以六倍体(AnAnCnCnCoCo)为桥梁创制抗旱新型甘蓝型油菜(AnArCnCo)
2020-09-11万华方魏帅冯宇霞钱伟
万华方,魏帅,冯宇霞,钱伟
(西南大学农学与生物科技学院/西南大学农业科学研究院,重庆 400716)
0 引言
【研究意义】甘蓝型油菜(Brassica napus,2n=38,AACC)大约起源于 7 500年前,是由白菜型油菜(Brassica rapa,2n=20,AA)和甘蓝(Brassica oleracea,2n=18,CC)杂交,并自然加倍形成的异源四倍体物种[1-2],是温带地区第二重要的油料作物[3]。干旱严重影响作物(包括油菜)的生长发育和经济产量[4]。随着菜籽油需求量的不断上升,油菜产业受到干旱的威胁日益严重[5],仅中国长江流域的油菜受秋旱影响就可减产20%左右[6]。因此,改良甘蓝型油菜的抗旱性是一个亟待解决的科学问题。然而,有限的原始亲本生态型、暂无其野生群体,以及“双低”品质选育过程引起的种质资源遗失,致甘蓝型油菜遗传背景狭窄[2,7-9],进而限制了甘蓝型油菜的抗旱育种研究[10]。利用其祖先种向甘蓝型油菜导入外源遗传成分是拓展其遗传背景的有效途径[11-15]。白菜型油菜 A基因组的遗传渗入有利于拓展甘蓝型油菜的遗传背景[16]。白菜型油菜的农艺性状变异丰富[17-19],并具有耐迟播、生育期短、耐贫瘠、耐干旱等突出优点[20],是甘蓝型油菜遗传改良的重要资源[11,21-23]。因此,进行白菜型油菜抗旱性资源的鉴定,并以其为亲本合成具有优良抗旱性的新型甘蓝型油菜,对于提高油菜抗旱性能具有重要意义。【前人研究进展】利用亲本物种丰富的遗传变异拓宽遗传背景是改良现有甘蓝型油菜的一个重要方法[24]。RIPLEY等[25]利用甘蓝与甘蓝型油菜杂交,成功将甘蓝的自交不亲和性导入甘蓝型油菜。岳芳等[26]以六倍体(ArArAnAnCnCn)与甘蓝种间杂交,合成了新型甘蓝型油菜。钱伟等[27]以甘蓝型油菜和甘蓝为亲本,创建了六倍体(AnAnCnCnCoCo),并以其为桥梁,利用白菜型油菜丰富的遗传资源拓宽了现有甘蓝型油菜的遗传背景。MEI等[28]以六倍体(AnAnCnCnCoCo)作为桥梁,与甘蓝型油菜不断回交,获得了转移野生甘蓝菌核病抗性的新型甘蓝型油菜。李勤菲等[29]以六倍体(AnAnCnCnCoCo)为桥梁,将甘蓝型油菜亲本种的优良性状导入甘蓝型油菜中。【本研究切入点】甘蓝型油菜相对狭窄的遗传背景限制了其抗旱育种。耐干旱的白菜型油菜是重要的抗旱资源,但以其与甘蓝型油菜杂交、自交或回交选育新型甘蓝型油菜所需的周期长,效率低[29]。六倍体是拓宽甘蓝型油菜遗传背景的新材料[27-29],但以其为桥梁,转移白菜型油菜优良抗旱性的研究鲜有报道。【拟解决的关键问题】本研究通过PEG-6000溶液模拟干旱,评价白菜型油菜的抗旱性,并以六倍体材料为桥梁,以抗旱的白菜型油菜为抗性父本,创建具有优良抗旱性的新型甘蓝型油菜,为甘蓝型油菜抗旱育种提供优良的种质资源。
1 材料与方法
1.1 试验材料
以11份六倍体作为母本(电子附表1)、68份白菜型油菜(电子附表 2)作为父本,创建新型甘蓝型油菜。
9份白菜型油菜、9份新型甘蓝型油菜和甘蓝型油菜中双11号(抗旱性好,种植广泛)为抗旱性分析材料(电子附表3),以中双11号为对照。
1.2 PEG-6000模拟干旱
参照SHANGGUAN 等[30]方法,设 0(CK)、100、150、200、250和300 g·L-16个PEG-6000质量浓度,对不同的 9份白菜型油菜在萌发期进行模拟干旱处理;设 0(CK)、100、150、200、250、300、350、400和600 g·L-1PEG-6000质量浓度,对9份新型甘蓝型油菜和中双11号在萌发期进行模拟干旱处理。分别筛选用于白菜型油菜和甘蓝型油菜抗旱性鉴定的PEG-6000最适浓度。
取直径9 cm的培养皿,铺垫双层滤纸,并以相应浓度的PEG-6000溶液5 mL浸润滤纸;取种子30粒均匀放置于上述浸润过的滤纸上,并盖上培养皿盖,置于组培室(25℃黑暗8 h/27℃光照16 h)培养,每天定时向培养皿中加入对应浓度的1 mL PEG-6000溶液,每个处理2个重复。以清水浸泡滤纸为对照。
