不同流态中长江江豚的栖息活动比较
2022-03-19王子璇陈敏敏王康伟连玉喜于道平
王子璇 陈敏敏 王康伟 连玉喜 于道平*
(1 水生生物保护与水生态修复安徽省高校工程技术研究中心,安庆 246003)(2 安庆师范大学生命科学学院,安庆 246003)
长江江豚(Neophocaena asiaeorientalis)是鼠海豚科(Phocoenidae)中唯一的淡水齿鲸(Zhouet al.,2018),目前数量1 012 头(Meiet al., 2014),仅分布在长江中下游及其毗邻的鄱阳湖和洞庭湖,2013年被IUCN 列为极危物种(CR),全流域已建立8 个自然保护区和4 个迁地保护区,并已实施10 年禁渔,目前种群下降趋势初步得到遏制(Huanget al.,2020)。但是,水利工程(华元渝和陈佩薰,1992)、桥梁工程(于道平等,2002)、航道整治(于道平等,2012)、护岸工程(陈敏敏等,2018) 等人为活动,仍持续造成长江江豚栖息地水文环境的破坏。
长江江豚在长江干流中呈斑块状分布,92%出现在离岸400 m 内(张先锋等,1993),喜好在鹅头型分汊河道的缓水区,以及毗邻芦苇沼泽或淹没沙丘的水域(于道平等,2005)。在鄱阳湖江豚种群数量调查时,发现江豚经常在定置网密集的敞水区活动,认为江豚集群活动与鱼类资源有关(肖文和张先锋,2002)。对长江八里江段江豚的栖息地利用的研究,也认为江豚活动与饵料鱼类聚集相关,对水体流速和透明度没有明显的选择(魏卓等,2003);东洞庭湖江豚活动头次与鱼类密度显著正相关,可能有随鱼群迁移的行为(王崇瑞等,2019),甚至夜晚随鱼群进入危险港区觅食(Wanget al., 2015)。也有研究表明,江豚的活动与小型鱼类数量相关,而与渔获物总量无关(Zhanget al.,2015;张晓可等,2016;连玉喜等,2020)。但是,在一些特殊水文环境中,江豚活动有明显的季节性变化,例如鄱阳湖湖口的长江江豚活动,夏季明显偏多(Kimuraet al., 2012),而安庆江段的皖河口却正好相反(Chenet al.,2017)。
河流中生物群落,包括鱼类聚集及其饵料资源明显受河流水文条件的影响(Freeman, 2001;Brookset al., 2005)。大西洋鲑鱼(Salmo salar) 产卵偏好的水域,与水力参数(深度、速度和弗劳德数)和沉积变量有关(Moriet al.,1998);在影响栖息地质量的众多环境因子中,栖息地水动力学特征最能反映中华鲟(Acipensor sinensis)对栖息环境微观机理的响应(杨宇等,2007)。除此之外,四大家鱼对产卵场的特定地形和水动力条件有选择(柏海霞等,2014),也体现了水力学应用在水生生物栖息地选择研究中的价值。
水深、流速、水温、水体透明度、鱼类资源等生态因子以及底质、江心洲、河道弯曲率等河床地貌因子,虽然能表象解释淡水豚栖息地主要特征,但是没有从本质上对形成机制或驱动因子进行研究。虽然国内淡水豚研究涉及回水区(Huaet al.,1989)、流态(于道平等,2012)、挑流和涡漩(于道平等,2002)等,但是没有涉及长江江豚对这些常见流态是否有选择。本文根据1993—2021年,15次野外考察资料,统计分析不同流态中长江江豚目击率和集群规模,比较研究江豚栖息活动的水文特征,尝试运用水力学解释淡水豚栖息地选择。
