马可·波罗盘羊和5种家养绵羊MHC-DRB3基因第2外显子遗传多态性分析
2022-03-17张永浩郭丽君阿布都热合曼吐尔逊王玉涛
张永浩,肖 逸,郭丽君,阿布都热合曼·吐尔逊,王玉涛
(喀什大学生命与地理科学学院,新疆帕米尔高原生物资源与生态重点实验室,喀什 844000)
主要组织相容性复合体(major histocompatibility complex,MHC)是参与细胞表面转膜蛋白的主要基因,其通过传递抗原给T淋巴细胞来激发免疫反应,是基因组中多态性最为丰富的区域[1-3],其产物MHC抗原在脊椎动物的免疫调节过程中发挥着重要功能[4-7]。1906年,Tyzzer等进行了有关小鼠肿瘤移植的试验,是对MHC的初步探索[8];1956年,Snell等在移植小鼠皮肤时产生了供体组织与受体组织的排斥反应,他率先对组织相容性基因进行了定义[9]。小鼠MHC存在于H-2系统的17号染色体上,而人MHC简称为HLA(human leukocyte antigen),位于6号染色体上[10]。大多数脊椎动物体内都发现了MHC系统,且不同种类动物的组成、结构和功能有较多相似的地方[11]。为便于区分,采用不同编码的抗原系统来命名不同种属动物的MHC,最早用血型系统来命名的是小鼠和鸡[12],该命名方法延用至今,对其他动物MHC的命名方式是保留该动物英文全称的第一个字母,再在第一个字母后加上LA,这样不同动物的MHC系统都有了相应的简写,如人、大猩猩、犬、猪、马、牛、绵羊、山羊和兔的MHC系统都是通过这种简写得到的[13-18]。
马可·波罗盘羊分布于中国、巴基斯坦、阿富汗、塔吉克斯坦和吉尔吉斯斯坦五国接壤的边境区域,并被列为一级保护动物[19]。王玉涛等[20]调查发现,分布于东帕米尔高原的马可·波罗盘羊的种群数量为3 000~3 500只;龚明昊等[21]研究表明,新疆塔什库尔干野生动物自然保护区马可·波罗盘羊的种群数量为1 500~1 700只[22],该保护区是中国以马可·波罗盘羊为首要保护对象的最重要的自然保护区[23]。近几年,关于MHC基因多态性的研究多集中在家畜中,有关珍稀濒危动物的MHC研究相对较少[24-27],而关于马可·波罗盘羊MHC基因多态性的研究鲜见报道。