数据,利用BIOMOD 物种分布集成平台建立组合物种分布模型,结合气候、地形和土壤等环境数据,刻画鹅绒委陵菜在黄河源区的潜在分布,揭示影响其空间分布的主要影响因子,并预测全球气候变化背景下其空间分布格局的变化。以期为高寒草地经济作物的合理管理与利用、特色农业经济发展以及黄河源区生态恢复和高寒地区的生态建设提供一定的理论和实践基础。
1 材料与方法
1.1 研究区概况
黄河源区位于青藏高原东北部边缘(95°50′~103°30′ E,32°30′~36°10′ N),包括青海、四川和甘肃三省的6 个州、18 个县。该区平均海拔在4 000 m以上,地形地貌复杂多样,地势西高东低,属于典型的高原大陆性气候,多年平均气温为-3~3 ℃,自西北向东南递增,降水多集中于夏秋季节,多年平均降水量在200~700 mm,由东南向西北递减[25]。源区内河源区高寒草地呈现出显著退化趋势,具体表现为覆盖度降低、破碎化与干旱化加剧[26],优势植物以莎草科(Sedge)和禾本科(Gramineae)为主,优势种有矮嵩草(Kobresia humilis)、垂穗披碱草(Elymus nutans)和青藏苔草(Carex siderosticta)等[27],土壤以高寒草甸土和高寒冻土为主[28]。
1.2 鹅绒委陵菜基础分布数据获取
鹅绒委陵菜基础分布数据由2018-2020 年进行的无人机调查获得(图1)。使用无人机型号为大疆创新科技有限公司(DJ-Innovations, DJI)生产的Phantom 3 Professional 和Mavic Pro。由南通大学脆弱生态环境研究所自主开发的FragMAP 系统设置工作点及航线[20],控制无人机自动按照设定航线(Belt mode)飞行并定点垂直向下拍照(图2a),飞行高度设置为2 m。每条航线覆盖范围为40 m × 40 m(本研究中每条航线视为一个样本采集点),每个样本采集点包含16 张航拍照片,单张航拍照片的覆盖地面范围约为9.1 m2(3.5 m × 2.6 m) (图2b),分辨率为0.55 mm。借助FragMAP 系统的Proposal Classifier软件目视辨别获取鹅绒委陵菜存在与否的信息并记录(即0,1 形式数据) (图3)。在研究区共设置了202个样本采集点(包括202 条航线 × 16 张航拍照片 =3 232 张航拍照片)。因此,尽管放牧是影响高寒草地群落结构的重要因素,但本研究布设的202 个拍摄点(工作点)以及每个工作点所含的16 幅航拍照片跨越了不同放牧强度,即基于此数据获取的结果是涵盖了不同放牧梯度下的综合结果。在数据提取时,只要鹅绒委陵菜出现于任意一张航拍照片,就记录为该样本采集点存在鹅绒委陵菜(即“1”),该采集点的地理位置记为出现该物种最多的航拍照片的经纬度;如果某一采集点的所有航拍照片都没有鹅绒委陵菜,则记录为不存在(即“0”),其地理位置记为该工作点下任意一张照片的经纬度。在黄河源区内共获得96 条存在记录和106 条不存在记录作为基础分布数据。