1 研究方法
1.1 研究区域
研究江段为长江下游湖口至梅龙,总长213 km,由7 个自然分汊河段组成,包括10 个江心洲,4 个河流入口和3个通江湖泊闸口(图1)。
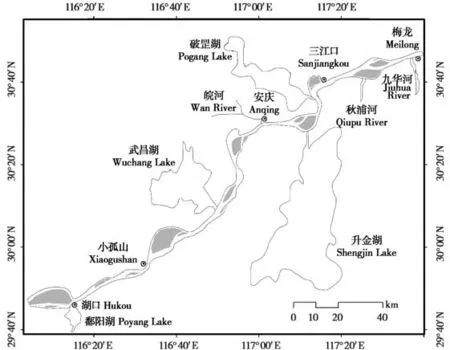
图1 长江下游湖口至梅龙段地理位置示意图Fig. 1 The geographical location from Hukou to Meilong section in the Yangtze River
根据长江下游梅龙镇大通水文站1955—2020年水文统计资料,长江下游湖口至梅龙江段全年水位3~15 m(黄海高程),每年10 月至次年3 月的黄海高程8 m以下称为枯水期,每年4—9月8 m 以上称为丰水期。
1.2 流态分类
每一个分汊河段至少有1个江心洲,其上游的深泓分成两支进入汊道,在洲尾下游汇合进入单一河段(图2)。

图2 分汊河段流态分布示意图. 三角形表示洲头分流区(ZT)或洲尾汇流区(ZW) 和河口汇流区(HK),弧形表示边滩分离区(BT),其他水域即为深水区(SS),实线为深泓线Fig. 2 Schematic of the flow patterns in the branching river. Trian‐gle indicates bifurcation upon bar (ZT) or confluence below bar (ZW)and estuarine (HK), arcs indicates separation aside shoal (BT), the oth‐er water means deep channel(SS),solid line indicates deep stream
由于长江中不同的河床地貌引起的水流阻力、流速和方向存在差异,形成了不同的流态(姚仕明等,2006),分汊河段的流态主要有:(1)洲头分流区:单一河段向分汊河段过渡时,河宽增大,深泓线在洲头分成干支两股水流,两侧形成指向岸边的横向流,以洲头为中心形成三角形的分流区(Leop‐old and Wolman,1957)。分流点至洲头纵轴线,纵向流速极低,水深一般由5 ~ 6 m 递减至0 m。(2)洲尾汇流区:分汊河段江心洲两侧干支流在洲尾汇集,相互顶托,尾下部延伸一条沙脊埂和Y型冲槽,以洲尾为中心形成三角形的汇流区(王晓刚等,2008)。沿沙脊埂从洲尾至汇流点,纵向流速很小,水深由0 m递增至5~6 m。(3)河口汇流区:径流或大型通江湖泊与长江交汇,形成一个斜向下游的冲坑,以河口上端为中心形成三角形的汇流区(Bironet al., 1996),河口上下两侧形成浅滩。由于河流(如皖河)和湖泊(破罡湖)流量近似,本文统一记为河口汇流区。(4)边滩分离区:由弯曲河道的离心力或交汇水域的切应力形成的横向流,导致近岸的水流减缓或反向流动,形成一个弧形的边滩(Best and Reid,1984)。(5)深水区:指不在上述流态范围内,水深6 m以上的水域。
1.3 数据来源
