甘蓝型油菜phytocyanin家族全基因组鉴定和表达分析
2022-03-16张帅林科运侯纯朴方玉洁王幼平
张帅,林科运,侯纯朴,方玉洁,王幼平
(扬州大学生物科学与技术学院,江苏 扬州,225009)
质体蓝素(phytocyanin,PC)是一类植物特异性I型蓝铜蛋白(blue copper proteins,BCP),包括plastocyanin 和PC 相关蛋白[1,2]。尽管PC 蛋白家族成员之间的序列一致性通常并不高,但该家族所有成员的蛋白质序列均具有两个保守的能形成二硫键的半胱氨酸(cysteine,Cys),其中部分成员具有4 个保守的铜离子结合位点,由2 个组氨酸(histidine,His)、1个Cys和1个甲硫氨酸(methionine,Met)或谷氨酰胺(glutamine,Gln)组成[2,3]。PC 蛋白有4 个结构域(IIV),包括1 个必要的类质体蓝素域(plastocyaninlike domain,PLCD)(结构域II)和3 个可选结构域(结构域I、III和IV)[4]。
根据铜配体残基的特性和蛋白结构组分等特征,PC 家族可分为4 个亚家族:类质体蓝素蛋白(plantacyanin-like protein,PLCL)、类花青素苷蛋白(uclacyanin-like protein,UCL)、类漆树蓝蛋白(stellacyanin-like protein,SCL)和类早期结瘤素蛋白(early nodulin-like protein,ENODL)[5,6]。其中UCL与PLCL 的铜配体残基相同,由2个His、1个Cys和1个Met 组成,而UCL 则属于嵌合型糖蛋白,PLCL 为非糖蛋白[7]。与UCL 和PLCL 不同,SCL 铜配体残基中的Gln被Met所取代,但SCL与作为嵌合型糖蛋白的UCL 相似,它们不仅包含铜结合结构域,而且其蛋白质骨架中还包含糖蛋白样结构域,SCL 和UCL通过Asn-X-Thr/Ser 中的Asn 残基提供N-糖基化位点,部分SCL 和UCL 通过Ser 和羟脯氨酸(Hyp)残基提供O-糖基化位点[8]。与另外三个亚家族不同,ENODL 家族成员的蛋白骨架缺乏完整的铜离子结合位点,铜配体中的4 个氨基酸残基部分或全部被其它氨基酸残基取代[9]。研究发现,在拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)中,大部分PC中存在类阿拉伯半乳聚糖蛋白域(arabinogalactan protein-like region,ALR),因此PC基因家族也被归为AGP超家族中的一个亚家族[9,10]。
在已有的研究报道中,PLCL、SCL、UCL 蛋白功能研究较少。在拟南芥中,过表达plantacyanin抑制花粉粒萌发,破坏花粉管的定向生长,并导致种子结实率降低[11]。在辣椒中,SCL 参与了被野油菜黄单胞菌叶斑病无毒菌株感染的叶片的超敏反应[12]。过表达OsUCL8导致花粉管的生长和授粉出现明显异常,影响水稻结实率;相反,敲除OsUCL8和过表达miR408 的植株花粉萌发率显著提高[13]。与功能研究较少的PLCL、SCL、UCL 相比,ENODL 蛋白的功能研究更为广泛和全面,且在根中的研究居多。豆科植物中的研究表明,大豆中的GmENODL55[14],豌豆中的VsENODL5、VsENODL12和VsENODL40[15],以及苜蓿中的MtENODL16和MtENODL20[16]在根瘤中高表达,参与了豆科植物的结瘤过程。已有研究证实,VsENOD5在受根瘤菌侵染前的根瘤原基中并不表达,而在受侵染后携带侵染线的豌豆根皮层细胞和被侵染的根瘤细胞中表达[15,17]。大豆GmENOD55也显示出相似的表达模式,仅在受慢生型大豆根瘤菌侵染后的大豆根瘤细胞中表达[14]。MtENOD20是一个在根皮层细胞激活过程中受结瘤因子诱导的标志基因,该基因的转录激活起始于对应根瘤原基形成位置处于分裂状态的内皮层细胞,随后该基因在含有侵染线的根毛细胞中表达[18]。最近的研究显示,蒺藜苜蓿MtENODL27和MtENODL28 在根瘤中特异表达,且在被侵染的根中表达上调;抑制MtENODL27和MtENODL28表达会阻碍根瘤菌的侵染和根瘤形成,说明MtENODL27/28 是根瘤菌侵染和根瘤发育的必需因子[4]。除了在根系中表达之外,ENODL基因也被发现在水稻的高蛋白淀粉层中表达[19]。ENODL家族中的Pn14基因在牵牛花(Pharbitis nil)的分生组织、原形成层、茎尖周围的叶原基表皮毛以及根尖的分生组织和原形成层中表达,暗示其可能参与植物的细胞壁重建和器官分化[20]。此外,拟南芥中At3g20570编码的ENODL 蛋白在筛管分子的细胞膜中积聚,并参与调控拟南芥的生殖能力[21]。这些研究表明ENODL 不仅在根中发挥作用,而且可能在植物发育的其它阶段发挥功能。
