雄性麋鹿胆量和侵犯耦合与等级序位变化的关系
2022-03-19孟庆辉宋苑单云芳李俊芳白加德孟秀祥柏超张成林
孟庆辉 宋苑 单云芳 李俊芳 白加德孟秀祥 柏超 张成林
(1 北京市科学技术研究院,北京麋鹿生态实验中心,北京 100076)(2 中国人民大学环境学院,北京 100081)(3 圈养野生动物技术北京市重点实验室,北京动物园,北京 100044)
在野外生境中,动物通过竞争获得有限的资源,如配偶、食物、水源、卧息地(巢穴)、活动及隐蔽场所等,逐渐形成了相对稳定的社群秩序(于贵瑞等,2021)。等级序位(dominance hierarchy)的建立和维持,不仅可使社群内等级较高的个体优先获得或者独占资源,降低同类侵犯、减少能耗和伤害(Clutton-Brocket al.,2006);还可使序位较低的个体逐渐改变策略,如采取回避、仪式化展示(Mattiangeliet al.,1999)等,较少发生高强度的追逐打斗等(李春旺等,2003;Clutton-Brocket al.,2006)。不同序位的动物通过改变集聚和调整标记行为、睾酮与雌二醇分泌变化、配偶选择等方式应对群居和等级的压力(Grahamet al.,2009)。麋鹿(Elaphurus davidianus)栖息于水、陆、湿地三大生境,生活史不同阶段形成了不同的稳定社群结构(张付贤等,2022),如幼龄麋鹿群可通过不断模仿、学习采取不同的应对策略适应环境变化(李春旺等,2003;张付贤等,2022),这个过程在麋鹿群体中等级始现;在亚成体阶段,种内个体间应对不同的生活压力,逐渐形成“隐形”的序位(Sih and Watters,2005;张付贤等,2022);成年阶段,雄性麋鹿发情期激烈竞争交配资源,形成“金字塔”式等级序位(Wemmer and Watling, 1986;张付贤等,2022),分为群主、挑战者和单身汉(Liet al.,2007)。
动物在生活史的不同阶段,因种内和种间资源竞争及栖息地等的不同,形成多样的性格(personali‐ty) (Gosling, 2001)、脾 气(temperament) (Réaleet al.,2007)、习性和应对策略(coping style)(Koolhaaset al.,2007);有些研究为了避免动物行为拟人化的描述,如个性等,进而采用行为型(behavioral type,BTs) 来统一替代,并在此基础上提出行为综合征(behavioral syndrome,BS)理论,即用两个以上的跨时空行为型(个性)耦合演化来解释一些生态现象和生态问题(Sihet al.,2003,2004,2012;Sih and Wat‐ters,2005)。国内近年来开展了灵长类个性、东北虎(Panthera tigris altaica)个性(苏彦捷,2007)、警犬(Canis lupus familiaris)个性测量(丁晓麟等,2013)、圈养马麝(Moschus chrysogaster)个性偶联演化(孟庆辉,2014)、林麝(Moschus berezovskii) 胆量(Yanget al.,2021)、麋鹿胆量测定及马麝行为型格局与泌香量(孟庆辉,2014;周杨等,2018a,2018b)等的相关研究。尽管对动物胆量的相关研究进展很快,但目前尚未见到个性行为偶联演替及其生态意义的相关报道,本研究从麋鹿胆量和侵犯耦合的角度,对等级序位变化机制进行初步探讨。
针对这一问题,本文首先尝试规范麋鹿个体间胆量的测定,并对麋鹿生活史不同阶段等级序位形成与波动过程中,胆量和侵犯耦合的变化开展相关测试,并依此来分析麋鹿个体从幼年到成年序位跨越的策略,其结果对动物种群结构演替及麋鹿保护提供理论帮助。
