硒对热应激条件下放牧绵羊瘤胃发酵及瘤胃微生物区系的影响
2022-03-19乌日力嘎阿拉达尔
乌日力嘎, 阿拉达尔, 崔 双, 晓 敏, 弓 剑
(内蒙古师范大学生命科学与技术学院,内蒙古呼和浩特 010022)
动物遭受超过自身体温调节能力的过高温度刺激时所产生的非特异性应答反应的总和称为热应激(闵力,2017)。 放牧条件,尤其是牧区放牧条件,从早到晚都在野外,长时间暴露在太阳直射下, 无法保证及时且充足的饮水很容易引发热应激。对于牧区放牧绵羊而言,在温度最高的中午到14:00 左右喜欢在柏油马路上聚集, 这更加重了热应激。热应激可导致反刍动物的采食量降低,抑制瘤胃蠕动,减少反刍次数,同时会对瘤胃上皮造成破坏且影响瘤胃乳头的发育(Mishra 等,1970)。热应激不仅会降低肠道菌群多样性及丰度(屈倩,2017),还会影响瘤胃内环境稳态和瘤胃菌群组成及数量,进而导致饲料效率及生产性能降低。热应激还可诱导脂质过氧化物和自由基产生, 进而影响动物机体的抗氧化和免疫功能(李晗等,2019)。因此,通过营养、管理等途径减缓或消除热应激对动物造成的不利影响, 对于保证动物的健康和生产具有重要的意义。
目前, 在动物日粮中添加抗氧化剂是缓解热应激最常用且可行的营养调控措施。 必需微量元素硒(Se)作为一种抗氧化剂通过合成硒蛋白来行使其强大的抗氧化能力以及抗炎作用, 因而具有缓解热应激的作用。 夏伟光(2017)在肉鸡饲粮中添加硒发现,其可以减轻热应激导致的氧化应激,提高机体的免疫应答能力。 Habibian 等(2015)报道, 硒能改善热应激引起的细胞因子分泌的异常和氧化损伤。劳雪芬(2015)研究发现,基础日粮中添加富硒女贞子对热应激山羊的免疫功能、 营养物质代谢和产奶量具有改善作用。 王尚(2017)研究报道, 酵母硒能提高热应激奶牛的免疫功能和抗氧化能力,最终达到缓解热应激的效果。 综上,目前有关硒和热应激关系的研究多集中在抗氧化和免疫功能方面。如前所述,热应激可影响瘤胃发酵以及胃肠道微生物活动, 而硒添加可改善瘤胃发酵(倪丽丽,2011),而且其在瘤胃中代谢可影响瘤胃微生物区系(Liu 等,2019),肠道微生物对硒的代谢活动可影响宿主对硒的利用(Kjernlie,2014)。 此外,目前相关的研究报道多集中在集约化饲养条件下, 很少关注放牧条件下热应激对动物的影响以及如何通过营养途径缓解这种不利影响。 鉴此,本文以放牧绵羊为研究对象,探讨了热应激条件下补硒对瘤胃发酵及瘤胃微生物区系的影响, 旨在为硒作为缓解热应激的营养添加剂在放牧绵羊生产实践中的应用提供理论依据。
1 材料与方法