图1 黄河源区鹅绒委陵菜样本采集点分布Figure 1 Location of sampling points for Potentilla anserina in the source region of the Yellow River
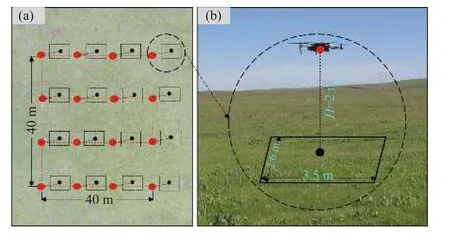
图2 基于无人机路径规划和信息提取系统的Belt 航线设置示意图Figure 2 Belt mode of the FragMAP system
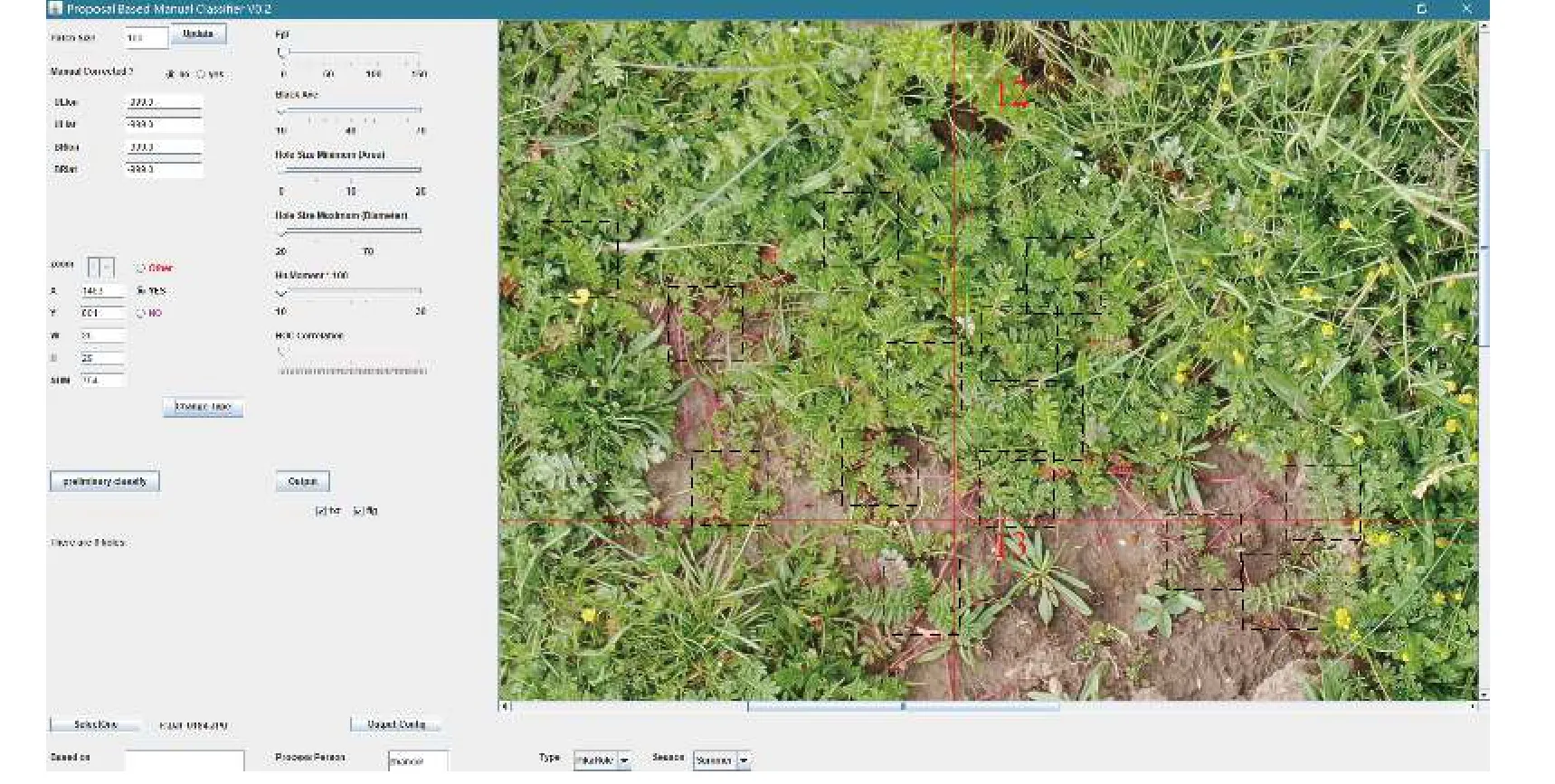
图3 基于无人机的协同航拍和分析系统的Proposal-Classifier 软件提取鹅绒委陵菜信息Figure 3 Extraction of information relating to Potentilla anserina from aerial photographs obtained using the FragMAP system in conjunction with Proposal Classifier software
1.3 环境数据
1.3.1 气候数据
本研究从WorldClim (Global Climate Data)下载得到30 s 分辨率的气候数据(www.worldclim.org/cmip5_30s),包括当前气候数据(1970-2000 年)和未来气候数据。未来气候数据为BCC-CSM1.1[29](Beijing Climate Center Climate System Model version 1.1)气候模式构建在RCP 4.5 和RCP 8.5 两种代表性路径浓度(representative concentration pathways, RCPs)下2050年(2041-2060 年间的平均)和2070 年(2061-2080年间的平均)的气候数据。代表性浓度路径是政府间气候变化专门委员会在2014 年第五次评估报告中采用的4 种温室气体浓度轨迹[30],包括RCP 2.6、RCP 4.5、RCP 6.0 和RCP 8.5,是以2100 年相对于工业化前的辐射强迫的增加值命名(分别代表增加2.6、4.5、6.0 和8.5 W·m-2)[31]。该数据集包含了气温和降水两类气候因子,共有19 个栅格图层。
1.3.2 地形数据
数字高程数据(digital elevation model,DEM)来自美国地质勘探局(www.usgs.gov),为SRTM 90 m DEM 数据。基于DEM 数据,使用QGIS Desktop 提取坡度和坡向,共获得海拔、坡度、坡向3 个栅格图层。
1.3.3 土壤数据
采用Soil Grids (www.soilgrids.org)的土层厚度,0.3 - 0.6 m 土层有机碳储量,0.3 m 深度处土壤容重、黏土含量、粗屑体容积、淤泥含量、含沙量和土壤pH,共获得分辨率为1 km 的8 个栅格图层。