绵羊MHC由Ⅰ、Ⅱ和Ⅲ类区域组成,其中在Ⅱ类区域的DQ和DR亚区多态性较为集中,其外显子2为功能区,因此,MHC的研究主要集中在DRB3基因外显子2上[28]。本研究采用PCR-SSCP和克隆测序技术对马可·波罗盘羊和5种家绵羊MHC-DRB3基因第2外显子的多态性和系统发育进行分析,确定马可·波罗盘羊和5种家绵羊MHC-DRB3基因第2外显子的等位基因数及核苷酸、氨基酸的多态位点,并对各等位基因的遗传关系和进化意义进行了初步探讨,以期为进一步探究绵羊MHC-DRB3基因的调控机理及进化关系提供参考。
1 材料与方法
1.1 样品采集
采集6种绵羊样本共408份,其中马可·波罗盘羊肌肉样本采自帕米尔高原塔什库尔干县的卡拉其古、野生动物自然保护区管理局、红其拉甫、萨热吉力嘎、克孜勒苏柯尔克孜自治州阿克陶县木吉乡以及喀什市动物园等地,共39份;藏绵羊血液样本采自甘肃玛曲县,共72份;宁夏滩羊血液样本采自宁夏盐池县,共76份;柯尔克孜羊血液样本采自新疆喀什市,共36份;塔什库尔干羊血液样本采自新疆塔什库尔干县,共159份;多浪羊血液样本采自新疆麦盖提县,共26份。以上样本均-80 ℃保存于新疆帕米尔高原生物资源与生态重点实验室。
1.2 主要试剂及仪器
Tris饱和酚、蛋白酶K、DNA Marker、2×TaqPCR Mix、2×DNA Loading Buffer和质粒小提试剂盒均购自中科瑞泰(北京)生物科技有限公司;大肠杆菌DH5α感受态细胞购自山东宝信生物科技有限公司。
冰冻离心机、立式超低温保存箱和PCR仪均购自Eppendorf公司;冷藏箱购自青岛海尔特种电器有限公司;电泳仪购自北京六一仪器厂;垂直电泳槽购自Bio-Rad公司;电热恒温培养箱购自上海一恒科学仪器有限公司;超净工作台购自苏州安泰空气技术有限公司。
1.3 PCR扩增及PCR-SSCP分型
参考Amills等[29]设计引物序列,上游引物:5′-TATCCCGTCTCTGCAGCACATTTC-3′;下游引物为:5′-TCGCCGCTGCACACTGAAACTCTC-3′,预计扩增产物长度为285 bp。引物由金唯智生物科技有限公司合成。