1.1 试验设计与饲养管理 利用体内饲养试验,选用10 只体重(25.88±4.14)kg、年龄为7 ~8 月龄的健康杜寒杂交羊作为试验动物,采用单因子完全随机试验设计, 随机分为2 组, 每组5只,对照组日粮中不添加硒,试验组硒添加水平为0.2 mg/d(硒源为酵母硒)。
试验在放牧条件下于夏季温度最高的初伏到中伏期间进行,试验期共20 d,其中预试期5 d,正试期15 d,试验期内每日06:00 对硒添加组的羊只投喂酵母硒 (与100 g 玉米面混合投喂),08:00左右放牧,直到19:00。草场牧草种类分布见表1,其中主要的牧草种类及占比为:短花针茅(30%)、白皮锦鸡儿(20%)、冰草(3%)、山蒿(2%)、两裂薇菱菜(2%)、野韭(7%)、砂葱(7%)、冷蒿(2%)、银灰旋花(3%)、羊草(10%)、糙隐子草(4%)、长芒草(6%)、其他(4%)。 牧草中的平均硒含量为0.204 mg/kg DM。

表1 试验地草场中的牧草种类分布
1.2 样品采集与处理 采集试验草场中的牧草样本,烘干后粉碎,保存备用。 试验结束后在晨放前颈静脉采集血液于肝素钠的真空采血管中,静置10 min 后3500 r/min 条件下离心15 min,获得的血浆分装于离心管中。 试验结束后在晨放前通过口腔采集瘤胃液,经四层纱布过滤后立即用pH计测定瘤胃液pH,取1 mL 瘤胃液放入预先装有9 mL 盐酸(0.2 mol/L)的离心管中,用于NH3-N 测定, 取4 mL 瘤胃液放入预先装有1 mL 偏磷酸(25%)的离心管中,用于挥发性脂肪酸(VFA)测定,取5 mL 瘤胃液放入无菌离心管中,用于瘤胃微生物区系的分析。上述样品迅速投入液氮,带回实验室转入-80 ℃冰箱冻存备用。
1.3 测定指标与方法
1.3.1 热应激的判断 依据卢曾奎(2019)报道的温湿度指数(THI)计算公式判断动物是否发生热应激。
THI=Td-(0.31-0.31RH)( Td-14.4)(Td 为温度,RH 为相对湿度),THI ≥22.2 表示动物处于热应激状态。
1.3.2 牧草硒含量 参照索兰弟和李秉真(1991)描述的方法, 采用荧光分光光度法检测牧草样本中的硒含量。
1.3.3 血浆热休克蛋白-70 采用ELISA 法测定血浆热休克蛋白-70(HSP-70),试剂盒购于南京建成生物工程研究所。
1.3.4 瘤胃液pH 在预先校正好的pH 计的样品盘中滴入2 滴瘤胃液,读取数值,用蒸馏水清洗样品盘后重复测量3 次,计算平均值。
1.3.5 瘤胃液NH3-N 采用冯宗慈和高民(2010)描述的比色法测定瘤胃液NH3-N 浓度。