1.3.4 环境数据预处理
为了降低环境变量在数据集上的多重共线性,计算了每一对变量之间的Pearson 相关系数。当两个环境变量之间的相关系数高度相关(|r| > 0.8)时,其中一个将被剔除。经过筛选,保留了年平均气温、气温年较差、年降水量、最干月降水量、海拔、坡向、坡度、0.3 - 0.6 m 土层有机碳储量和0.3 m 深度处土壤粗屑体容积、淤泥含量、pH。
1.4 构建模型和模型评价
BIOMOD 共提供了10 种物种分布模型,可以通过不同指标计算模型精度来评估不同模型在研究区的适用性,从而筛选出最优的模型。包括广义线性模型(generalized linear models, GLMs)[32]、广义增强回归模型(generalized boosted regression models, GBMs)[33]、广义加性模型(generalized additive model, GAM)[34]、分类树分析(classification tree analysis, CTA)[35]、人工神经网络(artificial neural networks, ANNs)[36]、表面分布区分室模型(surface range envelope, SRE)[37]、弹性判别分析(flexible discriminant analysis, FDA)[38]、多元自适应回归样条(multivariate adaptive regression splines, MARS)[39]、随机森林(random forest,RF)[40]和最大熵模型(maximum entropy model, MaxEnt)[41]。
将输入的样本随机划分为两个子集,其中一个子集包含全部样本的70%,用于建模;另一个包含30%,用于检验。在模拟的过程中,biomod2 包可以计算出每个环境因子的重要性,重要性值越高表明该环境因子对物种分布的影响越大。利用真实技巧统计值(true skill statistics,TSS)和受试者工作特征曲线(receiver operator characteristic curve,ROC)下面积(area under the ROC curve,AUC)两种统计学评价指标对模型预测结果进行评估。
TSS 的计算方法为TSS=Sensitivity+Specificity-1 =TPR-FPR。其中Sensitivity为灵敏度,Specificity为特异度,TPR(true positive rate)为真阳性率,FPR(false positive rate)为假阳性率。由Allouche 和Kadmon[42]于2006 年提出,它既继承了Kappa 统计值的优点,且克服了传统模型评价方法的不足,即Kappa 统计值对物种发生率呈单峰曲线响应形式的弱点。TSS的值域为[0, 1],值越接近于1 表示真阳性率与假阳性率的差值越大,模型预测效果越好。