盘羊基因组DNA提取方法参照《分子克隆实验指南》(第4版),采用常规的酚-氯仿抽提法,并进行了优化和改进[30-31]。 PCR反应体系20 μL:2×TaqPCR Mix 12 μL,上、下游引物(10 pmol/μL)各0.4 μL,模板DNA 1 μL,ddH2O 6.4 μL。PCR扩增程序:94 ℃预变性5 min;94 ℃变性1 min,63 ℃退火1 min,72 ℃延伸1 min,共35个循环;72 ℃延伸10 min;4 ℃保存。取2.5 μL PCR产物经2.0%琼脂糖凝胶电泳检测,鉴定正确后对其进行分型。在电泳电压和电泳时间恒定时,于Arc∶Bis=29∶1、胶浓度为12%、6 ℃条件下检测,待垂直板22 h电泳结束后用简易银染法进行染色,拍照保存。对MHC-DRB3基因第2外显子不同基因型进行基因型频率分析。
1.4 克隆及测序
利用琼脂糖凝胶回收试剂盒对不同基因型个体的PCR扩增产物进行目的片段回收,回收产物连接pMD19-T克隆载体,转化大肠杆菌DH5α感受态细胞,挑选阳性菌落进行菌落PCR鉴定,对鉴定正确的阳性菌落提取质粒送金唯智生物科技有限公司测序。
1.5 生物信息学分析
测序结果通过NCBI BLAST程序比对正确后,利用DnaSP 6.12.03软件分析MHC-DRB3基因第2外显子序列的碱基组成、单倍型及氨基酸变异位点。利用Mega 6.06软件对马可·波罗盘羊MHC-DRB3基因第2外显子序列进行遗传进化分析,所用序列为本研究获得的66条绵羊的单倍型序列和1条鹅喉羚单倍型序列(E1),采用Kimura-2模型,以邻接法构建绵羊MHC-DRB3基因第2外显子序列的系统进化树,并对拓扑图进行自展检验(bootstrap),重复检验次数为1 000。利用多种在线工具分析MHC-DRB3的结构和功能,详细信息见表1。

表1 生物信息学分析软件
2 结 果
2.1 MHC-DRB3基因第2外显子PCR扩增
对马可·波罗盘羊及家养绵羊MHC-DRB3基因第2外显子进行PCR扩增,扩增产物经2.0%琼脂糖凝胶电泳检测得到一条清晰、明亮、特异性高的条带,大小为300 bp左右(图1),与预期相符,可进行PCR-SSCP检测。
2.2 MHC-DRB3基因第2外显子PCR-SSCP检测及基因型频率分析