本文数据取自1993—1995 年铜陵淡水豚国家级自然保护区科考资料和2016—2021 年安庆师范大学科考资料,合计15 次野外考察。每次考察,租用相同的2 条渔船,每条渔船马力35 匹,每条渔船聘请3 名专业渔民进行观察,这些渔民都有20 年以上江豚野外考察经验。研究人员填表记录观察到江豚的时间、GPS 坐标点、数量、近岸距离及环境因子。本文中的样本指在考察江段中持续航行距离100 km 以上的记录数据,样本总数15 个,其中枯水期10 个,丰水期5 个。累计记录长江江豚412 次,累计1 117 头(表1)。
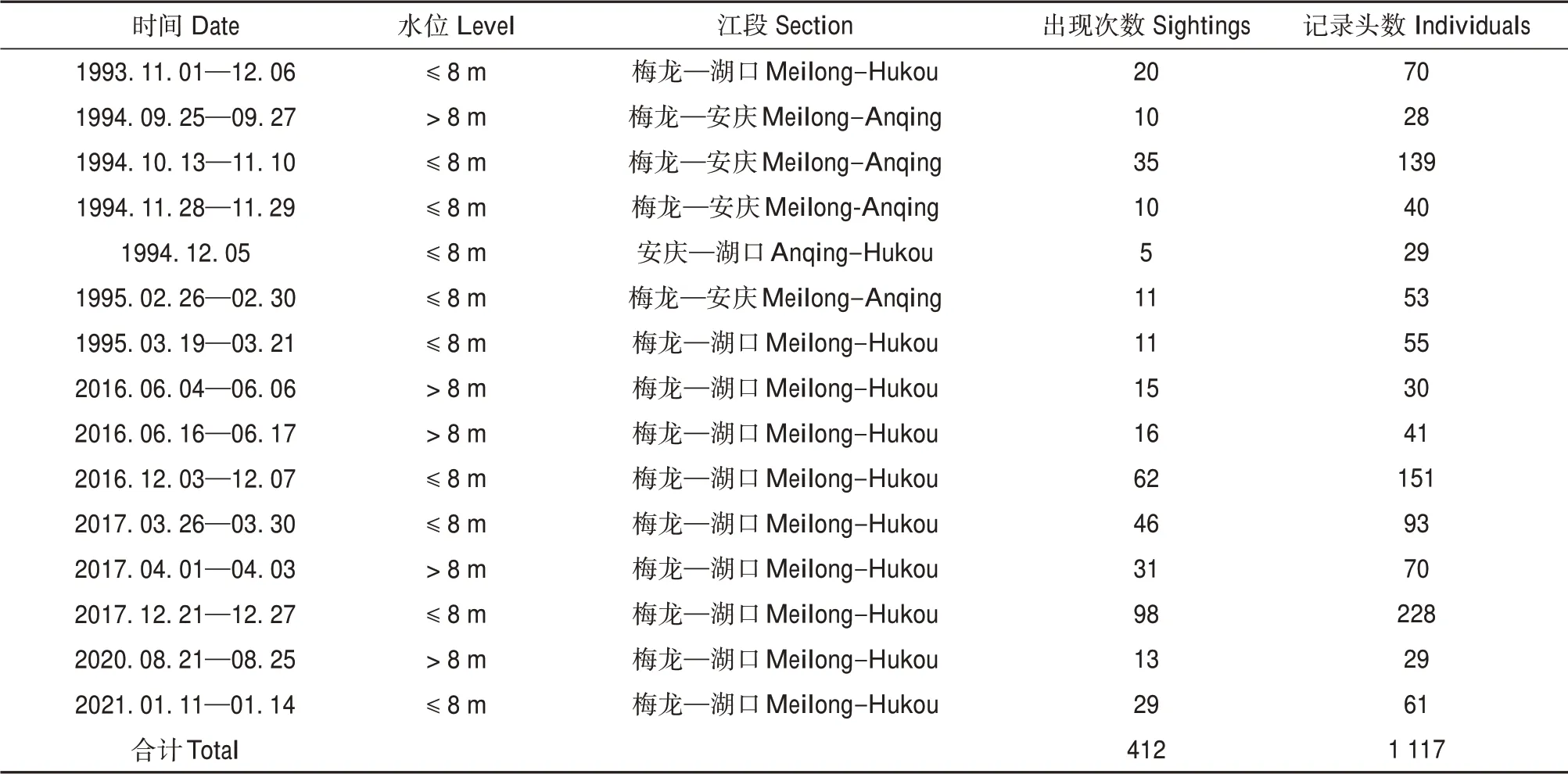
表1 1993—2021年长江江豚野外考察记录Table 1 Records in field of Yangtze finless porpoise between 1993 to 2021
根据手持GPS 航线,以洲头和洲尾端点为半径,500 m 水域内记录的江豚分别属于洲头分流区(ZT)和洲尾汇流区(ZW);以边滩弧线的中点、河口包括闸口宽度中点为半径,500 m 水域内记录的江豚分别记入边滩分离区(BT) 和河口汇流区(HK);凡在河道内,但不在上述4种流态范围内出现的江豚,统一归入深水区(SS)。
1.4 数据分析
提取每个样本的长江江豚记录次数、5 种流态中的次数和每次记录的头数。通过下式计算出流态中的目击率,即:
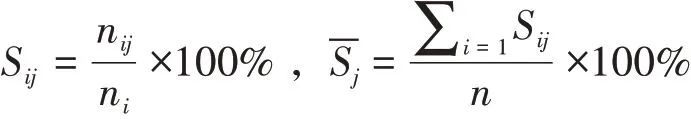
式中,n为样本数,ni为第i样本中记录的次数,nij为第i样本j流态中记录的次数,Sij为样本i中j流态的目击率,为j流态的目击率均值。同理,可计算出样本i中j流态中的集群均值Gij及j流态中的集群均值,其中N为每次记录的头数,即:

目击率和集群均值均进行正态分布和方差齐性检验,然后运用R4.0.2(R Core Team,2020)对各种流态中目击率和集群均值进行单因子方差分析(Oneway ANOVA)和Tukey多重比较,对枯水期和丰水期进行独立样本t检验,显著性判定标准为P≤0.05。
2 结果
2.1 江豚目击率
长江下游梅龙至湖口段的10个江心洲、4个河流入口和3 个通江湖泊闸口,都有江豚栖息活动。长江江豚在洲尾汇流区目击率较高,深水区最少(图3),但流态之间的目击率差异不显著(df= 4,F= 0.976,P= 0.426);部分流态的目击率在丰水期与枯水期有差异,即在洲头分流区中,丰水期江豚目击率明显高于枯水期(P= 0.044),而河口汇流区却相反,且差异显著(P=0.049)(图3)。

图3 长江江豚在各流态中目击率Fig. 3 Encounter rate of Yangtze finless porpoise in the flow patterns
2.2 江豚集群均值
无论枯水期还是丰水期,江豚在流态之间的集群均值差异都不显著(df=4,P=0.101),其中深水区集群均值全年都较小,而河口汇流区丰水期最低(图4)。长江江豚枯水期集群均值为2.778±2.106,略高于丰水期的集群均值2.329 ± 1.554,但两者差异不显著(P= 0.134);枯水期河口汇流区江豚集群均值(3.111±0.765)明显高于丰水期(1.600±1.140)(df= 13,P= 0.007),其他流态中江豚集群在枯水期与丰水期没有显著性差异(P>0.05)。
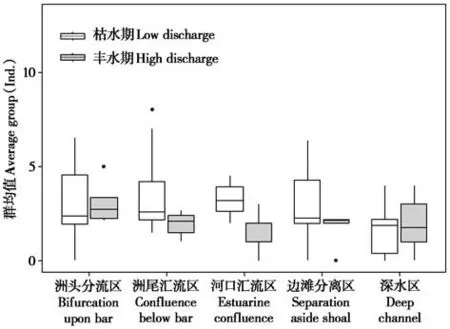