1.3.6 瘤胃液VFA 依据张英等(2013)描述的高效液相色谱法稍加调整测定瘤胃液乙酸、 丙酸和丁酸浓度。 色谱条件为: 柱温40 ℃, 流速1.0 mL/min,进样体积20 μL,流动相由0.02 mol/L 的NaH2PO4(流动相A)和100%的甲醇(流动相B)组成。 梯度洗脱条件为:0 ~4 min 为95%的流动相A 和5%流动相B;4.1 ~35 min 为90%的流动相A 和10%的流动相B;35.1 ~50 min 为95%的流动相A 和5%的流动相B。
1.4 数据统计分析 用Excel 2010 对数据进行初步整理,采用SAS 8.1 统计软件中的t 检验对所测指标进行统计分析。 利用美吉生信云平台依据相关方法对瘤胃微生物多样性、 物种组成及丰富度、物种差异以及主成分进行分析。 数据以“平均值±标准差” 表示。 用P<0.05 表示差异显著,0.05<P≤0.10 表示差异趋于显著。
2 结果与分析
2.1 试验期THI 的变化 由图1 可知, 整个试验期间除了第3 天和第9 天08:00 以及第13 天12:00 的THI 低于22.2 之外,其他时间的THI 指数均高于22.2。 整个试验期间每天的平均THI 均高于22.2,说明动物处于热应激状态。
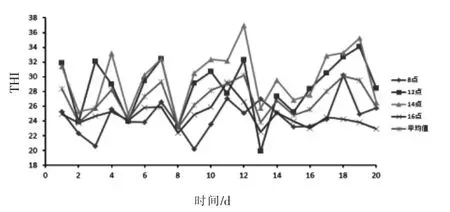
图1 试验期间THI 的变化
2.2 血浆HSP-70 含量的变化 如图2 所示,与对照组相比, 硒添加组绵羊血浆HSP-70 含量显著降低(P<0.05)。
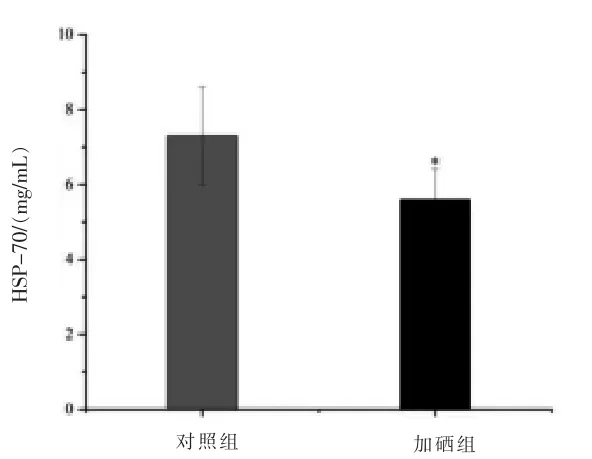
图2 对照组与硒添加组绵羊血浆HSP-70 含量
2.3 硒对热应激条件下放牧绵羊瘤胃发酵的影响如表2 所示,热应激条件下添加酵母硒对放牧绵羊的瘤胃pH 和乙酸、丙酸、丁酸浓度无显著影响,但可显著降低瘤胃NH3-N 浓度(P<0.05),对瘤胃总挥发性脂肪酸(TVFA)浓度有提高趋势(P=0.10)。
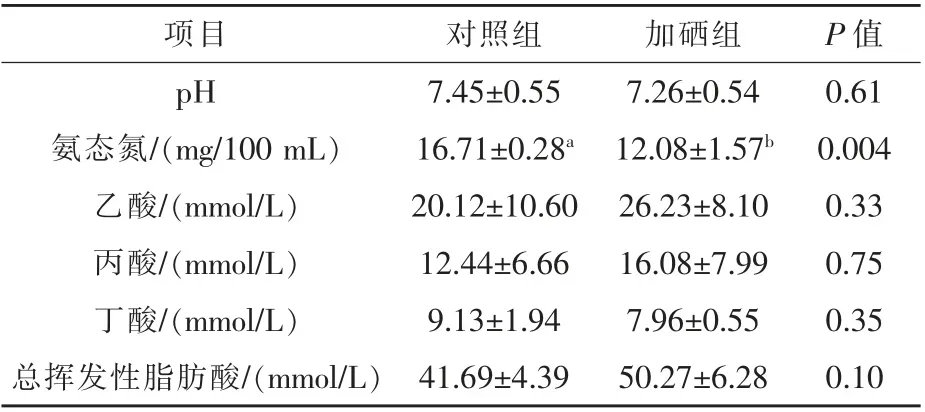
表2 硒对热应激条件下放牧绵羊瘤胃发酵的影响
2.4 硒对热应激条件下放牧绵羊瘤胃微生物区系的影响
2.4.1 测序结果及合理性
2.4.1.1 基于OTU 的Venn 图 Venn 图可统计多组或多个样本中所共有和独有的物种数目。 如图3 所示,通过测序共得到1912 个OTU,其中对照组OTU 数为1621 个, 加硒组OTU 数为1711个,共享OTU 数为1420 个,共享OTU 数占总OTU 数的74.27%。
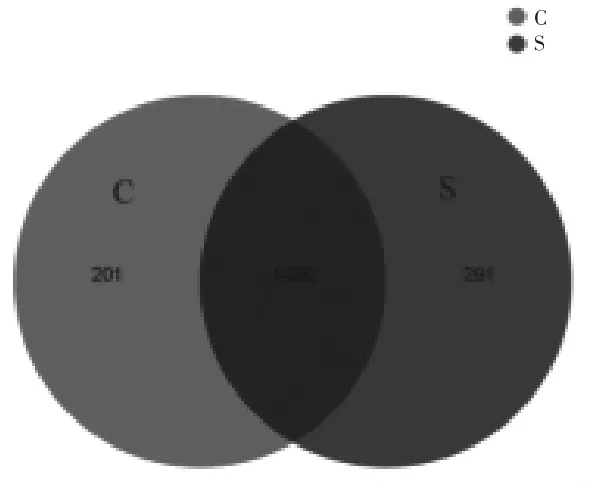
图3 对照组和硒添加组绵羊瘤胃微生物的Venn 图(基于OTU)