PCR-SSCP检测结果共发现15种基因型,分别命名为A~O(图2)。15种基因型中频率最高的为H型(表2),说明H型为马可·波罗盘羊和5种家养绵羊品种的优势基因型。
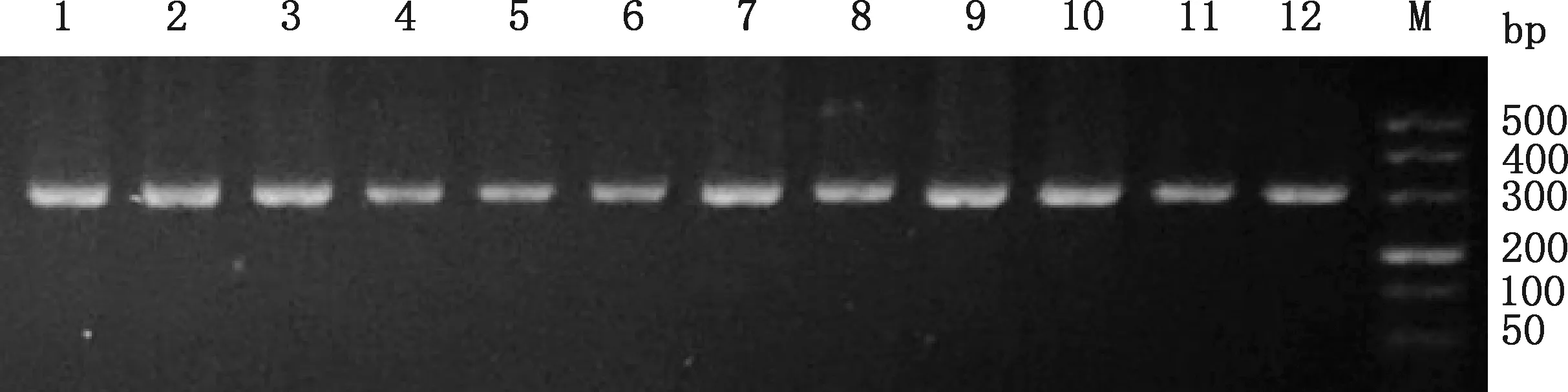
1、2,马可·波罗盘羊;3、4,塔什库尔干羊;5、6,藏绵羊;7、8,宁夏滩羊;9、10,柯尔克孜羊;11、12,多浪羊;M,DNA Marker1 and 2,Ovis ammon polii;3 and 4,Taxkorgan sheep;5 and 6,Tibetan sheep;7 and 8,Ningxia Tan sheep;9 and 10,Kirgiz sheep;11 and 12,Duolang sheep;M,DNA Marker图1 MHC-DRB3基因第2外显子PCR扩增产物Fig.1 PCR amplification products of MHC-DRB3 gene exon 2
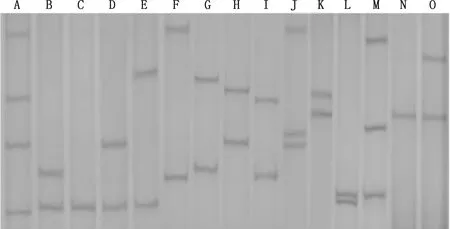
图2 MHC-DRB3基因第2外显子PCR-SSCP检测Fig.2 PCR-SSCP detection of MHC-DRB3 gene exon 2
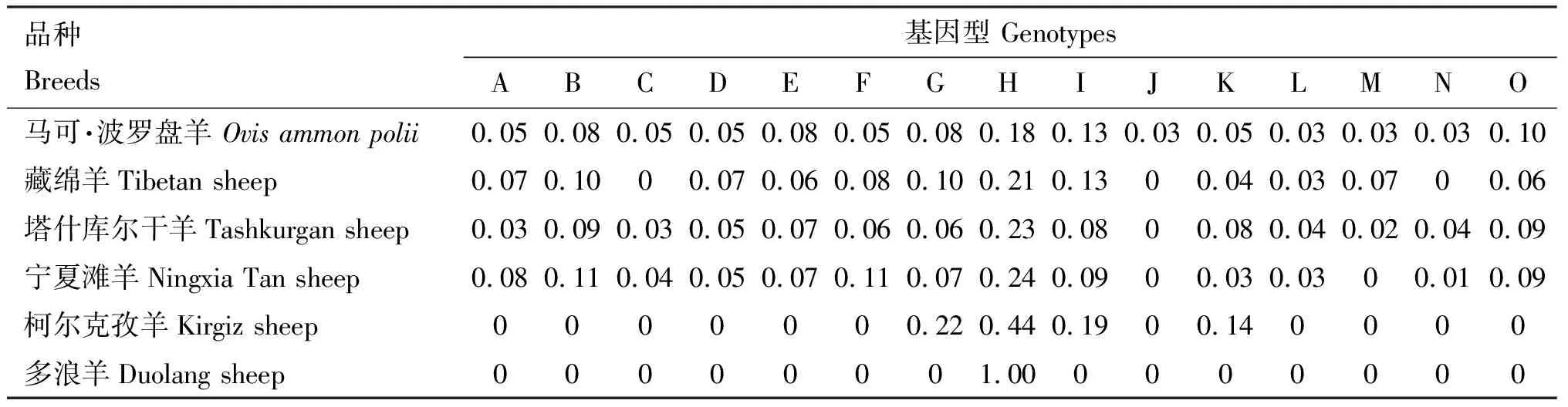
表2 马可·波罗盘羊和5种家养绵羊基因型频率统计
2.3 MHC-DRB3基因第2外显子等位基因氨基酸变异位点分析
通过Mega 6.06软件对66个等位基因进行氨基酸序列比对分析,共翻译成79个氨基酸位点,其中多态位点为55个,占氨基酸位点总数目的69.6%(图3),略高于MHC-DRB3基因第2外显子核苷酸多态位点占比(在去除引物序列的237 bp中发现107个多态位点,占比为45.1%),充分印证了DRB3基因第2外显子多态性较高。
2.4 MHC-DRB3基因第2外显子碱基组成及单倍型分析
经序列比对发现,MHC-DRB3基因第2外显子长285 bp,A、T、C和G占比分别为23.38%、16.50%、23.07%和37.05%,其中CG含量(60.12%)明显高于AT含量(39.88%),说明存在一定的碱基偏倚性。对马可·波罗盘羊和5种家养绵羊MHC-DRB3基因第2外显子的不同基因型进行克隆测序后发现66个单倍型,经序列比对得出,本研究新发现了64条等位基因序列(表3)。
