香青兰总黄酮调节MEK/ERK信号通路诱导BMSCs增殖及成骨分化的作用
2022-03-21王俊井邵银初谌少波
王俊井,邵银初,谌少波,李 浩
(中国人民解放军联勤保障部队908医院骨科,南昌 330002)
随着年龄的逐渐增长,特别是妇女绝经后雌激素的减少,骨髓间充质干细胞(BMSCs)分化的成骨细胞逐渐减少,破骨细胞生成增多,从而引起骨质疏松症,容易导致老年人病理性骨折,因此促进BMSCs向成骨细胞分化,对于骨质疏松症的防治具有重要意义[1]。香青兰是兰科青兰属植物,广泛分布于我国西南地区,可入药,具有清热解毒、利咽止咳、凉肝止血之功效,主要用于呼吸道感染性疾病的治疗,临床上也用来治疗原发性高血压、冠状动脉粥样硬化性心脏病等心血管疾病[2]。近年来,随着中医的快速发展,对香青兰的研究愈加深入,从中分离出的黄酮类化合物受到重点关注,因其广泛的生物学功能,成为各研究学者的研究热点[3]。香青兰总黄酮具有调节内分泌和心血管系统的功效,对该类疾病效果显著,其内分泌调节功能使其对骨髓微环境的改变成为可能[4]。细胞外调节蛋白激酶(ERK)包括ERK1和ERK2,其能够将信号从细胞表面受体传导至细胞核受体[5]。作为分裂原的活化抑制剂,MEK1、MEK2可通过调节Tyr和Thr双位点的磷酸化来激活ERK的表达[6]。MEK/ERK信号通路还参与多种细胞功能的调控,与多种疾病的发生、发展密切相关[7]。本研究探讨香青兰总黄酮是否能够通过调节MEK/ERK信号通路蛋白的表达来促进BMSCs细胞向成骨细胞分化,为其作用机制提供可靠的实验依据。
1 材料与方法
1.1 主要仪器与试剂
Thermo ScientificTMMultiskanTMGO型酶标仪(美国Thermo Fisher公司);BX53显微镜(日本奥林巴斯公司);胎牛血清(美国Sigma公司,批号:502198);地塞米松(美国Sigma公司,批号:514984);香青兰总黄酮(南京建成生物工程研究所,纯度:99.99%,批号:1248548);细胞计数试剂盒8(CCK-8 kit,南京宏哲生物有限公司,批号:201364)、碱性磷酸酶(ALP)染色试剂盒、ALP比活性试剂盒(武汉赛培生物科技有限公司,批号:20184512、20184577);MEK1、ERK1蛋白抗体(美国abcam公司,批号:201548、201574)。
1.2 实验方法
1.2.1 实验动物
SPF级4周龄SD大鼠,雌性,体重60~90 g,购自重庆医科大学实验动物中心,动物生产许可证号:SCXK(渝)2018-0003,动物使用许可证号:SYXK(渝)2018-0027,动物质量合格证号:10124584。本研究已获得中国人民解放军联勤保障部队908医院医学伦理委员会审查和批准,符合动物伦理学实验标准要求。
1.2.2 细胞培养与分组
颈椎脱臼处死大鼠,70%酒精浸泡大鼠10 min,于超净工作台上无菌条件下取出双侧股骨和胫骨,骨剪剪去骨干骺端,采用含10%胎牛血清的DMEM完全培养基(含100 U·mL-1青霉素+100 μg·mL-1链霉素的双抗)反复冲洗骨髓腔,冲洗液收集到50 mL 离心管中,制成细胞悬液,在1000 r·min-1条件下离心10 min,弃上清,加入DMEM完全培养基,吹打混匀,以5×106个·cm-2的密度接种于细胞培养瓶中,恒温培养箱(37 ℃,5%CO2)中孵育48 h。待细胞铺满瓶底后给予传代培养,取P3代细胞进行实验。将培养后的细胞分为空白对照组(完全培养基)、阳性对照组(50 μg·mL-1抗坏血酸+10 mmol·L-1β甘油磷酸钠+10-8mol·L-1地塞米松)[8]、香青兰总黄酮高浓度组(100 μg·mL-1)、香青兰总黄酮中浓度组(50 μg·mL-1)、香青兰总黄酮低浓度组(25 μg·mL-1)[9]。