2.4.1.2 稀释曲线和覆盖度曲线 对照组和硒添加组绵羊瘤胃微生物稀释曲线和覆盖度曲线如图4。 随着测序深度的增加,稀释曲线呈现大幅度上升后趋于平缓, 说明深度覆盖到样本中的所有物种,足以代表样本中的物种丰富度。覆盖度曲线表明对照组和硒添加组物种覆盖度均在99%以上,可以完全反映微生物群落结构和种类。
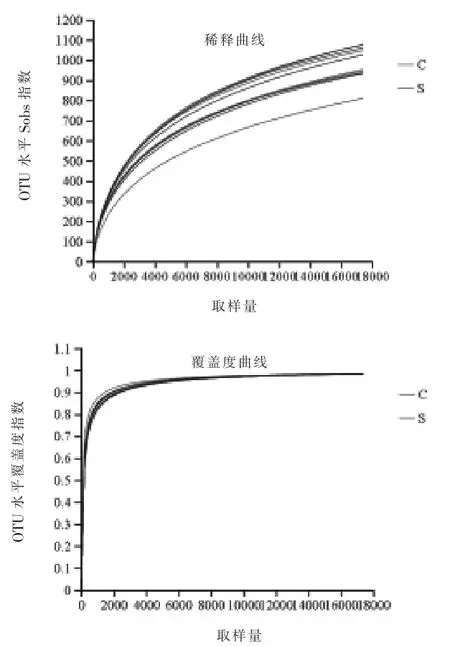
图4 对照组和硒添加组绵羊瘤胃微生物样本稀释曲线和覆盖度曲线
2.4.2 α-多样性 α-多样性中Sobs、Ace、Chao 和Shannon 指数越大,多样性越高;Simpson 指数越小,多样性越高。 如表3 所示,Sobs、Chao、Ace、Shannon和Simpson 指数无显著的组间差异(P>0.05)。

表3 硒对热应激条件下放牧绵羊瘤胃微生物多样性指数的影响
2.4.3 β-多样性 采用unweighted-unifrac 距离算法计算每个样本的菌群相似度, 分别在属水平和OTU 水平进行PCoA 分析(图5)。 对照组和硒添加组绵羊瘤胃微生物样本在属水平和OTU 水平均聚类于不同的坐标位置,具有显著的组间差异。
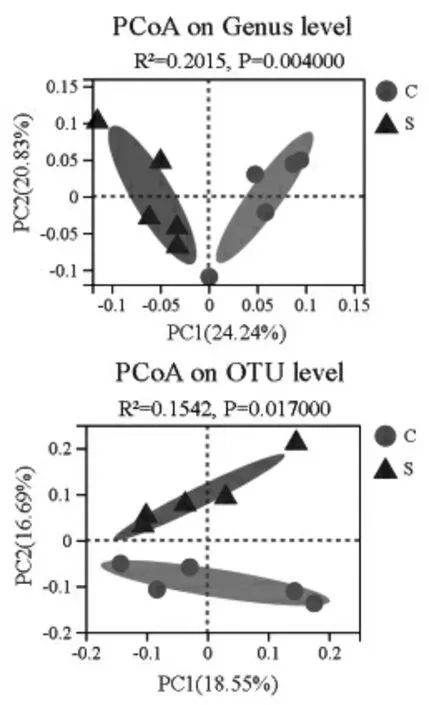
图5 对照组和硒添加组绵羊瘤胃微生物PCoA 分析(属水平和OTU 水平)
2.4.4 物种组成及丰富度
2.4.4.1 门水平 门水平瘤胃微生物群落组成相对丰度分布如表4 所示, 两组绵羊瘤胃优势微生物菌群主要分布在拟杆菌门(Bacteroidetes)和厚壁菌门(Firmicutes),这两类微生物在对照组中的比例分别为62.62%和31.28%, 在硒添加组中的比例分别为56.59%和37.42%。 次级优势菌群主要分布在蓝藻菌门 (Cyanobacteria)、 变形菌门(Proteobacteria)、螺旋体门(Spirochaetes)和髌骨细菌门(Patescibacteria);others 为相对丰度低于1%的菌门。
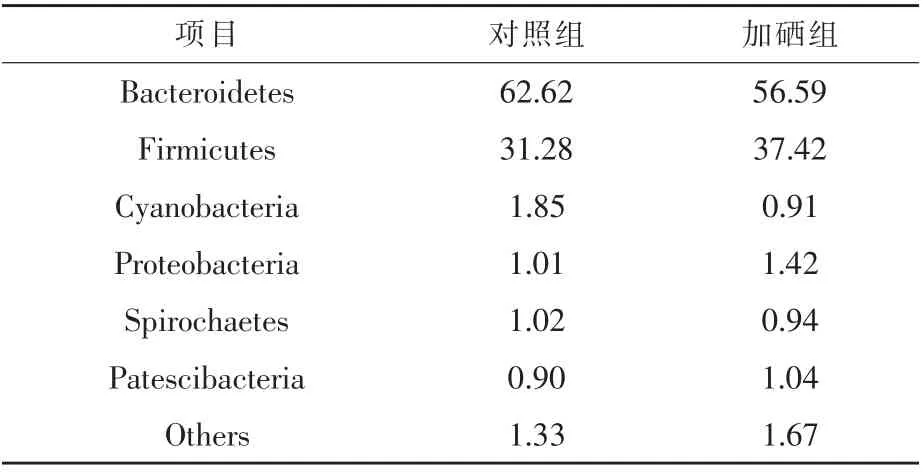
表4 对照组和硒添加组绵羊瘤胃微生物物种组成(门水平)%
如图6 所示,与对照组相比,硒添加组绵羊瘤胃内梭杆菌门(Fusobacteria)相对丰度显著提高(P<0.05)。
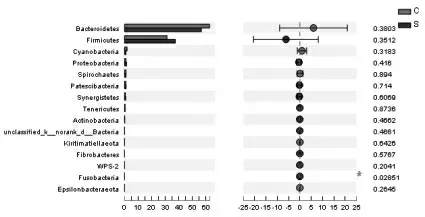
图6 对照组和硒添加组绵羊瘤胃微生物相对丰度前15 位的物种(门水平)