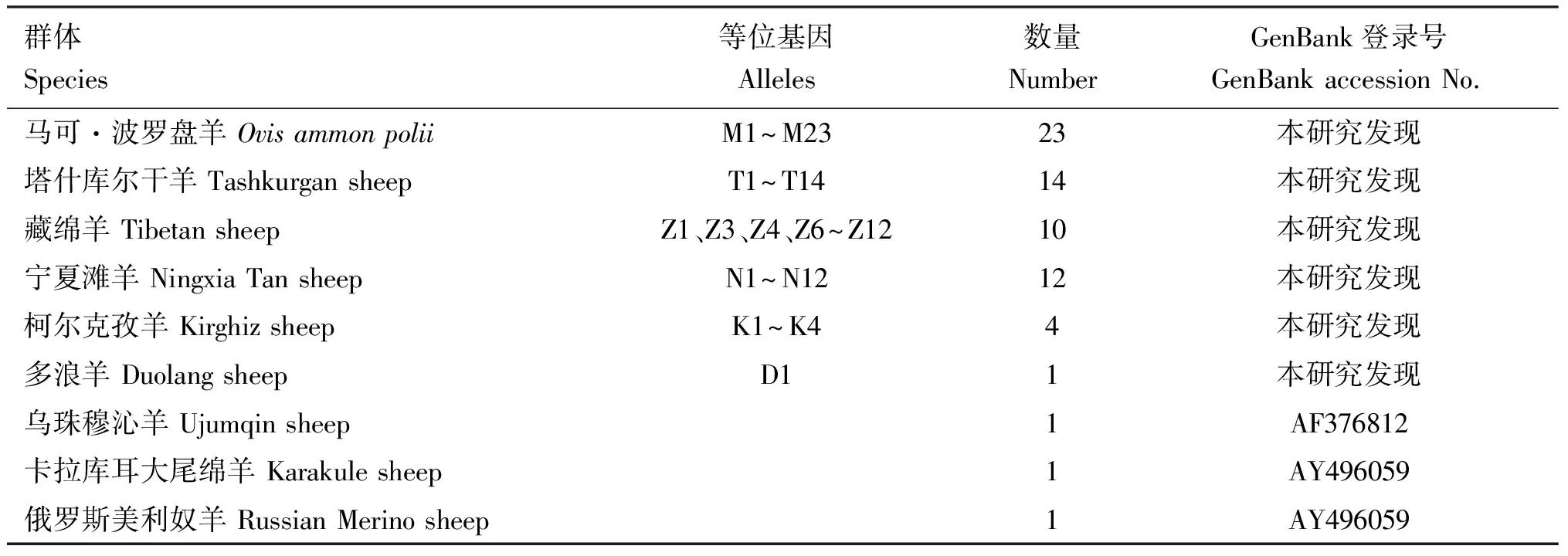
表3 绵羊MHC-DRB3基因第2外显子单倍型比较
2.5 系统进化树构建
由图4可知,马可·波罗盘羊和塔什库尔干羊MHC-DRB3基因第2外显子序列系统进化树呈现两支分化趋势。马可·波罗盘羊在Ⅰ支中有17个单倍型,在Ⅱ支中有6个单倍型;塔什库尔干羊在Ⅰ支中有12个单倍型,在Ⅱ支中有2个单倍型,马可·波罗盘羊和塔什库尔干羊Ⅰ支中的单倍型明显高于Ⅱ支中的单倍型。藏绵羊、宁夏滩羊、柯尔克孜羊和多浪羊并未出现两支分化,单倍型分别为12、12、4及1个。鹅喉羚(E1)的单倍型呈独立的分支,说明鹅喉羚的单倍型序列与马可·波罗盘羊、塔什库尔干羊、藏绵羊、宁夏滩羊、柯尔克孜羊和多浪羊6种绵羊的单倍型序列相似性较低。
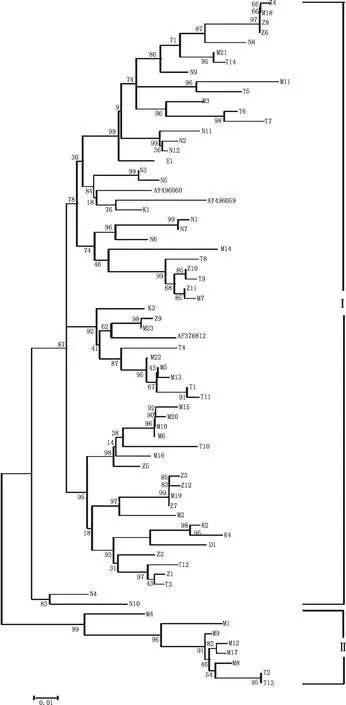
图4 马可·波罗盘羊和5种家养绵羊品种MHC-DRB3基因第2外显子系统进化树Fig.4 Phylogenetic tree based on MHC-DRB3 gene exon 2 of Ovis ammon polii and five domestic sheep breeds
2.6 MHC-DRB3基因第2外显子核苷酸多态性分析
用DnaSP 6.12.03软件对马可·波罗盘羊和家养绵羊MHC-DRB3基因第2外显子单倍型的多态位点进行分析,Pi1为马可·波罗盘羊,其余Pi2为与之参考的藏绵羊、宁夏滩羊、柯尔克孜羊、塔什库尔干羊。由图5~8可知,藏绵羊、塔什库尔干羊与马可·波罗盘羊核苷酸多样度变化趋势最为相似,宁夏滩羊、柯尔克孜羊与马可·波罗盘羊核苷酸多样度变化趋势相差较大。
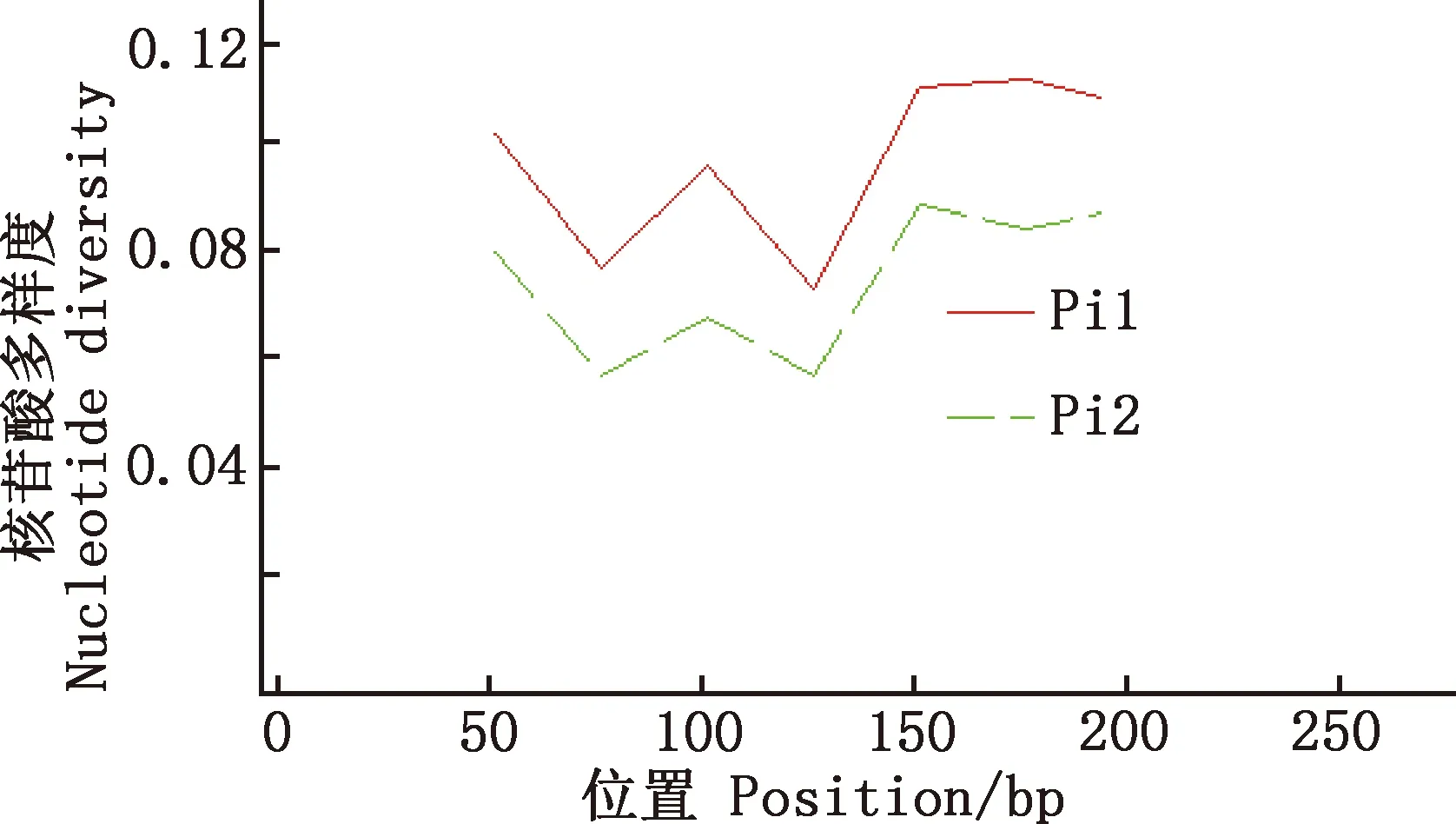
图5 马可·波罗盘羊和藏绵羊MHC-DRB3基因第2外显子核苷酸多样度分布Fig.5 Nucleotide diversity distribution of MHC-DRB3 gene exon 2 in Ovis ammon polii and Tibetan sheep