前人的研究表明,PC也参与植物对非生物胁迫的应答过程。厚叶蛛毛苣薹(Boea crassifolia)中BcBCP1受干旱胁迫和盐胁迫诱导上调表达,在烟草中异源过表达BcBCP1使转基因烟草的抗旱性增强,在渗透胁迫下表现出更高的存活率和更高的光合速率[22]。玉米中的PC基因家族也参与对盐胁迫和干旱胁迫的响应,在盐处理条件下,玉米中9 个PC基因(ZmUC10、ZmUC16、ZmUC19、ZmSC2、ZmUC21、ZmENODL10、ZmUC22、ZmENODL13和ZmENODL15)表达下调;在干旱处理条件下,5个PC基因(ZmUC19、ZmSC2、ZmENODL10、ZmUC22和ZmENODL13)的表达下调,而ZmUC16表达显著增强[23]。早期的研究证实,拟南芥蓝铜结合蛋白基因AtBCB的表达受光负调节,其编码的蓝铜结合蛋白可能作为电子载体参与细胞膜区的电子传递反应[24]。AtBCB/AtSC3受铝胁迫和氧化胁迫诱导表达[25,26]。后续研究表明,AtBCB蛋白主要定位于细胞膜上,与野生型植株相比,过表达AtBCB的转基因拟南芥在根中积累更多的木质素,过表达AtBCB的酵母转化子对铝的抗性增强,说明AtBCB 能够抑制铝的吸收和铝毒害造成的氧化胁迫[27]。
全基因组系统分析能够为鉴定基因家族成员并阐明其生物学作用提供有效的途径和依据。目前研究者们已经对拟南芥[10]、水稻[9]、大白菜(Brassica rapa)[28]、玉米(Zea mays)[23]、蝴蝶兰(Phalaenopsis equestris)[29]和蒺藜苜蓿(Medicago truncatula)[4]中的PC 家族基因进行了全基因组鉴定和分析。甘蓝型油菜(Brassica napus)是一种由白菜和甘蓝自然杂交形成的异源四倍体植物[30],作为集食用、饲用和观赏价值于一身的常见农作物,该物种中PC基因家族的功能尚不明确。本研究对甘蓝型油菜PC基因(BnPC)家族进行了全基因组范围内的鉴定、生物信息学分析和表达分析,旨在了解BnPC基因家族成员的组成、分类和表达特征,为揭示该家族成员的生物学功能提供线索。
1 材料与方法
1.1 BnPC家族成员的鉴定
本研究利用HMMER 软件和PLCD 的HMM 在甘蓝型油菜全基因组范围内对PC 家族成员进行鉴定,并进一步利用SMART 和UniProt 数据库对这些潜在的PC 蛋白进行PLCD 结构域的确认。在Pfam 33.1 数据库(http://pfam. xfam. org/)中利用关键词“phytocyanin”和“plastocyanin”进行搜索,并利用PLCD 的隐马尔可夫模型(Hidden-Markov Model,HMM)PF02298(Plastocyanin-like domain)和HMMER 软件对甘蓝型油菜数据库(http://www. genoscope.cns.fr/brassicanapus/)全基因组蛋白序列进行检索(E-value ≤1e-5),获得潜在的BnPC 家族成员信息。进一步利用SMART(http://smart. embl-heidelberg. de)和UniProt(http://www. uniprot. org/)数据库对获得的假定PC 成员的PLCD 结构域进行鉴定,筛选确认具有完整PLCD 结构域的序列,即为BnPC家族成员。
1.2 BnPC 家族基因的染色体定位、共线性和选择压力分析
从甘蓝型油菜数据库获取各BnPC基因相应的染色体位置等信息,利用TBtools(https://github.com/CJ-Chen/TBtools/releases)软件[31]中的基因可视化工具绘制BnPC基因在甘蓝型油菜19条染色体上的定位图。采用MCSCANX(http://chibba.pgml.uga.edu/mcscan2/)分析甘蓝型油菜(B. napus)、拟南芥(A.thaliana)、白菜(B. rapa)和甘蓝(B. oleracea)之间的基因复制事件和共线性关系[32]。为了确认BnPC之间的同源关系,以及BnPC与其它物种PC之间的同源关系,分别用TBtools 软件中的AmazingSuper-Circos 和Dual Systeny Plotter 进行可视化[31]。用KaKs 计算器2.0 计算各基因对的同义替换率(Ks)、非同义替换率(Ka)和Ka/Ks比值[33]。
1.3 BnPC家族蛋白系统发生分析