2.4.4.2 属水平由表5 可知, 热应激绵羊在属水平上检测到的相对丰度大于1%的瘤胃微生物有:普雷沃氏菌属1(Prevotella_1)、理研菌科RC9菌群(Rikenellaceae_RC9_gut_group)、norank_f_F082、 普雷沃氏菌科UCG-003 菌属(Prevotellaceae_UCG-003)、 未排位的鼠杆菌科(norank_f_Muribaculacae)、 瘤胃球菌科NK4A214 菌群(Ruminococcaceae_NK4A214_group)、未分类的毛螺菌科(unclassified_f_Lachnospiraceae)、解 琥珀酸弧菌属(Succiniclasticum)、 月形单胞菌属3(Selenomonas_3) 和月形单胞菌属1 (Selenomonas_1)、未分类的韦荣氏球菌科(unclassified_f_Veillonellaceae)、 克里斯滕森菌科R-7 菌群(Christensenellaceae_R-7_group)、未分类的普雷沃氏菌科、 未排位的拟杆菌科RF16 菌群(norank_f_Bacteroidales_RF16_group)、奎因氏菌属(Quinella)、普雷沃氏菌科UCG-001 菌属、瘤胃球菌科UCG-014 菌属、norank_o_Chloroplast、 丁酸弧菌属2(Butyrivibrio_2)、未排位的拟杆菌目(norank_o_Bacteroidales)、 瘤胃球菌科UCG-002 菌属、 Eubacterium_coprostanoligenes_group。 其中,普雷沃氏菌属1、 理研菌科RC9 菌属和norank_f_F082 为优势菌属。

表5 对照组和硒添加组绵羊瘤胃微生物物种组成(属水平)%
图7 所示,在属水平相对丰度前15 位的物种中, 硒添加显著提高了未排位的鼠杆菌科和奎因氏菌属的相对丰度(P<0.05)。 图8 所示,属水平相对丰度15 位之后的物种中,硒添加显著提高了瘤胃球菌科UCG-010 菌属、 艰难杆菌属(Mogibacterium)、毛螺菌科FE2018 菌群、毛螺菌科AC2044 菌群、 未排位的克里斯滕森菌科以及FD2005、Family_XIII_AD3011_group、Caviibacter和Kandleria 的相对丰度(P<0.05);显著降低了假单 胞 菌 属 (Pseudomonas) 和M2PT2-76_termite_group 的相对丰度(P<0.05)。

图7 对照组和硒添加组绵羊瘤胃微生物相对丰度前15 位的物种(属水平)

图8 对照组和硒添加组绵羊瘤胃微生物相对丰度15 位之后有显著差异的物种(属水平)