ROC 曲线是以假阳性率(false positive rate)为横轴,真阳性率(true positive rate)为纵轴,根据接受者在不同条件下,用不同判别标准得出的结果所绘制的曲线。AUC 为ROC 曲线与坐标轴所围成的面积[43]。与Kappa 统计值和真实技巧统计值不同的是,AUC的值因不受诊断阈值的影响,且对物种发生分布率(prevalence)不敏感,可对两个诊断试验的准确度进行综合比较,目前被公认为是最佳的生态位预测结果评价指标之一[44]。AUC 的值域为[0.5, 1],值越接近于1 表示模型拟合优度越高,模型预测效果越好;而越趋近于0.5 则表示模型预测效果越差,接近于随机估计。
参考Guo 等[45]的方法,根据模型精度,择优构建组合模型。首先,将模拟结果的范围从[0, 1 000]调整到[0, 1]。其次,通过加权平均法对所选模型进行集成。模型权重为单个模型AUC 值与所有模型AUC值之和的比值。计算公式如下:

式中:Wj表示第j个模型的权重,rj表示第j个模型的AUC 值,h表示集合建模中的模型个数,在本研究中为8。最后,将单个模型的归一化结果依次乘以相应的权重后求和,得到组合模型下研究区内鹅绒委陵菜分布概率。计算公式如下:
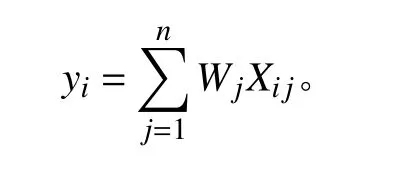
式中:yi表示鹅绒委陵菜在第i个栅格的分布概率,Wj表示第j个模型的权重,Xij表示第i个栅格在第j个模型中的值。yi的取值范围为[0, 1],yi越接近1,则该栅格所代表的地理空间越适合于鹅绒委陵菜的生长。
2 结果
2.1 模型精度评估
基于黄河源区鹅绒委陵菜基础分布数据和环境数据,在BIOMOD 所含的GLM、GBM、CTA、ANN、SRE、FDA、MARS、RF 和MaxEnt 9 种模型下(GAM由于获取参数困难而运行失败)重复运行50 次得到TSS 和AUC 统计结果(图4)。RF 的TSS 和AUC 的平均值分别为0.776 和0.926,表现出较高的精确度和较好的稳定性;GBM 的整体表现次之,其TSS 和AUC 平均值分别为0.771 和0.925。SRE 模型表现较差,其TSS 和AUC 平均值分别为0.422 和0.711。
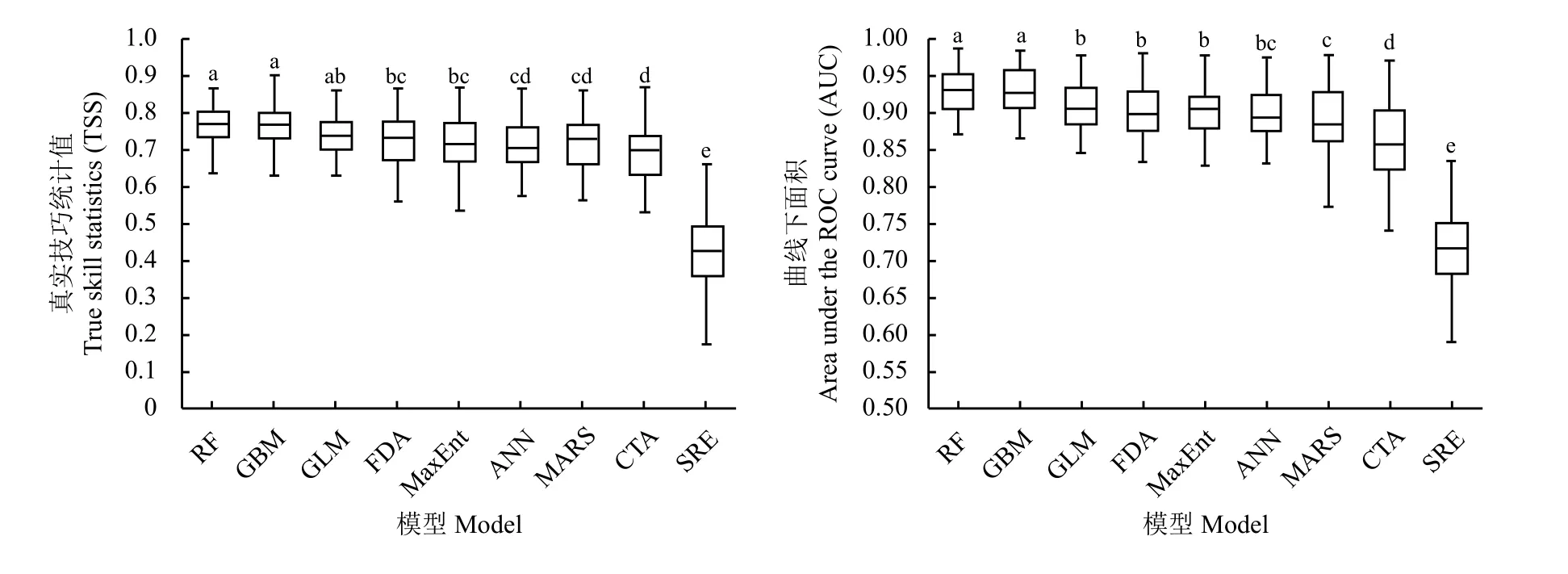
图4 基于TSS 和AUC 的9 种物种分布模型统计结果Figure 4 Results of statistical analyses of the nine species distribution models based on true skill statistics and area under the ROC curve analyses
2.2 环境因子重要性及其对物种分布的影响