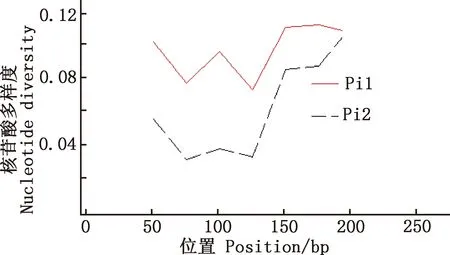
图6 马可·波罗盘羊和宁夏滩羊MHC-DRB3基因第2外显子核苷酸多样度分布Fig.6 Nucleotide diversity distribution of MHC-DRB3 gene exon 2 in Ovis ammon polii and Ningxia Tan sheep
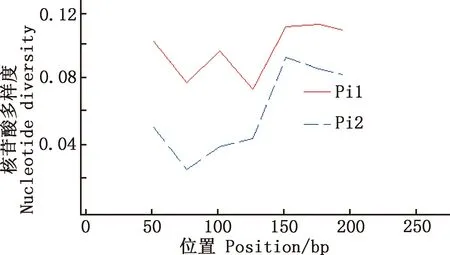
图7 马可·波罗盘羊和柯尔克孜羊MHC-DRB3基因第2外显子核苷酸多样度分布Fig.7 Nucleotide diversity distribution of MHC-DRB3 gene exon 2 in Ovis ammon polii and Kirgiz sheep
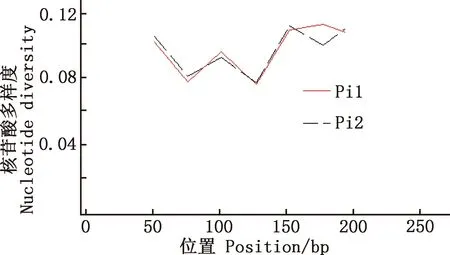
图8 马可·波罗盘羊和塔什库尔干羊MHC-DRB3基因第2外显子核苷酸多样度分布Fig.8 Nucleotide diversity distribution of MHC-DRB3 gene exon 2 in Ovis ammon polii and Tashkurgan sheep
2.7 生物信息学分析
2.7.1 理化性质预测 利用ExPASy中的在线工具ProtScale完成对马可·波罗盘羊MHC-DRB3基因第2外显子蛋白序列亲/疏水性进行性分析,结果显示,马可·波罗盘羊MHC-DRB3基因第2外显子氨基酸序列绝大多数为亲水性残基(图9),推测马可·波罗盘羊MHC-DRB3基因第2外显子蛋白为水溶性蛋白。
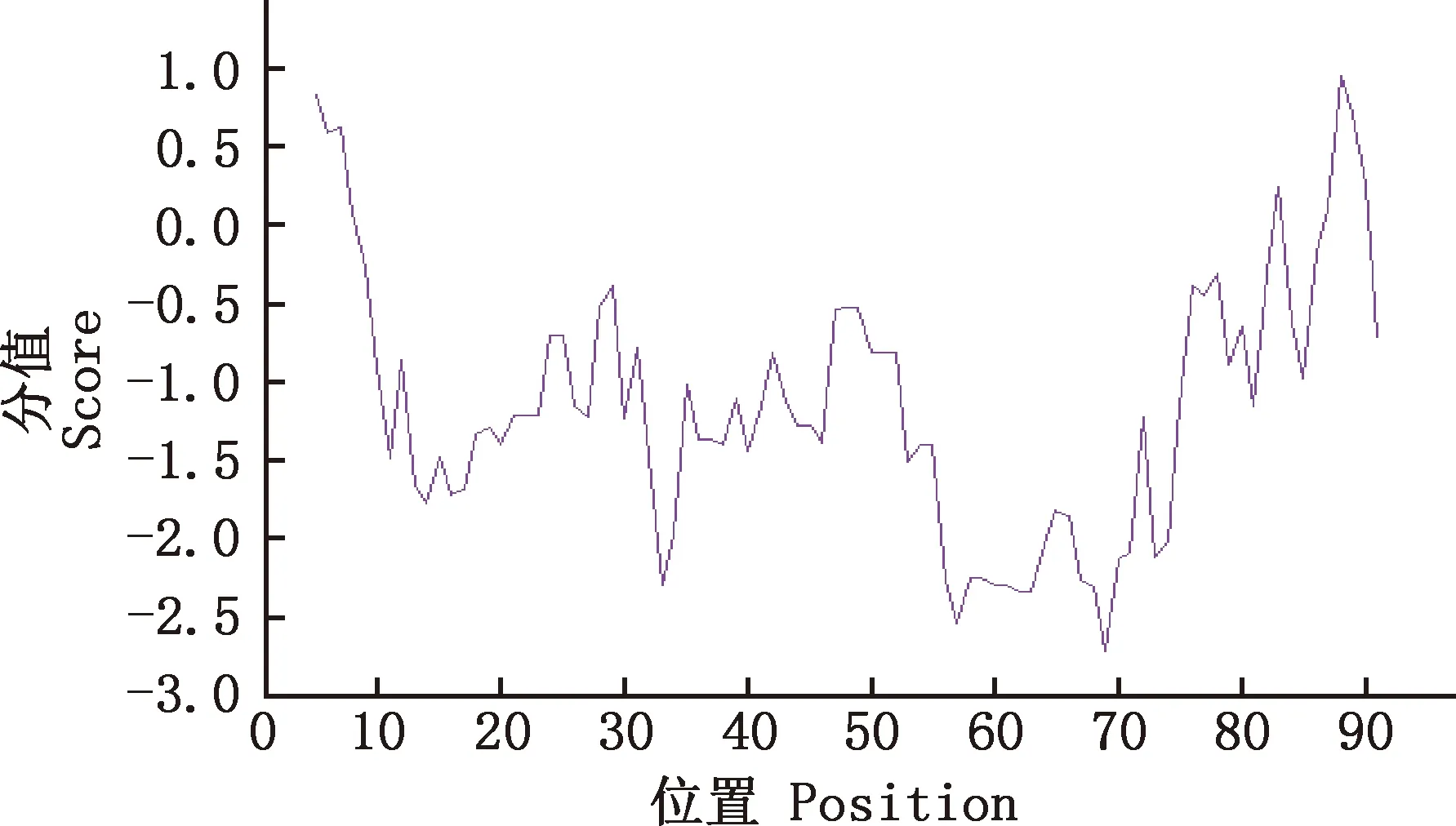
图9 马可·波罗盘羊MHC-DRB3基因第2外显子氨基酸序列亲/疏水性分析结果Fig.9 Hydrophilic and hydrophobic prediction of amino acid sequence of MHC-DRB3 gene exon 2 in Ovis ammon polii