2.4.5 LEfSe 物种差异判别分析 如图9 所示,门到属水平LEfSe 物种差异判别分析结果表明,对照组中月形单胞菌属 3、g_norank_c_MVP_15、c_MVP_15、o_norank_c_MVP_15、f_norank_c_MVP_15、g_Anaerococcus、f_Family_XI_o_Clostridiales、假单胞菌属、 假单胞菌科(Pseudomonadaceae)、g_M2PT2_76_termite_group 以及琥珀酸单胞菌属(Succinatimonas)显著富集,而加硒组中奎因氏菌属、鼠杆菌科、鼠杆菌科未排位的属、瘤胃球菌科UCG-004 、UCG-005 和UCG-010 菌属、普雷沃氏菌科NK3B31 菌群、FD2005、梭杆菌门、梭杆菌目(Fusobacteriales)、梭杆菌纲(Fusobacteriia)、g_Family_XIII_AD3011_group、 毛螺菌科FE2018菌群、巴斯德氏菌科(Pasteurellaceae)、巴斯德氏菌目 (Pasteurellales)、 艰难杆菌属以及g_Bibersteinia 显著富集。
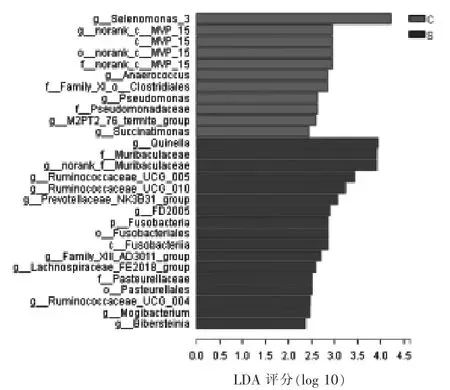
图9 对照组和硒添加组绵羊瘤胃微生物LEfSe 物种差异判别分析(LDA=2)
3 讨论
3.1 硒对热应激条件下放牧绵羊血浆HSP-70 的影响 HSP-70 是机体为了抵抗应激引起的组织细胞损伤而产生的一组特殊的蛋白质 (薛惠琴等,2016)。热休克蛋白是家族中最保守、含量最丰富的一种非特异性内源保护蛋白,其参与机体免疫功能调节以及作为分子伴侣修复损伤,进而提高细胞耐受力和抗细胞凋亡, 但其保护作用具有时效性,而且过度生成会影响细胞的正常功能,进而导致细胞损伤甚至死亡(Malyshev,2013;陈艳,2007;王枫等,2000)。热应激条件下,HSP 作为一种细胞防御和修复机制被诱导表达,因而血液或组织中的水平会提高(Slimen 等,2016)。 研究表明,在HSP 家族中,HSP-70 对温度最为敏感, 因而可作为热应激的生物指示物(Nagayach 等,2017;Yadav 等,2016)。 薛惠琴等(2016)研究发现,日粮中添加硒显著下调了蛋鸡肝脏HSP-70 mRNA 的表达。 施力光等(2016)在补饲硒和维生素E 对高温季节山羊精液品质、抗氧化酶活性及热休克蛋白表达的影响研究报道中提出,加硒显著降低了山羊精液HSP-70 mRNA 的表达水平。 本试验也发现,热应激条件下加硒显著降低了绵羊血浆HSP-70 的含量,表明硒具有缓解热应激的作用。