在参与建模的11 个环境数据中,年降水量和海拔的重要性最高,这两种环境变量重要性之和的权重占70%以上,因此年降水量和海拔为鹅绒委陵菜空间分布的主要影响因子,年平均气温和土壤pH的重要性次之。鹅绒委陵菜的分布受土壤淤泥含量、坡度等环境因子影响较小(表1)。
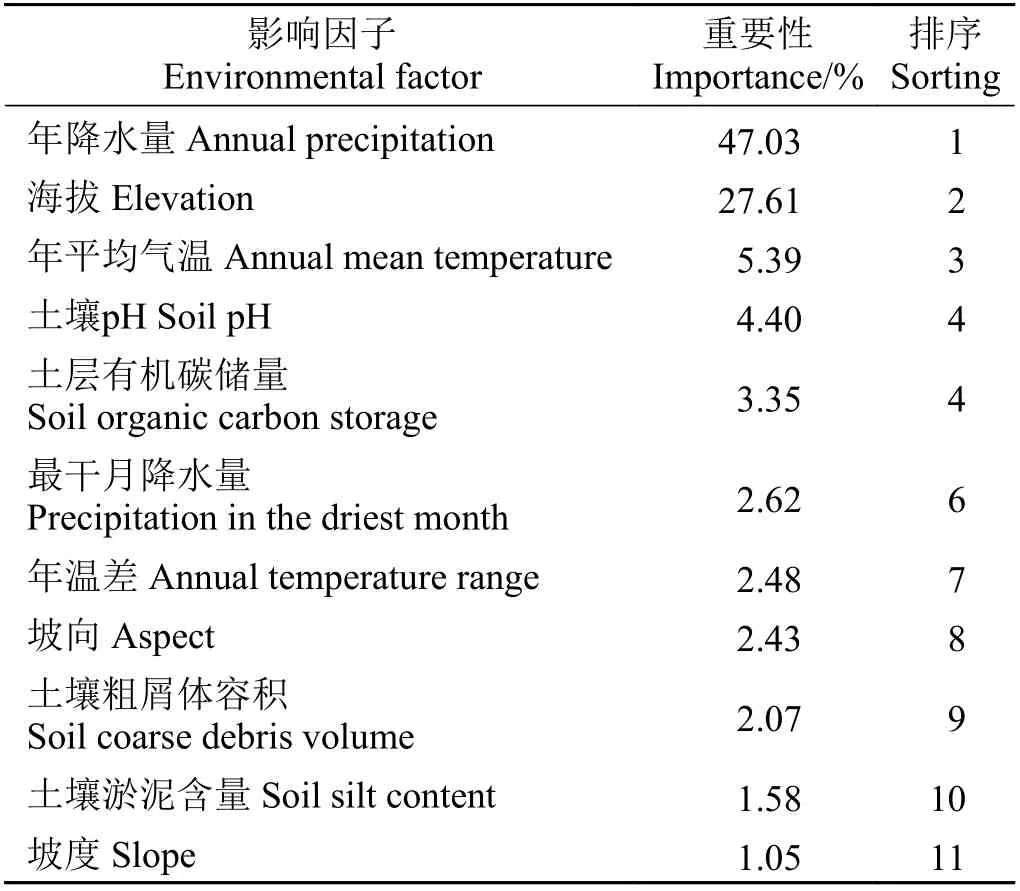
表1 影响因子重要性排序结果Table 1 Importance ranking of factors influencing the distribution of Potentilla anserina
鉴于鹅绒委陵菜适宜生境与环境变量之间并非特定的线性或单调关系,采用广义加性模型算法绘制了基于组合模型预测结果的环境变量响应曲线(图5)。结果表明,鹅绒委陵菜更适合生长于年降水量大于500 mm,海拔为3 400~4 100 m,年平均气温为-3~3 ℃,土壤pH 为5.8~7.3。

图5 生境适宜性与环境变量拟合曲线Figure 5 Fitting of curves for habitat suitability and environmental variables
2.3 鹅绒委陵菜潜在分布格局
根据模拟结果,将生境适宜性划分为4 个概率等级:0~0.25 为低适宜生境,0.25~0.50 为中适宜生境,0.50~0.75 为高适宜生境,0.75~1.00 为极适宜生境。根据模型精度评价结果,除了表现较差的SRE 模型和运行失败的GAM 模型,其余8 种模型的准确率均较好(TSS > 0.65,AUC > 0.85)。其中,ANN 模型能较好地模拟鹅绒委陵菜的极适宜生境,预测结果为其高度集中于黄河源区中部区域,其余7 种模型的预测结果大体一致,即鹅绒委陵菜主要分布于黄河源区的中部和东南部(图6)。

图6 基于单一模型的鹅绒委陵菜在黄河源区的潜在分布Figure 6 Potential distribution of Potentilla anserina in the source region of the Yellow River based on different models
将这8 种模型按照AUC 权重构建组合模型。根据组合模型结果,鹅绒委陵菜的适宜生境主要集中于黄河源区的中部和东南部,分布概率较高的县域主要有泽库县、河南蒙古族自治县、玛曲县、久治县、若尔盖县、阿坝县、红原县、甘德县、达日县、玛沁县东部、同德县南部,其他区域则分布概率较小(图7)。鹅绒委陵菜在黄河源区的平均分布概率为34.25%,其中低适宜生境面积占比47.96%,中适宜生境面积占比15.08%,高适宜生境面积占比33.68%,极适宜生境面积占比3.29% (表2)。

图7 基于组合模型的鹅绒委陵菜在黄河源区的潜在分布Figure 7 Potential distribution of Potentilla anserina in the source region of the Yellow River based on ensemble models