首先利用Clustal W 软件对BnPC 的PLCD 结构域序列进行多序列比对分析,生成软件MEGA 可识别的比对文件(默认参数设置)[34],然后利用MEGA5.0 软件(https://www. megasoftware. net/),采用邻接法(Neighbor-joining Method)基于上述多序列比对文件进行系统发生分析[35]。Bootstrap 设置为1000,选用p-distance 模型,Gap 设置为Partial deletion,Site Coverage Cutoff设置为95%,生成无根树。
1.4 BnPC家族的结构特征分析
将存在PLCD 结构域的BnPC 蛋白序列文件保存为FASTA 格式,利用SignalP 5.0(http://www.cbs.dtu.dk/services/signalP/)、Big-PI Plant Predictor(http://mendel. imp. ac. at/gpi/plant_server. html)和NetNGlyc 1.0 Server 软件(http://www. cbs. dtu. dk/services/NetNGlyc/)分别预测其N-端信号肽(NSP)、GAS 和N-糖基化位点(默认参数设置)。依据前人研究拟南芥AtPC 蛋白序列中潜在阿拉伯半乳聚糖蛋白样域(Arabinogalactan protein-like region,ALR)的原则[10]:以非邻接的脯氨酸(Pro)残基基序([Ala/Ser/Thr/Gly]-Pro-X(0, 10)-[Ala/Ser/Thr/Gly]-Pro)和([Ala/Ser/Thr/Gly]-Pro3-4)为阿拉伯半乳聚糖(AG)糖模体;Ser-Pro2-4为假定的伸展蛋白糖模体;手动分析BnPC 蛋白序列中潜在的ALR。利用软件Clustal X 1.81(http://www. clustal. org/clustal2/)对BnPC 氨基酸序列进行多重序列比对分析(使用默认参数)[34],手动标记结构域特征位点。
1.5 基因外显子/内含子结构及保守模体分析
从甘蓝型油菜数据库获得油菜BnPC基因的基因组序列及CDS 序列,利用基因结构预测的可视化工具TBtools 绘制BnPC的基因结构图[31]。利用在线软件MEME Suite5.1.0(http://meme. nbcr. net/meme/cgi-bin/meme. cgi)分析BnPC 蛋白的氨基酸序列,预测其中的保守模体(motif),参数设定为软件默认值[36]。
1.6 启动子顺式元件分析
从甘蓝型油菜数据库获得BnPC基因的起始密码子上游区域2000 bp 序列,利用在线网站Plant-Care(http://bioinformatics. psb. ugent. be/webtools/plantcare/html/)对上述区域进行顺式元件分析[37],利用TBtools软件对该区域的顺式元件进行可视化[31]。
1.7 基因表达分析
根据本实验室前期创建的甘蓝型油菜不同发育阶段各组织器官中以及不同逆境胁迫条件下的转录组数据(BioProject ID PRJNA687395),分析各BnPC基因的时空表达模式以及在不同逆境处理条件下的表达变化。对于时空表达模式,分别对各BnPC基因在甘蓝型油菜子叶、根、茎、幼叶、茎顶端分生组织(shoot apical meristem,SAM)、成熟叶、花苞、子房以及受精后14 d、24 d、34 d、50 d 的种子和角果皮中的表达水平进行分析。对于逆境表达模式分析,分别对各BnPC基因在干旱、盐、冷和高温胁迫条件下的表达变化情况进行分析。将甘蓝型油菜扬油9 号种子在湿润滤纸上发芽后,选取生长健康且长势一致的幼苗移栽至光照培养箱(22℃,16 h 光照/8 h 黑暗)进行水培,待幼苗生长至四叶期开始胁迫处理。本研究采用15% PEG6000 模拟干旱胁迫(drought),利用150 mmol/L NaCl 处理进行盐胁迫(salt),分别在4℃和42℃培养箱中进行冷胁迫(cold)和高温胁迫(heat),以种植在正常条件下培养的植株作为对照。分别在干旱和盐处理后24 h,冷处理后6 h,高温处理后1 h采集处理材料的叶片,并在对应的时间点同时采集对照植株的叶片,立即用液氮冷冻,在-80℃冰箱中保存至提取RNA。使用华中农业大学(武汉)Illumina 测序平台(HiSeq 3000)进行RNA-Seq测序。
2 结果与分析
2.1 BnPC家族的鉴定与分类