1 研究方法
1.1 实验动物
选 取11 头 出 生 于2016 年3 月25 日 至4 月4 日的雄性麋鹿。11 头麋鹿同父、同饲养条件;按照出生时间佩戴耳标以阿拉伯数字进行编号。
1.2 行为型数据搜集
1.2.1 取样时间
2016年4—12月,2017—2019年1—5月,2020年1—9月每周一的09:00—11:00和14:00—16:00,持续追踪记录。本文年龄划分依据李春旺等(2000),划分为幼体:出生后一周至角柄发育之前,时间为2016年4月—2017年12月;亚成体:角柄生长至性成熟之前,时间为2018年1月—2020年5月;成体:性成熟后,2020年6—9月首次发情。
1.2.2 行为流记录
行为流(behaviour flow)指行为的跨时空流动属性(Sihet al.,2003;孟秀祥等,2006),本文对行为的跨时空流动特性进行赋值、编码与量化,记录行为发生的时间、地点、起因、经过和结果;分析行为型的有限多样化、行为型的可塑性、行为刚性与弹性、行为惯性及行为溢出等(孟庆辉,2014)。
1.2.3 交互行为记录
交互行为(interactive behavior)指行为交流与接收个体间所做出的状态响应(Sihet al.,2004;孟庆辉,2014)。本文以焦点行为取样(Focal sampling)和扫描行为取样(Scan sampling)法进行行为观察。主要记录行为发出者、行为动机、行为指向对象、接收者的行为判别、行为响应及持续时间等。所有观察由一人完成,以5 min不间断为一个行为取样单元,若个体走出视线,则取样中断(孟秀祥等,2006)。
1.3 粪样激素检测
每周一进行行为观察的同时对每头实验动物采集粪便样品(望远镜观察和肉眼观察相结合,早、晚各采集1次,每次采集10 g以上),将每份样品分成3小份(各0.5 g)作为平行样,首先用福尔马林进行固定,然后加入90%乙醇10 mL,震荡混匀1 min,60℃水浴30 min,3 000 r/min 离心15 min,取上清液,加热至蒸干,加入1.5 mL 40%甲醇混匀,采用放射免疫试剂盒(北京北方生物技术研究所)进行粪样睾酮测定,用γ-计数器测定各样品沉淀的放射性计数,绘制剂量反应曲线,据此获得待测浓度,计算粪样激素含量(李春旺等,2000)。
1.4 胆量、侵犯与等级序位判定
本文对11 头雄性麋鹿分别进行追踪,依幼体、亚成体、成体进行计算和行为型判定。
1.4.1 行为多样性指数
计算样本行为流的多样性指数H (absolute di‐versity index)(孟秀祥等,2006)。计算公式:
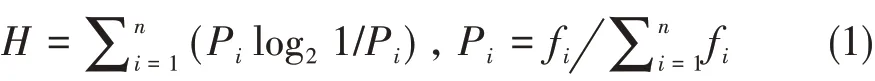
式中fi为样本行为流中第i种行为的发生频次,f为每次取样观察记录的总频次。将个体的多样性指数区分为低行为多样性组(R < 0.3)、中行为多样性组(0.3 ≤R ≤0.4)和高行为多样性组(R>0.4)。
1.4.2 刻板行为发生频率
刻板行为在总行为数据中的发生频次及持续时间,表示刻板行为的表达频率(孟秀祥等,2006)。计算公式:
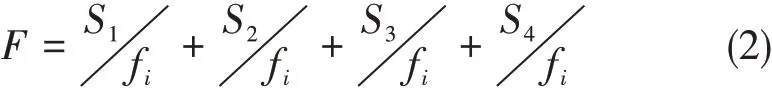
式中S1为刻板行为漫无目的地取食异物,S2为往返走,S3为狂奔,S4为凝视;fi为样本行为流中第i种行为的发生频次,f为每次取样观察记录的总频次。
1.4.3 非同步行为指数
指个体间的行为样本流的非同步化水平(孟秀祥等,2006)。计算公式:

式中Pi是行为同步指数,表示个体间行为频次和持续时间的协调一致性。
1.4.4 前摄行为指数
指样本行为流中前摄行为概率(孟秀祥等,2006)。计算公式:

式中Pi是动物自主启动的、稳定的主动适应环境变化,为试图改变自身处境,有意图性的行为;fi为样本行为流中第i种行为的发生频次,f为每次取样观察记录的总频次。
1.4.5 胆量的判定
本文在个体行为充分表达的基础上,将非同步行为中较早发生的判断为胆大(孟庆辉,2014)。行为充分表达的判断依据是:刻板行为频率较少,且行为多样性指数较高。具体方法如下:
(1)计算个体生活史阶段的指数:包括行为多样性指数、刻板行为发生频率、非同步行为指数和行为前摄指数;
(2)排序筛选刻板行为小于1%,行为多样性指数大于50%,非同步行为指数大于50%,前摄行为大于50%的个体;
(3)对行为流样本进行主成分分析,前20%的个体判定为胆大型个体,后20%的个体判断为胆小型个体,其余60%为中间型个体;
(4) 将行为多样性指数、刻板行为发生频率、非同步行为指数、行为前摄指数4 个指标求均值,换算个体的胆量指数。
1.4.6 侵犯的判定
在行为充分表达的基础上,结合攻击行为指数和亲和行为指数进行侵犯的判定(孟庆辉,2014)。具体方法如下:
(1) 计算每头动物的每个生活史阶段的指数:包括行为多样性指数、刻板行为发生频率、攻击行为指数和亲和行为指数4个指标;
(2)排序筛选刻板行为小于1%,行为多样性指数大于50%,攻击行为指数大于50%,亲和行为大于50%的个体;
(3)对行为流样本进行主成分分析,前20%的个体判定为侵犯个体,后20%的个体判断为亲和型个体,其余60%为中间型个体;
(4) 将行为多样性指数、刻板行为发生频率、攻击行为指数和亲和行为指数4个指标求均值,换算个体的侵犯指数。
1.4.7 个体等级序位的判定
依据实验动物之间发生防御、进攻、追击、威胁、取代、亲和时的总冲突频次和胜败次数,指数越大等级序位越高(孟庆辉,2014)。计算公式:
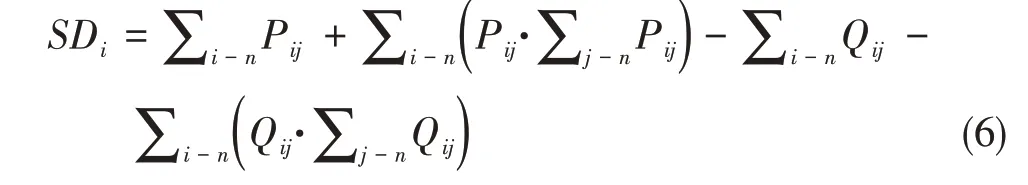
式中SDi指个体i的等级序位指数,Pij=n/N,Qij=1 −Pij,n代表个体i战胜j的次数,N代表i和j冲突的总频次。指数越大,代表等级序位越高。
1.5 数据统计及分析方法
采用Kolmogorov−Smirnov 检验数据的正态性,结合t检验分析睾酮水平与等级序位之间的关系;若数据呈非正态分布,则采用Mann WhitneyUtest检验两者间的关系。采用Spearman 相关分析检测胆量与侵犯耦合的相关性;显著性水平设定为P=0.05。对行为型数据,先计算行为多样性指数,进行泊松排布,然后结合主成分分析进行个性判定;所有数据分析均在SPSS 25.0进行。
2 结果
本研究的11 头麋鹿中,1 头成为鹿王,并成功交配(怀孕母鹿已于2021 年产仔),2 头为挑战者,8 头为单身汉(本研究数据截止2020 年9 月)。行为型判定结果:幼体麋鹿判定为胆大型个体3头、胆小型个体3 头、中间型个体5 头;幼体麋鹿判定为侵犯个体3 头、亲和型个体3 头、中间型个体5 头。亚成体与成体麋鹿,采用相同的方法进行判定。
2.1 雄性麋鹿不同年龄段等级波动
麋鹿幼体处于等级序位的发育阶段,等级尚不明显(r= 0.027 5,P= 0.005 6);亚成体麋鹿等级序位逐渐开始形成(r= 0.434 3,P= 0.630 0);到了发情期,成体麋鹿通过打斗等形成了3个明显的等级(r= 0.864 4,P= 0.008 7):分别是鹿王(5#)、挑战者(1#、6#)、单身汉(2#、3#、4#、7#、8#、9#、10#、11#)(图1)。