2.4 气候变化对鹅绒委陵菜空间分布的影响
与潜在分布的模拟一致,选择GLM、GBM、CTA、ANN、FDA、MARS、RF 共8 种模型建立组合分布模型预测鹅绒委陵菜的未来分布情况(图8)。结果显示,鹅绒委陵菜主要分布区域仍在黄河源区中部及东南部,总体分布概率呈上升趋势,从现在到2070 年,极适宜生境面积将会先增加后降低。
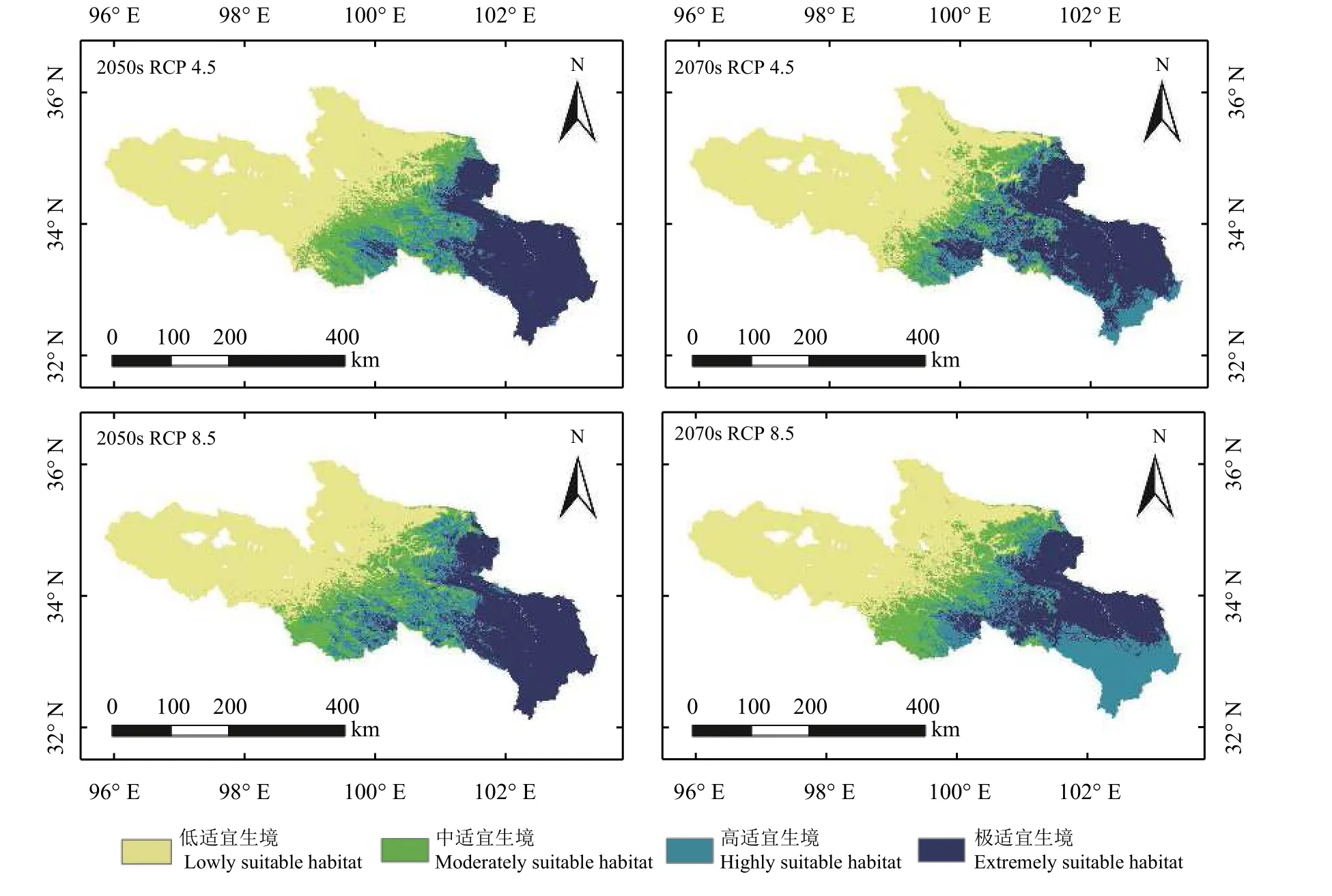
图8 基于组合模型的鹅绒委陵菜在黄河源区的未来分布Figure 8 Predicted future distribution of Potentilla anserina in the source region of the Yellow River based on ensemble models
在RCP 4.5 情景下,2050 年黄河源区鹅绒委陵菜的平均分布概率为34.17%,低适宜生境面积占比52.32%,中适宜生境面积占比14.29%,高适宜生境面积占比12.80%,极适宜生境面积占比20.60%;2070年黄河源区鹅绒委陵菜的平均分布概率为35.87%,低适宜生境面积占比50.25%,中适宜生境面积占比10.60%,高适宜生境面积占比23.68%,极适宜生境面积占比15.48% (表2)。
在RCP 8.5 情景下,2050 年黄河源区鹅绒委陵菜的平均分布概率为36.35%,低适宜生境面积占比47.94%,中适宜生境面积占比15.77%,高适宜生境面积占比14.79%,极适宜生境面积占比21.50%;2070年黄河源区鹅绒委陵菜的平均分布概率为36.14%,低适宜生境面积占比49.28%,中适宜生境面积占比12.82%,高适宜生境面积占比23.33%,极适宜生境面积占比14.56% (表2)。
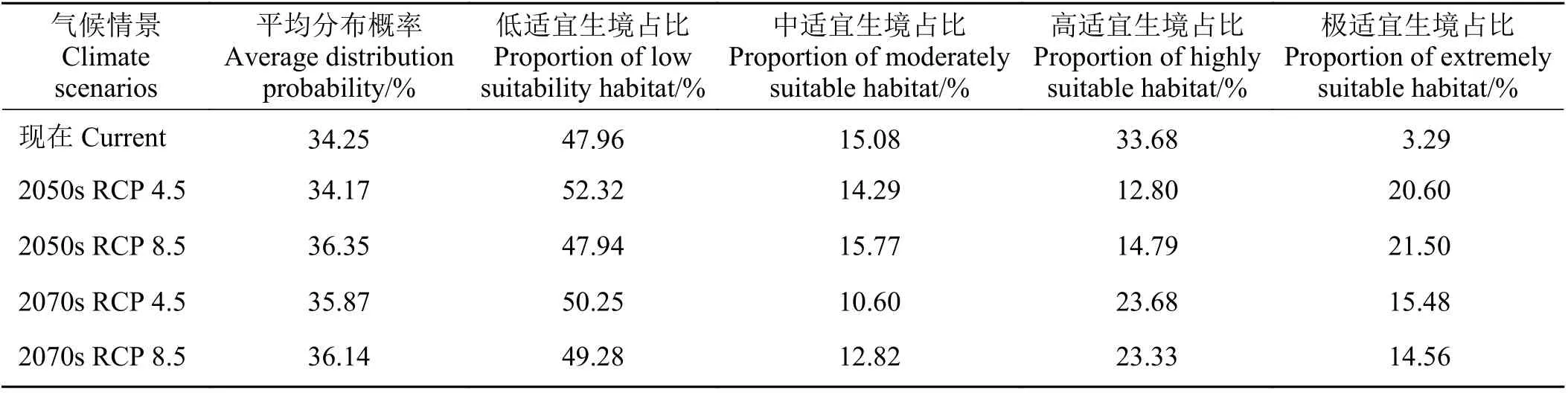
表2 鹅绒委陵菜平均分布概率及适宜生境占比Table 2 Average distribution probability and proportions of suitable Potentilla anserina habitat
3 讨论
3.1 鹅绒委陵菜潜在分布及主要影响因子
不同模型的模拟过程及算法存在差异性,其模拟结果亦存在不确定性[46]。本研究利用BIOMOD 物种分布集成平台对单个模型进行评估后加权构建组合模型,可有效降低单一模型模拟引起的偏差[47]。为了防止过拟合,本研究采取Pearson 相关分析法剔除相关性较强的部分环境因子,最终选取4 种气候因子、3 种地形因子和4 种土壤因子参与建模。结果表明,鹅绒委陵菜的适宜生境主要集中于黄河源区的中部和东南部,影响其空间分布的主要因子是年降水量和海拔,即鹅绒委陵菜的适宜生境为年降水量大于500 mm 的区域(图5),该结果与在缺水条件下,其幼苗高度、单株生物量和匍匐茎数量均呈下降趋势的研究结果相吻合[48]。