结果表明,甘蓝型油菜中有183 个蛋白具有PLCD结构域,其中10个蛋白由于保守PLCD结构域中关键Cys 残基缺失而被排除,因此,在甘蓝型油菜中共鉴定得到173 个PC 蛋白(附表1,见首页OSID二维码),远多于前人在拟南芥中的发现PC 成员的数量(38个)[28]。
对BnPC 家族成员的PLCD 结构域进行多重序列比对分析,结果表明:140 个成员的PLCD 结构域中含有完整的2 个Cys 残基,其余8 个BnPC 蛋白只含有一个Cys残基,Cys在PLCD结构域中高度保守;49个BnPC 蛋白中含有4个完整的铜结合配体(His、Cys、His、Met/Gln),其中28 个成员的铜配体为His、Cys、His、Met 类型,21 个成员的铜配体为His、Cys、His、Gln 类型(附表1,见首页OSID 二维码)。根据铜结合位点以及糖基化位点的差异,可以将BnPC蛋白划分为4 个亚家族:其中25 个成员归属于类花青苷蛋白(BnUCL),21 个成员归属于类漆树蓝蛋白(BnSCL),3 个成员归属于类质体蓝素蛋白(Bn-PLCL);其余99 个不含铜结合位点的BnPC 归属于类早期结瘤素蛋白(BnENODL)亚家族。与以往的研究结果不同,甘蓝型油菜中有5 个SCL(SCL5、SCL6、SCL13、SCL15 和SCL16)不具有N-糖基化位点,这可能是由于进化过程中蛋白序列的趋异造成的。此外,多重序列比对的结果显示,在BnENODL亚家族成员的序列中,与其它亚家族成员铜结合位点相对应的位置上有一些氨基酸取代His、Cys、His、Met/Gln从而形成了其它形式的保守模体,其功能可能与结合铜有关。
2.2 BnPC基因的染色体定位和共线性分析
对BnPC基因在甘蓝型油菜19条染色体上的分布进行分析。如图1 所示,在173 个BnPC基因中,有25个BnPC基因由于甘蓝型油菜基因组组装信息不全而未被定位到特定染色体上,其余148 个BnPC基因在19条染色体上分布不均。共有67个和81个BnPC基因分别位于A 和C 亚基因组上,每条染色体上的BnPC基因数目从4到15不等,C07染色体上包含的BnPC基因最多(15 个),其次是C03、A03 和C04 染色体,分别有13、12 和10 个BnPC。A08 和A07 染色体上的BnPC基因数目最少(分别有4 个),各条染色体上BnPC基因的数目与染色体长度不存在明显相关关系。
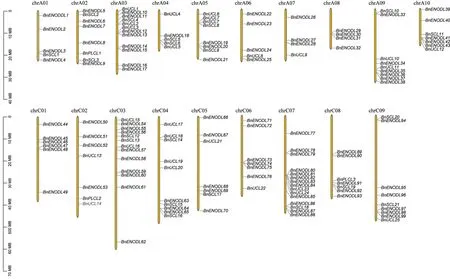
图1 BnPC基因在甘蓝型油菜染色体上的分布Fig.1 Distribution of BnPC genes on Brassica napus chromosomes
基因复制是物种进化过程中产生新基因的一种重要途径,主要是在多个基因非常接近的情况下通过串联复制,或者通过片段复制来实现[38]。为了研究BnPC 家族基因在进化过程中的扩增情况,本研究对BnPC 家族中的基因复制事件进行了分析。结果表明,137 个BnPC基因是复制事件的产物(附表2,见首页OSID二维码),其中123个BnPC来自全基因组复制(whole genome duplication,WGD)或片段复制(segmental duplication),另外14 个BnPC基因来自分散复制(dispersed duplication)。利用MCScanX 软件在BnPC 家族中共鉴定出120 对基因具有共线性。为了解甘蓝型油菜和拟南芥以及芸薹属植物BnPC基因家族间的进化关系,本研究进一步对甘蓝型油菜与自身、拟南芥以及两种二倍体祖先(白菜和甘蓝)基因之间的共线性关系进行了分析。如图2 和图3 所示,甘蓝型油菜与拟南芥、白菜、甘蓝中存在大量的同源PC基因。在148 个BnPC中,有118 个(79.73%)与其它物种的PC具有共线性关系,其中92 个BnPC基因(87 个来源于WGD 或片段复制,占94.57%)与拟南芥PC基因具有共线性关系,118 个BnPC(112 个来源于WGD 或片段复制,占94.92%)与白菜PC基因具有共线性关系,96 个Bn-PC(86个来源于WGD 或片段复制,占89.58%)与甘蓝PC基因具有共线性关系。上述结果表明,基因复制在很大程度上促进了甘蓝型油菜基因组中PC基因家族的扩张,其中WGD 或片段复制事件起主要的驱动作用。
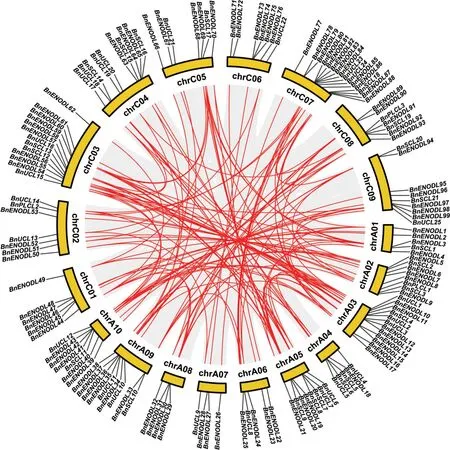
图2 甘蓝型油菜基因组内PC基因的共线性分析Fig.2 Syntenic relationship of PC genes in B.napus genome
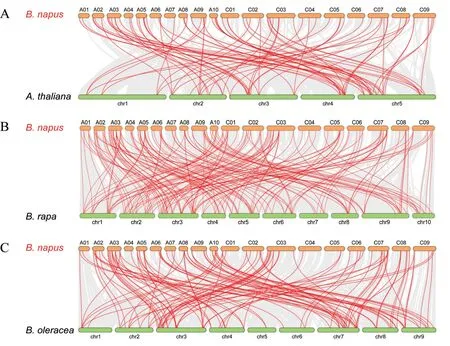