1.2.3 CCK-8法检测细胞存活率
用CCK-8法检测BMSCs细胞的活力:胰蛋白酶消化对数生长期的BMSCs细胞2 min,充分吹打,制作成单细胞悬液,3000 r·min-1离心10 min后弃上清,加入含有10%胎牛血清的RPMI1640培养基,轻微吹打,重悬细胞,以每孔100 μL接种到96孔板,37 ℃、5%CO2培养24 h后更换新的培养基,按实验分组,加入不同浓度的香青兰总黄酮和阳性对照物,各组6个复孔,培养24 h,更换培养液后每孔加入10 μL CCK-8溶液,持续培养3 h,使用Bio-Rad微孔板阅读器读取450 nm处的吸光度,重复3次,通过吸光度计算各实验组细胞存活率。
1.2.4 ALP比活性检测
细胞处理及给药同1.2.2,于6孔板上给药培养7 d后,收集细胞于1.5 mL离心管中,每管加入400 μL 的10 mmol·L-1Tris-HCL(含0.1%Triton X-100),吹打混匀,用超声细胞破碎仪破碎细胞,4 ℃下10 000 r·min-1离心10 min,取上清液于新的1.5 mL离心管中,根据ALP检测试剂盒说明检测细胞ALP活性,再根据蛋白测定试剂盒说明检测蛋白含量,两者比值即为ALP比活性。
1.2.5 ALP染色
细胞处理及给药同1.2.2,6孔板上培养细胞7 d 后取出,弃培养液,PBS冲洗,将固定液加入孔板中,作用1 min,流水冲洗2 min,晾干。滴加孵育液,于37 ℃恒温箱中孵育15 min,流水冲洗2 min,晾干,苏木精染色5 min,流水冲洗2 min,晾干,光学显微镜下观察各孔细胞形态。
1.2.6 MEK1、ERK1蛋白表达水平的检测
细胞处理及给药同1.2.2,在无菌操作台上取出24 h培养后细胞,弃去培养液,加入胰蛋白酶消化细胞,3000 r·min-1离心10 min后转移至1.5 mL EP重悬,蛋白提取试剂盒提取细胞蛋白,收集于新的1.5 mL EP管中。经SDS-PAGE分离后,蛋白转移到硝化纤维素膜上,将膜放在5%脱脂奶粉溶液中封闭2 h,然后用特异性抗体MEK1、ERK1和β-actin在4 ℃条件下孵育12 h,PBS溶液清洗3次,每次10 min,进行二抗孵育2 h,然后在LAS 4000成像系统上显影。
1.3 统计学方法

2 结果
2.1 香青兰总黄酮对BMSCs细胞存活率的影响
香青兰总黄酮低、中、高浓度组和阳性对照组BMSCs细胞存活率显著高于空白对照组,且呈浓度依赖性(均P<0.05),高浓度组和阳性对照组作用相似,差异无统计学意义(P>0.05)。见表1。

表1 香青兰总黄酮对BMSCs细胞存活率的影响
2.2 香青兰总黄酮对BMSCs细胞ALP比活性的影响
香青兰总黄酮低、中、高浓度组和阳性对照组BMSCs细胞ALP比活性显著高于空白对照组,且呈浓度依赖性(均P<0.05),高浓度组和阳性对照组作用相似,差异无统计学意义(P>0.05)。见表2。
表2 香青兰总黄酮对BMSCs细胞ALP比活性的影响
2.3 各组BMSCs细胞ALP染色结果
随着香青兰总黄酮干预浓度的增加,BMSCs胞浆内棕红色逐渐加深,且咖啡色颗粒逐渐变大且增多,且香青兰总黄酮高浓度组与阳性对照组染色结果相似。见图1。
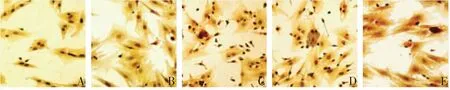
A:空白对照组;B:香青兰总黄酮低浓度组;C:香青兰总黄酮中浓度组;D:香青兰总黄酮高浓度组;E:阳性对照组。
2.4 香青兰总黄酮对BMSCs细胞MEK1、ERK1蛋白表达的影响