毛振华[49]发现2010 年6 月 - 9 月降水量248.4 mm 的条件下,鹅绒委陵菜仍需灌溉总量为110.7 mm,也佐证了本研究的结果。海拔梯度是包含温度、湿度和光照条件(太阳辐射)等因素的综合环境因子,是影响物种分布规律的重要因素[50。本研究显示,3 400~4 100 m 是鹅绒委陵菜适宜生长的海拔范围(图5),其与张彦芬等[18]揭示的在青海省广泛分布于海拔1 800~4 000 m 的区域以及《中国植物志》[51]中描述其生长于海拔500~4 100 m 的上限分布区基本一致。本研究的区域选择黄河源区,海拔在3 000 m 以上,因而鹅绒委陵菜适宜生长区的下限仅限于研究区。本研究结果说明海拔对鹅绒委陵菜空间分布的影响明显,而且本研究方法能够较为准确地揭示其作用强度(表1 和图5)。此外,年平均气温和土壤pH 也在一定程度上制约鹅绒委陵菜的空间分布,年平均气温为-3~3 ℃的区域更适合鹅绒委陵菜生长。余青兰和李军乔[52]曾提出鹅绒委陵菜属于典型的低温植物,温度降到11 ℃左右时其生物量才达到高峰,且低温可刺激其块根的膨大;任飞等[53]通过模拟增温试验发现高温低湿的环境不利于鹅绒委陵菜可溶性糖含量的积累。刘贺贺等[54]的研究表明,鹅绒委陵菜适宜的土壤环境为中性、近中性,与本研究认为的鹅绒委陵菜较适宜在中性微酸的环境中生长一致。
3.2 全球气候变化对鹅绒委陵菜分布的影响
年降水量是影响鹅绒委陵菜空间分布的主导因子(图5),未来降水模式的变化将对其分布产生主要的影响。全球变暖将导致中国西部,特别是青藏高原降水增多[55],未来的降水模式将会有利于鹅绒委陵菜的生长。从鹅绒委陵菜的整体生态适宜性来看,在当前气候情景下鹅绒委陵菜极适宜生境的面积占比较低,而在两种未来气候情景下,极适宜生境的分布面积都表现出了先增加后降低的趋势,说明鹅绒委陵菜将会较好地适应短期内的气候变化,这与全球变暖促进植被生长的结论相一致[56]。然而,温度的持续上升将会对植被产生负面影响[57],本研究也证实了随着全球变暖趋势的不断加强,鹅绒委陵菜的极适宜生境面积可能将会产生缩减的现象。在当前气候情景下极适宜生境空间分布范围主要在黄河源区的中部而且面积占比较低,而在RCP 4.5 和RCP 8.5 情景中,鹅绒委陵菜的空间分布都表现为在2050 年向南迁移,在2070 年又向北回迁的趋势,其原因可能是南部地区地形所导致的生态复杂性影响着鹅绒委陵菜的迁移方向和速度[58]。从大空间尺度来看,随着全球气候变化,鹅绒委陵菜分布始终保持在黄河源区的中部和东南部地区。作为重要的经济作物,鹅绒委陵菜未来的生长条件可能将对未来的相关产业发展产生重大影响[59]。
3.3 无人机技术为物种分布模型提供基础数据