图4 长江江豚在流态中集群规模Fig. 4 Cluster size of Yangtze finless porpoise in the flow patterns
3 讨论
3.1 江豚对流态选择利用
本文结果表明,全年在江心洲目击江豚的概率约50%,与许多学者报道的江心洲是江豚重要活动水域较为一致(Zhanget al., 2013; Chenet al.,2020)。丰水期长江江豚在洲头分流区目击率明显高于枯水期(P< 0.05),丰水期长江江豚在洲头分流区停留时间相对较长,可能因为洲头浅水域开阔安静,可以规避外界干扰,有利于繁殖和抚育活动。
长江江豚在流态之间的目击率和集群均值差异都不显著(P> 0.05),这个结论也进一步提示,江豚在分汊河段内不同流态形成的食物斑块之间短距离频繁移动,均值统计分析几乎没有意义。但是,本文报道的枯水期河口(含闸口) 汇流区江豚目击率和集群均值都明显增大(P< 0.05),与于道平等(2005) 报道的春季江豚在支流交汇水域偏爱指数不高,趋于随机选择,以及皖河口每年4—10 月几乎没有江豚活动较为一致(Chenet al.,2017)。由此可见,河口包括闸口汇流区是长江江豚冬季索饵的重要水域。由于河口汇流区具有多样的地貌特征和复杂的水动力条件,许多无脊椎动物会充分利用汇流区作为栖息地(Beckmannet al., 2005),也是鱼类由长江干流返往支流或湖泊的主要节点(Czegle´diet al.,2016)。丰水期长江干流流量达90 000 m3/s,而径流汇入的流量较小,因此河口汇流区水动力条件如雷诺数(Rr)、动能梯度及涡量都有很大差异(Constantinescuet al.,2011)。丰水期江豚很少在河口汇流区索饵,是食物资源组成不适合还是捕食能耗大,尚需要进一步研究。
分离区存在于弯曲河段的凸岸、分流区两侧或者汇流区下游一侧,甚至桥梁、礁石下游也有分离区存在,所以有时又称回水区,即存在反向水流(Best and Reid, 1984; Ashmoreet al., 1992)。长江江豚在分离区的目击率和集群规模不及洲头分流区或洲尾汇流区,原因可能是分离区与这些流态相邻,流态不易辨别。野外考察时,江豚受惊后快速离开分离区,容易把这些记录误归入分流区或汇流区。
3.2 人类活动对流态的影响
汇流区范围和水动力条件,是由两支水流的流量比和入汇角度决定(Rhoads and Kenworthy,1995)。近年来的航道治理,增加了主汊流量,却降低了支汊的分流比,结果导致洲尾汇流区偏移或萎缩,长江江豚栖息活动明显受影响(于道平等,2020)。此外,航道整治的束水工程,一般建在洲头分流区,或河道自然演变,都会使分流点偏移或下挫,结果导致洲头被冲刷,长江江豚在洲头分流区活动减少或消失(于道平等,2002,2012)。
江豚重要栖息地如洲头、洲尾、边滩和河口的水文环境,都是由河床地貌及相应流量联合塑造的特殊流态,生活在其中的水生生物群落聚集与扩散,受水动力驱动下周期性变化(Shieldset al., 2003; Humphrieset al., 2008)。但是,长江江豚对河流动力学响应机制研究较少。本文对此仅作初步探究,研究结果可为江豚就地保护提供指导,例如安庆长江江豚省级自然保护区,2020 年地方立法就明确禁止枯水期船舶由皖河口进入支流中,强化了重要流态的保护力度。