3.2 硒对热应激条件下放牧绵羊瘤胃发酵的影响瘤胃pH、NH3-N 和VFA 是判断瘤胃发酵情况的重要参数(徐振松,2014)。 瘤胃正常pH 为5.5 ~7.5,过高或过低均不利于瘤胃微生物的繁殖,进而导致瘤胃发酵模式改变。 一些研究表明,热应激会导致瘤胃pH 降低(蔡丽媛,2015),但也有关于热应激提高瘤胃pH 的报道(张灿,2016;温雅俐,2011)。尽管多数研究支持热应激可降低瘤胃pH, 但本试验条件下对照组绵羊的瘤胃pH 仍在正常范围内(7.45),并没有发生明显的降低,这可能与试验动物处于完全放牧状态有关。 本研究结果显示,硒添加对热应激条件下放牧绵羊的瘤胃pH 无显著影响,这与多数研究支持的硒(亚硒酸钠和硒酸钠、包被硒、蛋氨酸硒及酵母硒)不影响瘤胃pH 的结论一致(张清月等,2018;孙国平等,2013;倪丽丽,2011;刘强等,2007)。 适宜的瘤胃NH3-N 浓度是瘤胃正常发酵的重要条件。 张灿(2016)研究表明,热应激会提高藏绵羊瘤胃NH3-N 浓度。 在奶牛(孙国平等,2013)、湖羊(朱翱翔等,2017)等动物日粮中添加硒发现,可显著降低瘤胃NH3-N 浓度,这与本试验结果一致。 Nonaka 等(2008)以及Freestone 和Lyte(2010) 均报道, 热应激显著降低了反刍动物瘤胃TVFA 的浓度。 朱翱翔等(2017)在湖羊日粮中添加0.3 mg/kg 的酵母硒提高了瘤胃TVFA 的含量。与上述研究结果相近,本试验中加硒组与对照组相比具有提高绵羊瘤胃TVFA 含量的趋势。 从瘤胃发酵指数看出,NH3-N 浓度显著降低,TVFA 浓度也有提高趋势,而pH 没有显著差异,可能与硒提高了瘤胃微生物丰度从而增强了微生物的降解能力有关。
3.3 硒对热应激条件下放牧绵羊瘤胃微生物区系的影响 在瘤胃微生物门水平上, 拟杆菌门和厚壁菌门为优势菌门。众所周知,厚壁菌门属于革兰氏阳性细菌,主要产生丙酸和丁酸盐,拟杆菌门属于革兰氏阴性细菌,主要分解纤维及难消化的物质并产生挥发性脂肪酸。研究发现,热应激导致厚壁菌门丰度逐渐降低,拟杆菌门丰度逐渐增加(钟书,2019)。 本试验发现,尽管两组间厚壁菌门和拟杆菌门相对丰度无显著差异,但与对照组相比,硒添加组拟杆菌门相对丰度降低6.03%、 厚壁菌门相对丰度提高6.14%。本试验发现,与对照组相比,硒添加组绵羊瘤胃中梭杆菌门的相对丰度显著提高。 梭杆菌门具有发酵氨基酸和葡萄糖的作用(陈双双等,2018),因而在热应激条件下添加硒可能提高了营养物质的利用率。 但梭杆菌门相对丰度较低(低于1%),其在缓解热应激过程中的贡献有待进一步研究。