图1 雄性麋鹿不同年龄段等级序位形成. 幼体麋鹿1#~11#个体间未形成明显的等级;亚成体麋鹿1#、5#、6#等级序位呈上升状态;成年发情期麋鹿5#等级序位继续上升,成为鹿王Fig. 1 The rank development of male Père David’s Deer in different age groups. No rank was formed among the young 1#-11#;The rank of 1#,5#and 6#in sub⁃adult became rank increased;Adult 5#order continues to rise,then become the deer king
从幼体到亚成体麋鹿等级波动:5#、6#、7#、8#分别上升了2 个序位(胆量和侵犯耦合相关系数r=0.720 1,P=0.012 9),3#、9#、10#分别上升了1 个序位(胆量和侵犯耦合相关系数r= 0.460 1,P= 0.001 7),1#、11#等级序位没有发生改变(胆量和侵犯耦合相关系数r= 0.080 3,P= 0.005 4),2#、4#分别下降9 个和4 个等级序位(胆量和侵犯耦合相关系数r=−0.596 4,P=0.006 6)。从亚成体到成体麋鹿等级波动:4 头雄性麋鹿等级序位上升(胆量和侵犯耦合相关系数r=0.852 3,P=0.000 3),5#、6#分别上升了2 个序位(胆量和侵犯耦合相关系数r= 0.921 5,P= 0.011 2),5#个体成为了鹿王,4#、7#分别上升了1个序位(胆量和侵犯耦合相关系数r= 0.791 1,P= 0.008 1);2#、9#、10#、11#等级序位则未发生改变(胆量和侵犯耦合相关系数r= 0.482 9,P= 0.006 3);1#、3#、8#分别下降了2 个、3 个和1 个等级序位(胆量和侵犯耦合相关系数r=0.251 7,P=0.003 5)(图2)。

图2 雄性麋鹿不同年龄段等级波动过程Fig. 2 The rank fluctuation of male Père David’s Deer in different age groups
2.2 雄性麋鹿胆量和侵犯耦合程度
在麋鹿不同年龄段,面临的资源压力不同,胆量和侵犯耦合程度不同,尤其是发情期麋鹿胆量和侵犯发生了较强的耦合,并伴随着等级序位的变化(图2)。幼体:胆量与侵犯耦合r= 0.461 3,P=0.039 1;亚成体:r= 0.120 6,P= 0.0424 1;成体:r=0.930 7,P=0.003 3(图3)。

图3 雄性麋鹿不同年龄段胆量和侵犯耦合指数Fig. 3 Boldness and aggression correlation of male Père David’s Deer in different age groups
幼体麋鹿胆量与等级序位呈正相关(r=0.207 7,P=0.038 6);侵犯与等级序位呈负相关(r=−0.457 9,P=0.026 4);胆量和侵犯耦合与等级序位呈负相关(r=−0.111 8,P=0.018 3)(表1)。粪样睾酮水平与等级序位呈正相关(r=0.860 7,P=0.005 5)。
亚成体麋鹿胆量与等级序位呈正相关(r=0.376 7,P= 0.041 1);侵犯与等级序位呈正相关(r=0.442 8,P=0.037 4);胆量和侵犯耦合与等级序位呈正相关(r= 0.414 7,P= 0.028 9)。粪样睾酮水平与等级序位呈正相关(r= 0.845 7,P=0.004 4)(表1)。
成体麋鹿胆量与等级序位呈正相关(r=0.941 5,P= 0.002 1);侵犯与等级序位呈正相关(r= 0.818 3,P= 0.007 2);胆量和侵犯耦合与等级序位呈正相关(r= 0.917 9,P= 0.002 6)。粪样睾酮水平与等级序位呈正相关(r= 0.954 6,P= 0.001 8)(表1)。