2.7.2 跨膜区预测 通过TMPRED在线软件对马可·波罗盘羊MHC-DRB3基因第2外显子氨基酸序列进行跨膜螺旋结构预测,结果显示,马可·波罗盘羊MHC-DRB3基因第2外显子中不存在跨膜螺旋结构(图10)。
2.7.3 亚细胞定位及二级结构预测 采用Psort在线软件对马可·波罗盘羊MHC-DRB3基因第2外显子编码产物进行亚细胞定位,结果显示,马可·波罗盘羊MHC-DRB3基因第2外显子蛋白主要在细胞质、细胞核、线粒体和高尔基体中发挥生物学作用(表4)。采用SOPMA在线软件预测马可·波罗盘羊MHC-DRB3基因第2外显子编码产物的二级结构,结果显示,马可·波罗盘羊MHC-DRB3基因第2外显子编码产物由α-螺旋、延伸链、β-转角和无规则卷曲组成,占比分别为30.53%、22.11%、11.58%和35.78%(表4)。
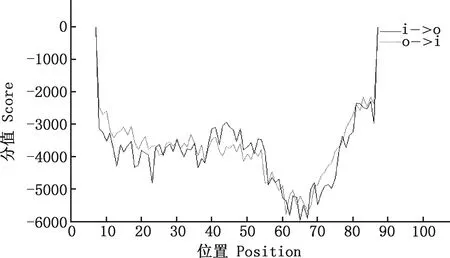
图10 马可·波罗盘羊MHC-DRB3基因第2外显子氨基酸序列跨膜区预测Fig.10 Transmembrane region prediction of amino acid sequence of MHC-DRB3 gene exon 2 in Ovis ammon polii
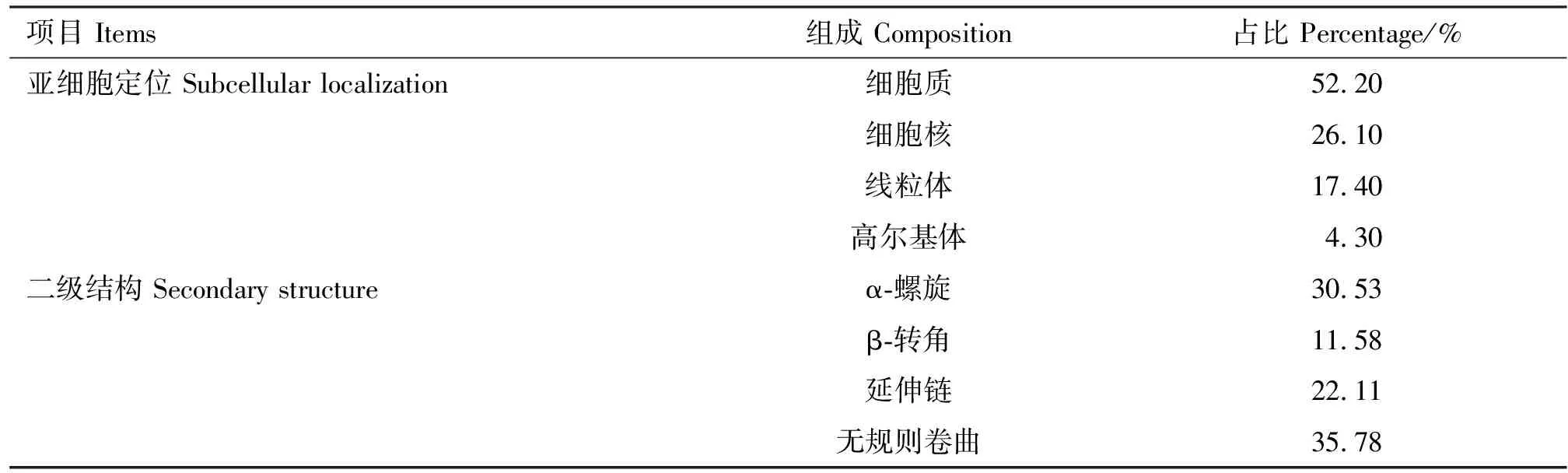
表4 马可·波罗盘羊MHC-DRB3基因第2外显子亚细胞定位及二级结构预测
3 讨 论
本研究采用PCR-SSCP技术对绵羊MHC-DRB3基因第2外显子进行检测,相较于PCR-RFLP方法,PCR-SSCP技术检测得到的等位基因数量明显较多[32]。本研究采用PCR-SSCP技术检测了408只绵羊MHC-DRB3基因第2外显子的多态性,共发现了66个等位基因,与GenBank中提交的3个等位基因都不相同,有2个等位基因与成述儒[33]发现的H14和H30等位基因相同,所以本研究共新发现了64个等位基因。39只马可·波罗盘羊中发现了23个等位基因,说明马可·波罗盘羊群体间等位基因出现的频率比其他5种家养绵羊更高,其多态性可能更高,也表明马可·波罗盘羊群体遗传多样性丰富,是弥足珍贵的遗传资源。