图3 甘蓝型油菜与拟南芥、白菜和甘蓝PC基因的共线性关系Fig.3 Syntenic relationship of PC genes in B.napus and three Cruciferae plants(Arabidopsis,B.rapa and B.oleracea)
为了评估在甘蓝型油菜进化过程中复制BnPC基因受到的选择压力,我们计算了甘蓝型油菜中的同源基因对的Ka、Ks和Ka/Ks比值。结果表明,大部分BnPC基因对的Ka/Ks<0.5,表明BnPC基因家族在进化过程中经历了强烈的纯化选择(附表3,见首页OSID二维码)。
2.3 BnPC家族成员的系统发生分析
为了阐明BnPC 蛋白之间的系统发生关系,我们利用PLCD 保守结构域序列对BnPC 蛋白进行无根树的构建。如图4 所示,BnPC 蛋白可分为6 个进化枝(Clade A-F)。与其它物种中的情况类似,Bn-PLCL 亚家族的成员最少,在甘蓝型油菜中仅有3 个成员,3个PLCL成员在进化关系上均与BnUCL亚家族比较接近,推测它们可能是由UCL 亚家族成员直接分化而来。有趣的是,与BnPLCL1、BnPLCL2 进化关系上相近的BnUCL23 却与UCL 亚家族其它成员相距较远,推测BnUCL23 可能是由BnENODL 亚家族分化而来,随后在进化过程中进一步分化为BnPLCL1 和BnPLCL2。成员数量最多的BnENODL亚家族,主要分布于三个进化枝,其中50 个BnENODL 占据了全部A 进化枝,C 进化枝中包含了32 个BnENODL,而剩余的BnENODL 几乎全部分布于F进化枝中,处于同一进化枝中的BnENODL 成员序列差异较小,可能具有类似的功能。除了BnUCL23 位于F 进化枝中,BnUCL 亚家族其它成员全部分布于B 进化枝中。BnSCL 亚家族成员中有3 个成员位于C 进化枝中,其余成员则主要存在于D 和E进化枝中,BnSCL成员分布相对分散,暗示着该亚家族成员在功能上存在分化。
对BnPC 蛋白的其它三个功能域(SP、ALR、GAS)进行分析(附表1,见首页OSID 二维码),根据四种结构域在BnPC 蛋白中的组成,BnPC 蛋白可分为10 种类型(I~X)(图5)。除VII、VIII、IX 和X 型成员外,共有133个BnPC 蛋白具有分泌蛋白所需要的N-端信号肽。V 型、VI 型和X 型成员具有两个PLCD 结构域,PLCD 结构域的重复可能来源于甘蓝型油菜进化过程中PLCD 结构域区段的复制。此外,属于II、IV、VIX 型的77 个BnPC 具有GAS,暗示着这些蛋白质可能定位于细胞膜上。除了I、II、VI和VII 型的BnPC 成员,其余95 个BnPC 具有一个ALR 域,在这些成员中,同时具有N-端信号肽的蛋白可归属于AGP 超家族。蛋白质结构域组成类似的PC 成员倾向于在系统发生树上聚集在一起。例如,I 型BnPC 主要聚集在E 进化枝中,IV 型BnPC 主要聚集在A进化枝中(图4)。
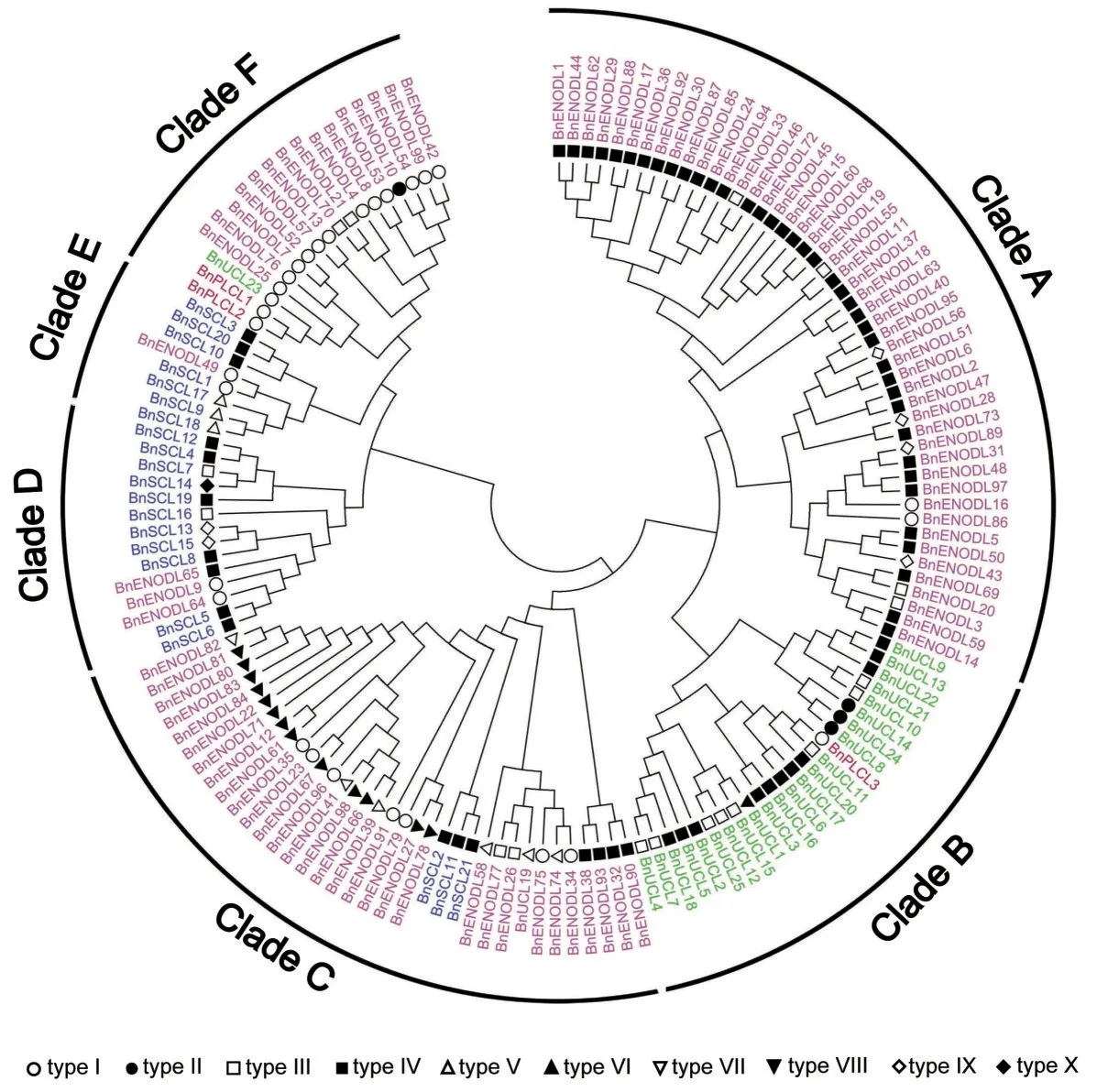
图4 甘蓝型油菜PC家族系统发生分析Fig.4 Phylogenetic analysis of the PC family in B.napus
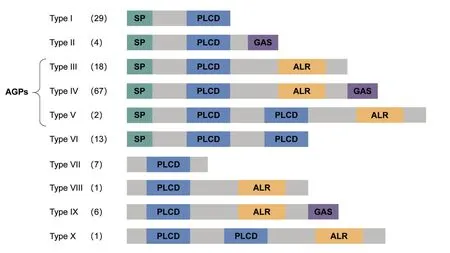
图5 基于结构域组成的BnPC分类Fig.5 Classification of BnPC proteins based on domain composition
2.4 BnPC家族成员的结构分析和模体分析
外显子-内含子结构的差异对于基因功能起重要作用,特别是对于重复基因[39]。本研究对148 个BnPC家族基因成员编码区序列进行结构分析的结果表明,BnPC基因的外显子数目在1 个到6 个之间,其中100个BnPC基因包含2个外显子,20个Bn-PC基因包含3 个外显子,9 个BnPC基因包含3 个外显子,18 个BnPC基因(BnENODL9/12/22/27/39/61/65/71/83/98、BnSCL4/5/6/7/8/12/13/15)只有1 个外显子,没有内含子,仅BnENODL42基因含有多达6 个外显子(图6)。此外,同一亚家族的同一系统发育类群的基因的外显子-内含子结构更相似,例如Clade A 中大多数成员有两个外显子,Clade D 中大多数基因没有内含子。属于相同亚家族的一些成员外显子长度接近,而内含子长度差异较大(例如:BnENODL30 和BnENODL87、BnUCL13 和BnUCL22、BnPLCL1和BnPLCL2),这可能与重复基因的功能分化有关。