完整的基础分布数据是准确模拟物种时空分布的前提和保障[60]。传统植物空间分布、群落大小以及演替趋势的研究中,所使用的物种分布数据主要来源于地面调查或网络标本馆[61]。传统地面调查方法因人力成本高和覆盖范围受限等因素而难以在大尺度、尤其是在恶劣或生境脆弱的研究区获取准确、全面的基础分布数据;网络标本馆的物种分布数据则通常缺乏时效性和完整性(例如缺乏精确的地理定位信息和物种缺失信息)[22]。从实践应用角度来说,无人机技术结合FragMAP 操作和分析系统在物种基础分布调查中具有效率高、成本低、适合大范围调查、无破坏性和受地形限制小等特点[62]。从方法论角度而言,基于FragMAP 系统的数据获取过程被有机拆分为野外取样和室内信息提取两部分[63],能够很好地提高样品(航拍照片)采集效率、减少人为对样本采集区的环境破坏,同时保证了数据的准确性和统一性,为物种时空分布研究提供了新的思路和方法。
3.4 存在问题及研究展望
本研究结果虽可为揭示黄河源区鹅绒委陵菜的分布及其对气候变化的响应提供一定依据,但仍存在一些局限性。首先,本研究基于无人机实测的鹅绒委陵菜分布数据集作为初始数据,但采集的样本多沿公路布局,空间分布不均匀,可能对模拟结果产生一定的影响。在后续物种样本采集时应充分考虑所选采集区的随机性、代表性和完整性。其次,本研究在进行环境变量的选择时,偏重于鹅绒委陵菜生长所需的气候、地形和土壤等环境因子,而没有涉及到其自身组织结构特征和人为干扰因素,例如物种的迁移能力、物种间的相互作用、放牧活动以及土地利用变化等[62],这些因素均对对鹅绒委陵菜的时空分布产生潜在的影响。但是,因为目前尚没有被认可的适宜应用于BIOMOD 模型的量化数据集,因此本研究中未予充分考虑。在后续的研究中,有关植被自身组织结构特征和人为干扰因素数据集的开发应受到充分重视,进而提升物种时空分布模拟的准确性。此外,FragMAP 系统具备野外高效获取高分辨率航拍照片(即样本)的优势,但是室内信息提取过程还是沿用人工目视识别的方法,效率低下、对研究者基本技能要求较高。因此,后续研究中,结合卷积神经网络(convolution neural network)等技术的图像自动识别的引入将进一步提升物种基础分布数据获取的效率和准确率。
4 结论
本研究利用组合物种分布模型模拟预测了鹅绒委陵菜在黄河源区的潜在空间分布格局,并探讨了主要的影响因子及其适宜生境对气候变化的响应。结果表明单个物种分布模型中,RF 的表现最优(TSS和AUC 的平均值分别为0.776 和0.926),而组合模型可以降低预测的不确定性,较好地模拟黄河源区鹅绒委陵菜的空间分布格局;在黄河源区,影响鹅绒委陵菜空间分布的主要环境因子为年降水量和海拔,鹅绒委陵菜在黄河源区的极适宜生境面积占比为3.29%,潜在分布区主要集中在中部和东南部;在未来气候情景下,黄河源区的鹅绒委陵菜极适宜生境面积呈先增加后降低、空间分布呈先向南后向北回扩散的趋势;放牧活动等人为因素可能对其时空分布产生直接影响,在后续的研究中应加强因子的量化工作进一步提升物种时空分布模拟结果的准确性。本研究结果为高寒草地经济作物的合理管理与利用以及黄河源区生态建设提供一定的理论和实践基础,也为草地生态系统生物多样性的监测和保护提供新方法和手段。