香青兰总黄酮低、中、高浓度组和阳性对照组BMSCs细胞MEK1、ERK1蛋白表达量显著高于空白对照组,且呈浓度依赖性(均P<0.05),高浓度组和阳性对照组作用相似,差异无统计学意义(P>0.05)。见表3,图2。
表3 香青兰总黄酮对BMSCs细胞MEK1、ERK1蛋白表达的影响
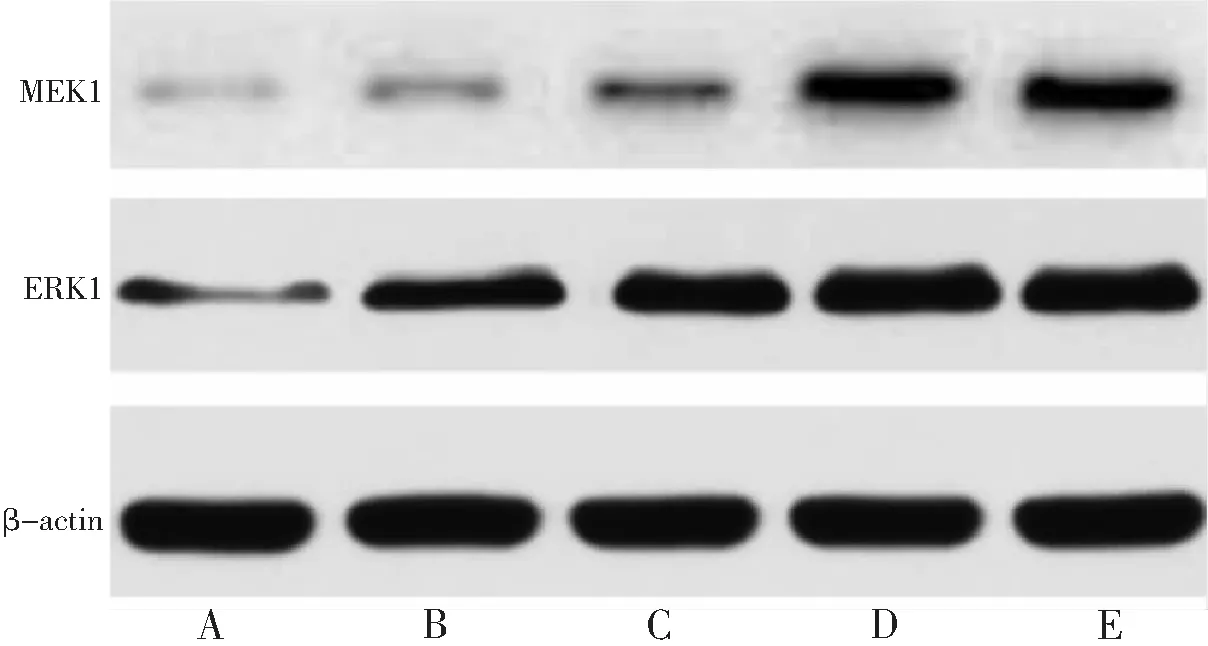
A:空白对照组;B:香青兰总黄酮低浓度组;C:香青兰总黄酮中浓度组;D:香青兰总黄酮高浓度组;E:阳性对照组。
3 讨论
BMSCs又称骨髓源性多能间充质干细胞,已被证实能向包括骨、软骨、肌肉和脂肪在内的所有间充质组织分化[10]。由于其具有多潜能的分化特性,且细胞易于获得和操作,骨髓基质干细胞已经成为组织工程应用和再生研究领域的理想候选者[11]。当上调与基质发育和成熟相关的基因表达时,可导致BMSCs在增殖完成后发生成骨分化,在分化过程中,由于基因和蛋白质表达的改变,BMSCs在细胞外基质矿化开始之前,经历了形态和功能的改变[12]。目前香青兰总黄酮是否具有成骨诱导作用尚不清楚,但是其可调节机体内分泌功能、改善机体内微环境的作用已被确定,这对BMSCs的分化可能具有促进作用[13]。本研究结果显示,在给予BMSCs不同浓度的香青兰总黄酮干预后,均可导致BMSCs分化数量增加,且随着香青兰总黄酮干预浓度的增加,BMSCs分化数量呈显著递增趋势,呈明显浓度依赖性。活跃的成骨细胞可释放出较多的ALP,因此ALP染色和ALP比活性可以反映出机体骨形成情况,本研究实验组中ALP染色颗粒的明显增加和ALP比活性的升高,均提示BMSCs向成骨分化。以上结果显示香青兰总黄酮可促进BMSCs向成骨细胞分化,其原因可能是香青兰总黄酮改善机体骨髓微环境,使得BMSCs利于分化。
MEK信号通路是由ERK、c-junnh2末端激酶(JNK)和p38 3种蛋白激酶级联而成的一种信号通路,它参与多种重要的细胞事件的进程,尤其是细胞增殖、分化和凋亡[14-15]。ERK通常被有丝分裂原和生长因子激活,在细胞生长、防止凋亡和细胞分化等方面有着重要的调节作用[16]。有文献[17]报道ERK和p38在在骨髓基质细胞成骨分化过程被激活。本研究通过检测骨髓间充质干细胞中ALP比活性、MEK1和ERK1信号通路蛋白的表达,证明激活MEK/ERK信号通路中相关蛋白表达,可促进细胞增殖能力。然而,也有研究[18]表明,MEK/ERK信号通路抑制细胞成骨分化,因此,MEK/ERK信号通路在成骨细胞分化中的作用目前尚有争议,使其成为本研究需要重点探讨的内容。本研究从细胞存活率、ALP比活性、MEK1及ERK1蛋白表达量等方面分析MEK/ERK信号通路在BMSCs成骨分化中的作用,阐明MEK/ERK信号通路在BMSCs成骨分化中的意义,结果显示MEK/ERK信号通路的激活可能有助于BMSCs成骨分化。其机制可能是香青兰总黄酮引起体内骨髓微环境发生改变,从而导致MEK/ERK信号通路激活,MEK1、ERK1相关蛋白表达水平增加,促进BMSCs成骨分化。且在本研究中,随着香青兰总黄酮干预浓度的增加,MEK1、ERK1蛋白表达水平呈显著递增趋势,表明香青兰总黄酮能够有效地激活MEK/ERK信号通路,其蛋白表达效果与香青兰总黄酮浓度密切相关,说明香青兰总黄酮可能是一种有效的促成骨分化的药物,而MEK/ERK信号通路就是其作用靶点之一。
综上所述,本研究结果初步证明了香青兰总黄酮可诱导BMSCs增殖及分化,其机制可能是香青兰总黄酮通过激活MEK/ERK信号通路,上调MEK1、ERK1相关蛋白表达来实现的,MEK/ERK信号通路极可能是BMSCs增殖及分化的重要调控靶点。