普雷沃氏菌和新月形单胞菌属于淀粉降解菌,而瘤胃球菌和毛螺菌属于纤维降解菌。多数研究表明, 热应激条件下瘤胃淀粉降解菌数量显著增加,纤维降解菌数量显著降低(Uyeno 等,2010)。 例如,钟书(2019)在THI 介导的山羊瘤胃细菌群落变化的研究中发现,随着热应激强度的增强,瘤胃中普雷沃氏菌属的丰度增加;蔡丽媛(2015)研究发现,热应激显著提高了山羊瘤胃中新月形单胞菌的丰度,显著降低了瘤胃中纤维降解菌的丰度。 本研究发现,对照组中新月形单胞菌属3 显著富集,加硒组中瘤胃球菌科UCG-004、UCG-005 和UCG-010菌属以及毛螺菌科FE2018 菌群显著富集,而且热应激条件下添加酵母硒显著提高了放牧绵羊瘤胃中瘤胃球菌科UCG-010 菌属以及毛螺菌科FE2018 和AC2044 菌群的相对丰度。
瘤胃微生物的活动与瘤胃发酵指标密切相关。 研究表明, 瘤胃中新月形单胞菌丰度的增加有利于丙酸和乳酸发酵(马峰涛,2010;于鸿玲等,2006)。尽管本试验对照组中月形单胞菌属3 显著富集,但两组间丙酸生成无显著差异,推测可能与试验动物处于完全放牧状态(日粮缺少丙酸发酵底物)有关。 研究表明,瘤胃球菌和毛螺菌会促进乙酸和丁酸 的生成 (Sibai 等,2020; 李 晗等,2019)。 而且丁酸具有促进瘤胃乳头发育的作用(Mccurdy 等,2019)。 本试验尽管没有发现两组间乙酸和丁酸生成有显著差异, 但TVFA 的生成在加硒组中有提高趋势, 这可能与加硒组中瘤胃球菌和毛螺菌的显著富集有关。 研究发现, 40 ℃的环境温度条件与35 ℃相比, 每千克DMI 的瘤胃甲烷生成显著提高(Yadav 等,2016),这与热应激导致瘤胃中与甲烷生成相关的微生物的活性改变以及瘤胃内温度的提高有关(King 等,2011)。 瘤胃中存在的产甲烷菌主要为甲烷短杆菌属、 甲烷细菌属、甲烷微菌属和甲性烷八叠球菌属。 此外,瘤胃中存在一些微生物,如奎因氏菌属和Kandleria,其丰度的提高与降低甲烷生成有关(Kumar等,2018)。本试验中,添加酵母硒显著提高了热应激条件下放牧绵羊瘤胃中奎因氏菌属(丰度前15位)和Kandleria 的丰度,并且在加硒组中显著富集,推测硒可降低热应激诱导的瘤胃甲烷生成。但本研究并未检测瘤胃中甲烷的生成, 有必要在今后的研究中进一步探讨。目前,关于鼠杆菌科功能的研究较少,一些研究发现,鼠杆菌科与丙酸产生和长寿有关(Mccurdy 等,2019),高温环境下老鼠肠道中鼠杆菌科的丰度显著降低 (Chevalier 等,2020)。 本试验结果显示,硒添加显著提高了瘤胃中鼠杆菌科丰度,且在加硒组中显著富集,提示硒抑制了热应激诱导的瘤胃中鼠杆菌数量的降低。
综上所述, 硒可通过影响瘤胃微生物区系实现对热应激的缓解作用。本研究还发现,很多未知功能的瘤胃微生物在对照组和硒添加组之间的差异, 这些微生物活性的改变对热应激的缓解是否有作用随着其功能的揭示有待于进一步研究。
4 结论
日粮中添加酵母硒对放牧绵羊的热应激具有缓解作用,这种缓解作用与硒改变瘤胃微生物菌群组成进而改变瘤胃发酵以及营养物质的利用有关。