表1 麋鹿胆量、侵犯与等级序位发育Table 1 Relationship between boldness,aggression and rank development in male Père David’s Deer

续表1 Continued from table 1
3 讨论
本文对11 头雄性麋鹿,从幼体到成体进行持续追踪观察,有4头麋鹿等级序位持续跃升,可能是通过胆量与侵犯耦合的策略实现;2 头麋鹿从幼体到成体发生等级序位持续下降,可能跟其母亲死亡较早有关,其中1 头麋鹿2018 年冬生病1 次,也可能与其侵犯行为和体重增速不匹配有关。
3.1 胆量和侵犯耦合与等级之间的关系分析
幼体麋鹿胆量和侵犯耦合与等级序位发育呈负相关,可能因幼体麋鹿的侵犯较强对等级序位发育不利。亚成体麋鹿胆量和侵犯耦合促进等级序位的定型,此期等级序位已基本形成,但保持着较低的侵犯。发情期麋鹿胆量和侵犯耦合与等级序位的强烈波动呈正相关;对于季节性繁殖的鹿类,发情季节睾酮分泌显著增加,可驱动胆量和侵犯发生较强耦合,促使麋鹿的等级序位发生较大波动。这与其他鹿类动物的研究结果一致,如圈养的幼体獐(Hydropotes inermis) 序位与侵犯频率呈负相关(Maugetet al., 2007);泌香期间,圈养马麝(Moschus chrysogaster) 也保持较低的侵犯(周杨等,2018b)。
3.2 不同个体等级波动分析
从幼体到亚成体麋鹿等级序位波动情况为:3#、5#、6#、7#、8#、9#、10#等级 序 位上升;2#、4#等级序位下降;1#、11#等级序位没有发生改变。从亚成体到成体麋鹿等级序位波动情况为:5#、6#、4#、7#等级序位持续上升,其中5#个体成为了鹿王;2#、9#、10#、11#等级序位则未发生改变;1#、3#、8#等级序位发生了下降。由此可以看出,胆量和侵犯耦合紧密时促进麋鹿等级序位的上升,两者耦合松散时等级序位几乎没有变化,两者解耦合时伴随着麋鹿等级序位的下降。这与其他灵长类动物研究结果相吻合,如黑脸长尾猴(Cercopithecus aethiops)未成年等级序位上升与胆量呈正相关(苏彦捷,2007);雄性黄山藏酋猴(Macaca thibetana) 交配期序位下降与强侵犯相关(陈燃等,2008)等。
3.3 粪样睾酮水平与等级序位的关系
动物的等级序位和社群冲突与性激素水平密切相关。本研究结果显示,幼体麋鹿粪样睾酮水平与等级发育呈正相关;亚成体麋鹿粪样睾酮水平与序位形成呈正相关;成体麋鹿粪样睾酮水平与序位波动间呈正相关。因此,麋鹿等级序位的发育、定型及波动与粪样睾酮水平呈正相关,由于动物体内睾酮水平的波动,导致胆量和侵犯耦合程度变化,直接影响动物等级序位的升降。这与其他动物研究结果一致,如非洲黑脸长尾猴等级序位高低与雄激素水平呈正相关(苏彦捷,2007);圈养獐侵犯频率与雄激素分泌呈正相关(Maugetet al., 2007);雄性蹄兔(Procavia capen‐sis)、雌性山羊(Capra hircus)和圈养八齿鼠(Abro‐comidae) 序位和侵犯冲突强度与睾酮水平正相关(Koren and Geffen, 2009);雄性黄山藏酋猴交配期粪睾酮水平与攻击频次正相关(陈燃等,2008)等。
麋鹿在不同年龄段等级序位发生不断的变化,尤其是争夺交配资源时,序位波动尤为显著(张付贤等,2022)。本文尝试用行为型耦合来解释等级序位变化,既丰富了等级序位变化的理论,又探讨了序位波动的机制,为鹿科动物社群和社会关系网络研究提供了思路和可靠依据。