利用MEME 分析BnPC 蛋白的模体组成,共鉴定出10 个保守模体,其长度在14~50 个氨基酸之间(图6,附图1,见首页OSID二维码),其中Motif 1/2/3/4/5/7/8 属于PLCD 保守结构域的一部分,Motif 6 出现在具有两个PLCD 保守结构域的蛋白中,位于两个PLCD 结构域之间,Motif 9/10 则是新发现的保守结构域。结果表明,同一亚类中BnPC 的结构相似。除了BnENODL58、BnENODL91 和BnSCL16 外,其他所有BnPC 都包含Motif 1,Motif 2/3/4/5 也存在于大部分BnPC 成员中,Motif 6/7/8/9 在Clade C 的成员中较为保守,部分PC成员的N端和C端包含Motif 10。
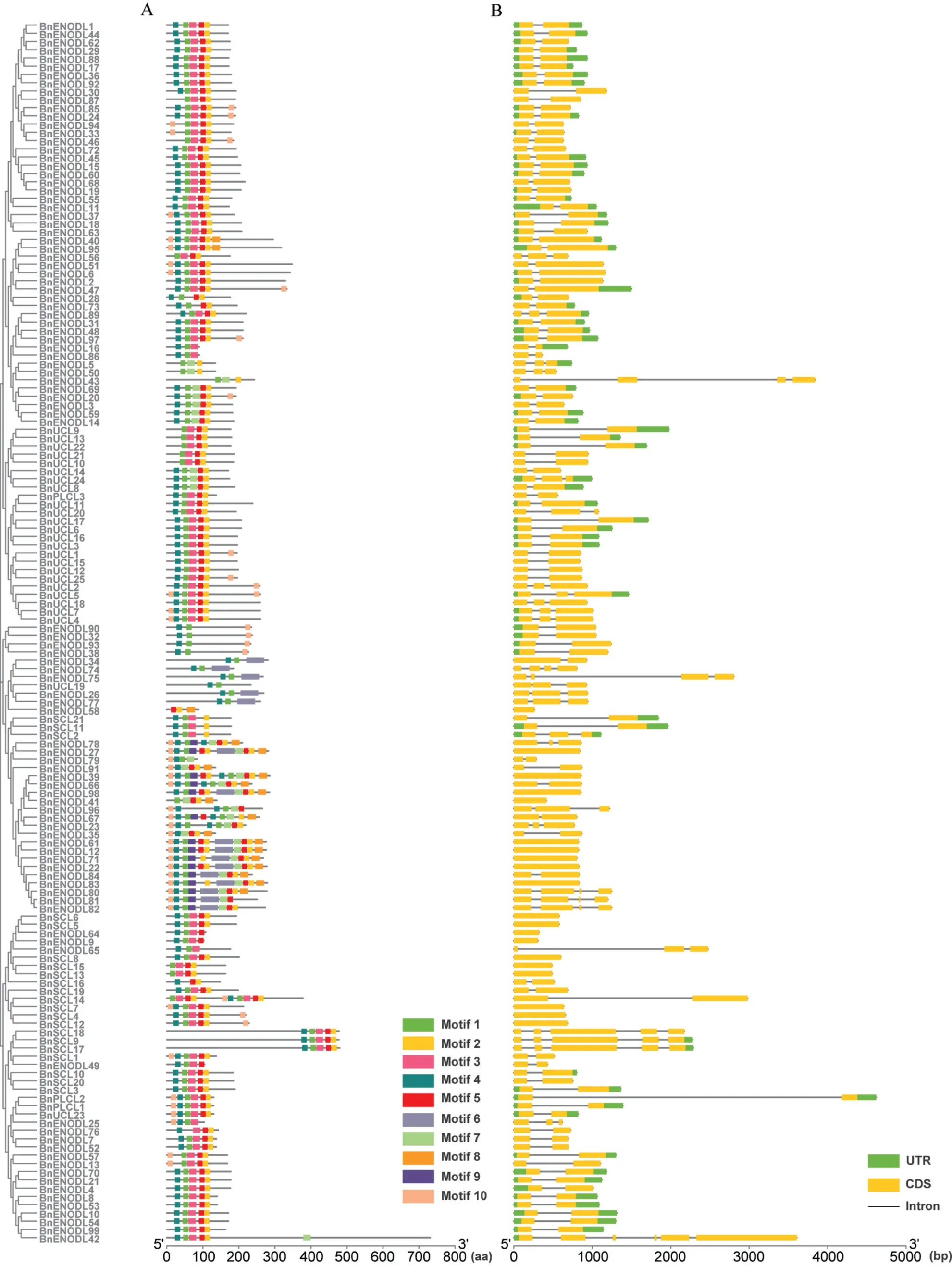
图6 BnPC家族成员的模体组成和基因结构分析Fig.6 Motif analysis and gene structure of the BnPC family members
2.5 BnPC 基因启动子区具有丰富的顺式调控元件
顺式作用元件在基因表达调控中起重要作用,基因启动子区域包含的不同类型的顺式调控元件可能介导基因在不同条件下表达。本研究利用PlantCARE 软件对148 个BnPC基因的启动子区域的顺式元件进行了分析,结果发现了10种顺式作用元件,主要可分为以下4 类:与植物生长发育、非生物胁迫应答、激素应答相关的元件,以及真核生物中的基本启动子元件(如CAAT 盒和TATA 盒)。与生长发育相关的启动子元件主要包括参与胚乳表达的GCN4 元件和AACA 元件;与分生组织表达相关的CAT元件;以及大量的光响应元件,如Box4、GBox 和TCT 元件(附表4,见首页OSID 二维码)。在BnPC启动子中,与光反应有关的元件是最常见的元件,此外,本研究还鉴定到一些与赤霉素(gibberellin acid,GA)、生长素(auxin)、脱落酸(abscisic acid,ABA)、MeJA 和水杨酸(salicylic acid,SA)应答相关的顺式元件,其中与ABA 应答相关的元件最为丰富,其次是与MeJA 反应相关的元件(表1,附表4,见首页OSID 二维码)。此外,在BnPC的启动子区域还发现了许多与胁迫答应相关的元件,如低温应答相关的低温应答元件(low-temperature responsiveness,LTR)、干旱诱导的MYB 结合位点(MBS)、与创伤反应相关的创伤应答元件(wound-responsive element)和厌氧诱导所必需的厌氧调节元件(anaerobic induction)。这些结果表明BnPC可能在调节植物生长发育、对非生物胁迫的应答和激素应答中发挥作用。大多数BnPC基因的启动子区具有不同类型顺式元件,暗示这些BnPC基因可能参与多种生理过程和调控途径。
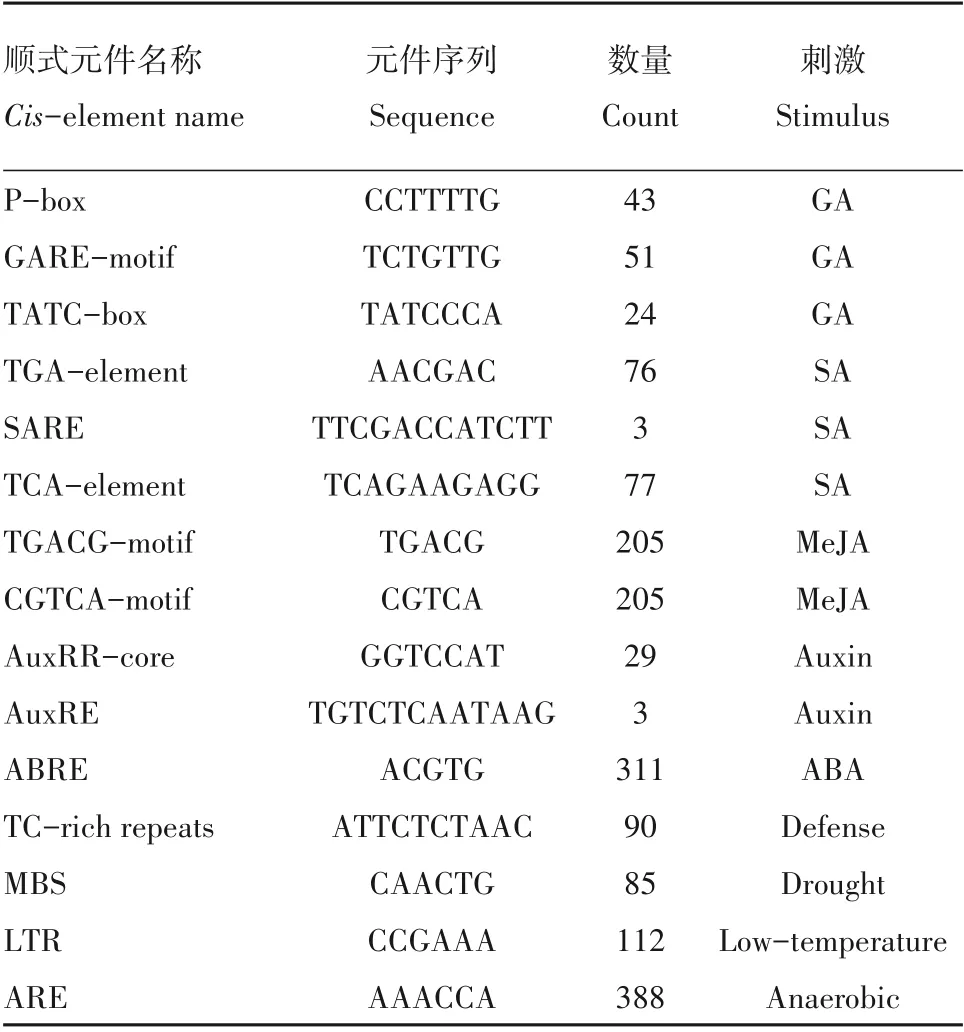
表1 BnPC基因启动子顺式元件分析Table 1 Cis-acting elements of the promoter regions of BnPC genes
2.6 BnPC基因的时空表达与逆境应答模式
基于甘蓝型油菜不同发育阶段各组织器官的转录组测序数据对BnPC基因的时空表达模式进行分析(图7A,各基因FPKM 值详见附表5,首页OSID二维码),结果显示,42 个BnPC基因在所有检测的组织器官中均未表达或者表达水平极低(FPKM<0.5,图7 中未显示)。根据时空表达特点,BnPC基因大致可以分为四种类型。第一种类型在所有组织器官中表达水平均较低或未表达。第二种类型在几乎所有组织器官中均有表达,且在各组织器官中的表达水平差别不明显,如BnENODL32、BnENODL38、BnENODL42和BnENODL93。第三种类型在大多数组织中表达,但在特定组织中高表达,如BnUCL9、BnUCL13和BnUCL22。第四种类型基因的表达表现出很强的组织器官特异性:例如BnUCL6、BnUCL17和BnUCL20只在根(Root)中表达,而BnENOD31、BnENODL48、BnENODL55、BnENODL97和BnUCL23只在受精后24 d 的种子(Seed-24 DAP)和角果皮(Silique wall-24 DAP)中表达。此外,几乎所有的BnUCL基因均在根中具有高水平的表达(除了BnUCL23),暗示着BnUCL亚家族成员可能在根的发育中起着重要作用,而BnPLCL3与BnUCL亚家族成员表现出非常相似的表达模式,这与上述系统发生分析的结果相符,BnPLCL3是由BnUCL亚家族成员进化而来,BnPLCL3与BnUCL亚家族成员在功能上可能也极为相似;而BnPC家族中其余的2 个BnPLCL基因BnPCLC1和BnPCLC2在各个组织中均有一定程度的表达。