6种绵羊的MHC-DRB3基因第2外显子单倍型序列系统发生分析中,马可·波罗盘羊和塔什库尔干羊呈明显的两支分化,这与成述儒等[34]研究的藏绵羊MHC-DRB3基因单倍型呈两支分化的情况相似。研究表明,MHC-DRB3基因的高度多态性与绵羊适应高寒、高海拔的恶劣环境而产生的耐寒、耐粗、适应能力较强的特性密切相关[34-35]。故推测马可·波罗盘羊、藏绵羊和塔什库尔干羊MHC-DRB3基因第2外显子单倍型多态性和相似性较高且相互聚为一支的原因与这3种绵羊所处的高寒、高海拔的地理环境有关。柯尔克孜羊、多浪羊和宁夏滩羊只与各自的单倍型聚为一支,说明这3种羊的单倍型相似性较低,可能与这3种绵羊所处的地理环境差别较大有关。本研究发现,H型为马可波罗·盘羊和5种家养绵羊群体中的优势基因型,通过对优势基因型的筛选有利于后续的品种保护和遗传育种。
本试验利用生物信息学方法对马可·波罗盘羊MHC-DRB3基因第2外显子以及编码产物的理化性质、序列特征和生物学功能进行预测和分析,结果显示,马可·波罗盘羊MHC-DRB3基因第2外显子主要在细胞质中发挥生物学作用,其编码的氨基酸序列中有78个氨基酸为亲水性残基,推测马可·波罗盘羊MHC-DRB3基因第2外显子蛋白为水溶性蛋白;无规则卷曲为马可·波罗盘羊MHC-DRB3基因第2外显子中占比最大的蛋白二级结构元件,α-螺旋、延伸链和β-转角则散在分布于整个蛋白质中,与徐飞[14]、黄兰[16]研究结果相一致。表明MHC-DRB3基因多态性与绵羊对环境的适应性密切相关,如抗病性、耐寒性发生变化,绵羊的基因频率也会随之改变,但马可·波罗盘羊与5种家养绵羊MHC-DRB3基因的内在分子调控机制仍待进一步研究。
4 结 论
马可·波罗盘羊MHC-DRB3基因第2外显子共发现66个等位基因,其中64个为新发现的等位基因。马可·波罗盘羊和塔什库尔干羊MHC-DRB3基因第2外显子的多态性更为丰富;马可·波罗盘羊、塔什库尔干羊和藏绵羊MHC-DRB3基因第2外显子单倍型的相似性较高。马可·波罗盘羊MHC-DRB3基因第2外显子基因编码产物为水溶性蛋白,不存在跨膜螺旋结构,其生物学作用主要在细胞质中体现,二级结构主要以无规则卷曲和α-螺旋为主。