BnENODL亚家族的成员在甘蓝型油菜不同组织器官中的表达呈现一定程度的分化,9 个成员(例如BnENODL21/70/37/18/63/7/52/6/51)在受精后14 d 的种子(Seed-14 DAP)中特异表达,9 个成员(例如BnENODL48/97/31/89/11/55/16/30/86)在受精后24 d 的种子(Seed-14 DAP)中表达水平较高,8 个成员(例如BnENODL28/73/59/3/5/14/20/69)在花苞(bud)中特异表达,说明BnENODL亚家族中的这些成员可能参与了甘蓝型油菜花和幼嫩种子的发育过程。
为探究BnPC基因对各种非生物逆境胁迫的应答模式,本研究利用转录组数据分析了在干旱、盐、冷和高温处理条件下各BnPC基因的表达水平(图7B,附表6,见首页OSID 二维码)。结果表明,95 个BnPC基因的表达水平在至少一种逆境处理前后发生变化,53 个基因在胁迫处理前后的表达水平没有明显变化。与组织器官表达数据对比发现,前文所述的在各组织器官中未表达和弱表达的42 个BnPC基因中,除了4 个基因(BnENODL9、BnENODL62、BnENODL71和BnENODL91)在逆境处理后表达明显上调,其余38 个基因仍检测不到表达,推测这38个基因可能在甘蓝型油菜的进化过程中因为功能丧失而不表达,或由于表达部位非常特异而在本文使用的数据中没有检测到表达量。在95 个逆境处理条件下表达水平发生变化的基因中,在干旱处理条件下,有38个基因上调表达,48个基因下调表达;在盐处理条件下,24 个基因表达量上升,59 个基因表达量下降;在冷处理条件下,25 个基因上调表达,58 个基因下调表达;在高温处理条件下,40 个基因表达量上升,44 个基因表达量下降。这些结果表明,相当一部分BnPC基因对非生物胁迫表现出明显的响应,暗示着BnPC可能也广泛参与甘蓝型油菜对不利环境条件的适应。
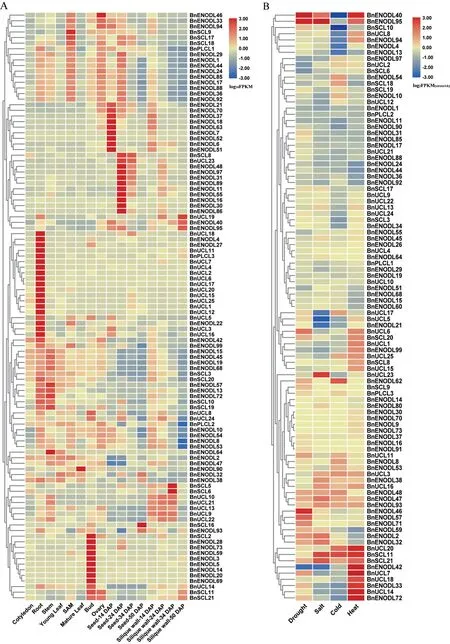
图7 BnPC基因的表达特征Fig.7 Expression profile of BnPC genes
3 讨论
PC基因作为古老的蓝铜蛋白基因,可能与铜原子结合,在各种生物系统中起电子转运的作用,如铜的转运和植物光合作用,因此PC基因家族成员可能受到光诱导广泛参与植物的生长发育[28]。在BnPC基因启动子中,我们也发现了大量的光反应元件,表明PC基因可能对甘蓝型油菜的生长发育有关键性作用。目前,植物中对PC基因家族的相关研究比较少,在油料作物甘蓝型油菜中PC基因家族的综合分析尚未见报道。本研究在甘蓝型油菜中鉴定到173 个BnPC基因。根据铜配体残基的特性、蛋白结构组分的差异,将这些BnPC基因分为BnENOLD、BnUCL、BnSCL和BnPLCL4 个亚家族,其中BnENODL亚家族基因数量最多,BnUCL亚家族、BnSCL亚家族其次,BnPLCL亚家族基因数量最少,与之前在拟南芥[10]和水稻[9]中报道的结果类似。Bn-PC基因结构的变化可能是由于在长期进化过程中内含子/外显子的增加或减少所致。基于BnPC 保守模体分析的结果,推测BnPC基因家族的功能分化可能是由类特异性和亚类特异性模体引起的。此外,在BnPC 的每个亚类中,相似的模体结构、外显子-内含子结构组成模式和氨基酸组成可能预示着进化过程中的基因复制事件。
甘蓝型油菜是一种由白菜和甘蓝自然杂交形成的异源四倍体植物[30]。基因复制有两种主要机制,WGD/片段复制和串联复制[40]。WGD/片段复制在植物基因组中很常见,许多植物在进化过程中经历了多倍化事件,因此在它们的基因组中保留了许多重复的染色体片段[41,42]。前人在拟南芥[10]、水稻(O.sativa)[9]、大豆(Glycine max)[4]、大白菜[28]、玉米[23]、蝴蝶兰[29]、蒺藜苜蓿[4]和铁皮石斛[43]中分别鉴定到38、62、90、84、60、30、82 和38 个PC基因,甘蓝型油菜中PC家族基因的数量(173 个)远远多于上述植物物种中PC基因的数量,说明在甘蓝型油菜进化过程中,基因组复制对BnPC基因的多样性有很大贡献。我们的结果表明,137 个BnPC基因是基因复制的结果,其中123 个来源于WGD 或片段复制,其余是分散复制的结果。串联复制是另一种类型的基因复制,其特征是一个基因家族的多个成员存在于同一个或相邻的基因间区域[40]。本研究发现,4 个BnPC基因:BnSCL8、BnENODL81、BnENODL82和BnENODL83来源于这种复制方式。为了比较不同物种中PC基因家族扩张机制的异同,我们也分析了其他物种中PC基因家族成员的复制类型。水稻中20 个OsPC来源于片段复制,18个来源于串联复制[9];玉米中14 个ZmPC来源于片段复制,8 个来源于串联复制[23];而在蒺藜苜蓿中,40 个MtPC来源于串联复制,其余32 个来源于其他复制方式[4]。在这些物种中,片段复制和串联复制在PC基因家族扩张中均起着重要作用。有趣的是,拟南芥中有12 个AtPC来源于片段复制,仅有2个PC来源于串联复制[10];白菜中63 个PC来源于片段复制,没有PC基因来源于串联复制[28]。与白菜和拟南芥中的情况类似,BnPC基因家族中仅有4个基因来源于串联复制,这表明WGD/片段复制是BnPC基因家族扩张的主要动力,该现象是否在芸薹属甚至十字花科植物中普遍存在,还有待进一步研究。在人类基因组中,诸如转座子插入和拷贝数变异等分散复制是非常常见的[44],但在植物中鲜有报道,已报道的植物物种中尚未发现有PC基因来源于这种复制方式。本研究发现,有14 个BnPC基因来源于分散复制。在异源多倍体植物中,基因组的扩张通常伴随着转座子因子的插入[45,46]。芸薹属植物的转座子也很丰富[46,47],然而,这些转座子是否是PC基因家族扩张的原因之一还需要进一步的研究。
PC 蛋白成员主要包含下列4 个结构域,结构域I 包含内质网靶向信号肽,介导PC 进入分泌途径。结构域II 是一个保守的PLCD 结构域,负责使PC 与铜氧化物结合,该结构域包含天冬酰胺-X-苏氨酸/丝氨酸(Asn-X-Thr/Ser)序列,提供潜在的N-糖基化位点,并进一步通过内质网分泌途径促进蛋白分拣,所有的PC 都至少有一个PLCD 域;结构域Ⅲ是一个类阿拉伯半乳聚糖蛋白域(arabinogalactan protein-like region,ALR)[4]。含有ALR 结构域的PC 被认为是形成细胞壁的氧化还原活性成分,并参与防御反应[5]。结构域IV 具有GAS的特征,能够将PC锚定在细胞膜上[5]。根据结构域组成,我们将BnPC 蛋白分成10 类(图5),而在蒺藜苜蓿[4]和大白菜[28]中,分为9类。在甘蓝型油菜中,我们比蒺藜苜蓿[4]多鉴定到Type VI 和Type X 成员,但并未鉴定到SPPLCD-PLCD-ALR-GAS 结构域形式组成的成员;与大白菜相比,我们多鉴定到Type VIII、Type IX 和Type X,但并未鉴定到PLCD-PLCD 和PLCD-GAS结构域组成形式的成员。而甘蓝型油菜由白菜和甘蓝自然杂交形成[30],我们推测在甘蓝型油菜起源过程中,PLCD-PLCD 和PLCD-GAS 结构域组成的成员可能发生了丢失,而Type VIII、Type IX 和Type X 的成员可能在进化过程中由Type III、Type IV 和Type V成员丢失结构域I进而演化而来。
BnPLCL是成员最少的亚家族,在甘蓝型油菜中仅有3 个成员,其中BnPLCL3仅在根中微弱表达而BnPLCL1和BnPLCL2在所有组织中均表达,推测这两个基因对植株的正常发育可能极为重要。几乎所有的BnUCL基因均在根中具有较高水平的表达量(除了BnUCL23),特别是BnUCL3和BnUCL16在其它组织中微弱表达或者不表达,但在根中的表达量非常高,暗示BnUCL亚家族成员可能在根的发育中起着重要作用;除此之外,BnUCL3/10/13/21/22在14 d、24 d和34 d角果皮中均高表达,而在其它组织中几乎不表达,说明这5 个基因可能在角果发育中发挥重要作用。SCL基因表达水平普遍较低,主要在根和茎中表达,有趣的是,BnSCL2基因是BnPC基因中在花苞中表达水平最高的基因,远超其它基因,且这个基因的表达受非生物胁迫影响不明显,推测这个基因在花苞中的功能可能非常重要且稳定。前人的研究结果表明BnENODL基因在根中高表达,参与了豆科植物的结瘤[4,14,15],与前人研究结果相似,我们发现大部分BnENODL基因也均在根中高表达,表明BnENODL基因在根中的功能可能十分关键。某些BnENODL基因在部分组织器官中特异性高表达,比如BnENODL21/37和BnENODL31/48/55/97分别在受精后14 d 和24 d 的种子中高表达,这些基因可能在种子发育过程中发挥重要功能。
本研究结果对甘蓝型油菜BnPC基因家族在全基因组水平上进行了系统分析,鉴定出173 个BnPC成员,并对它们的染色体分布情况、序列特征、系统发生关系、启动子顺式元件组成以及时空表达模式和逆境响应情况进行了分析,为深入研究甘蓝型油菜BnPC 蛋白功能和挖掘该家族中有利于甘蓝型油菜遗传改良的基因资源提供了一定的理论依据。