1.3 检测指标
参照陈郡雯等[31]方法进行指标检测。每天定时观察种子萌发的粒数,测量胚根部位长度,以其为种子直径的一半为萌发标准。按照萌发的基本情况,以第 3天的种子萌发数值计算萌发势和相对萌发势,以第8天的发芽数值计算萌发率和相对萌发率,并且在第8天随机选出10株,计算其相对根长和相对胚轴长。
种子萌发抗旱指数(drought resistance index,DRI)=水分胁迫下种子萌发指数(plumule index of stress,PIS)/对照种子萌发指数(plumule index of control,PIC);萌发指数(plumule index,PI)=1nd2+0.75nd4+0.5nd6+0.25nd8(nd2、nd4、nd6、nd8 相对应的是当天的种子萌发率),包括PIS和PIC类型;
萌发率(germination rate,GR)=正常萌发的种子数/供试种子的总数×100%;
相对萌发率(relaltive germination rate,RGR)=(处理萌发率/对照萌发率)×100%;
萌发势(sprout potential,SP)=达到高峰(第 3天萌发的种子数)时正常萌发的种子数/供试种子的总数;
相对萌发势(relative sprout potential,RSP)=(处理萌发势/对照萌发势)×100%;
相对根长(relative root length,RRL)=(处理根长/对照根长)×100%;
相对胚轴长(relative stem length,RSL)=(处理胚轴长/对照胚轴长)×100%。
1.4 数据分析与处理
采用SAS v8.0软件进行数据分析。参照陈郡雯等[31]方法进行综合隶属函数值法分析,公式为R(Xi)=(Xi—Xmin)/(Xmax—Xmin)。反函数隶属函数值计算公式为R(Xi)=1—(Xi—Xmin)/(Xmax—Xmin),式中Xi指试验数据,Xmax、Xmin表示的是所对应数据的最大值和最小值。
2 结果
2.1 新型甘蓝型油菜的鉴定
以11份六倍体与68份白菜型油菜杂交,获得124份新型甘蓝型油菜。在苗期(图1-Ⅰ、图1-Ⅱ和图1-Ⅲ),新型甘蓝型油菜的表型介于两亲本之间。体细胞观察结果表明,所利用的六倍体的体细胞染色体数目为56(图1-Ⅳ),而合成的新型甘蓝型油菜的体细胞染色体数目为38(图1-Ⅴ)。花粉育性结果表明,六倍体的花粉育性约90%(图1-Ⅵ),新型甘蓝型油菜的花粉育性约80%(图1-Ⅶ)。在苗期,随机选择59份长势优良的新型甘蓝型油菜、7份六倍体、10份白菜型油菜以及20份自然甘蓝型油菜提取DNA进行分子标记鉴定,并进行主成分分析。第一、第二和第三主成分的贡献率分别为13.92%、11.59%和9.09%。主成分分析将所有的供试材料分成3类(图1-Ⅷ),即白菜型油菜、六倍体、甘蓝型油菜(包括新型甘蓝型油菜和自然甘蓝型油菜),但新型甘蓝型油菜与自然甘蓝型油菜明显分开,表明以六倍体为桥梁创建新型甘蓝型油菜切实可行。
2.2 筛选合适的PEG-6000浓度
依据染色体数目、花粉育性,从59份新型甘蓝型油菜中选取9份进行模拟抗旱研究(图2),在不同浓度 PEG-6000溶液的处理下,油菜种子萌发均受到不同程度的抑制,并且随着浓度梯度增加,平均根长随之降低。当PEG-6000浓度达到250 g·L-1时,9份新型甘蓝型油菜与中双 11号的平均根长为对照的40.2%(图 2-Ⅰ),根长相对值变异系数最大,表明不同材料的平均根长有显著性差异,250 g·L-1的处理浓度为新型甘蓝型油菜模拟干旱的最适处理浓度(图2-Ⅱ)。当PEG-6000浓度达到200 g·L-1时,白菜型油菜的平均根长为对照的30.4%(图2-Ⅲ),白菜型油菜根长相对值变异系数最大,表明不同材料的平均根长有显著性差异,200 g·L-1的处理浓度为白菜型油菜模拟干旱的最适处理浓度(图2-Ⅳ)。
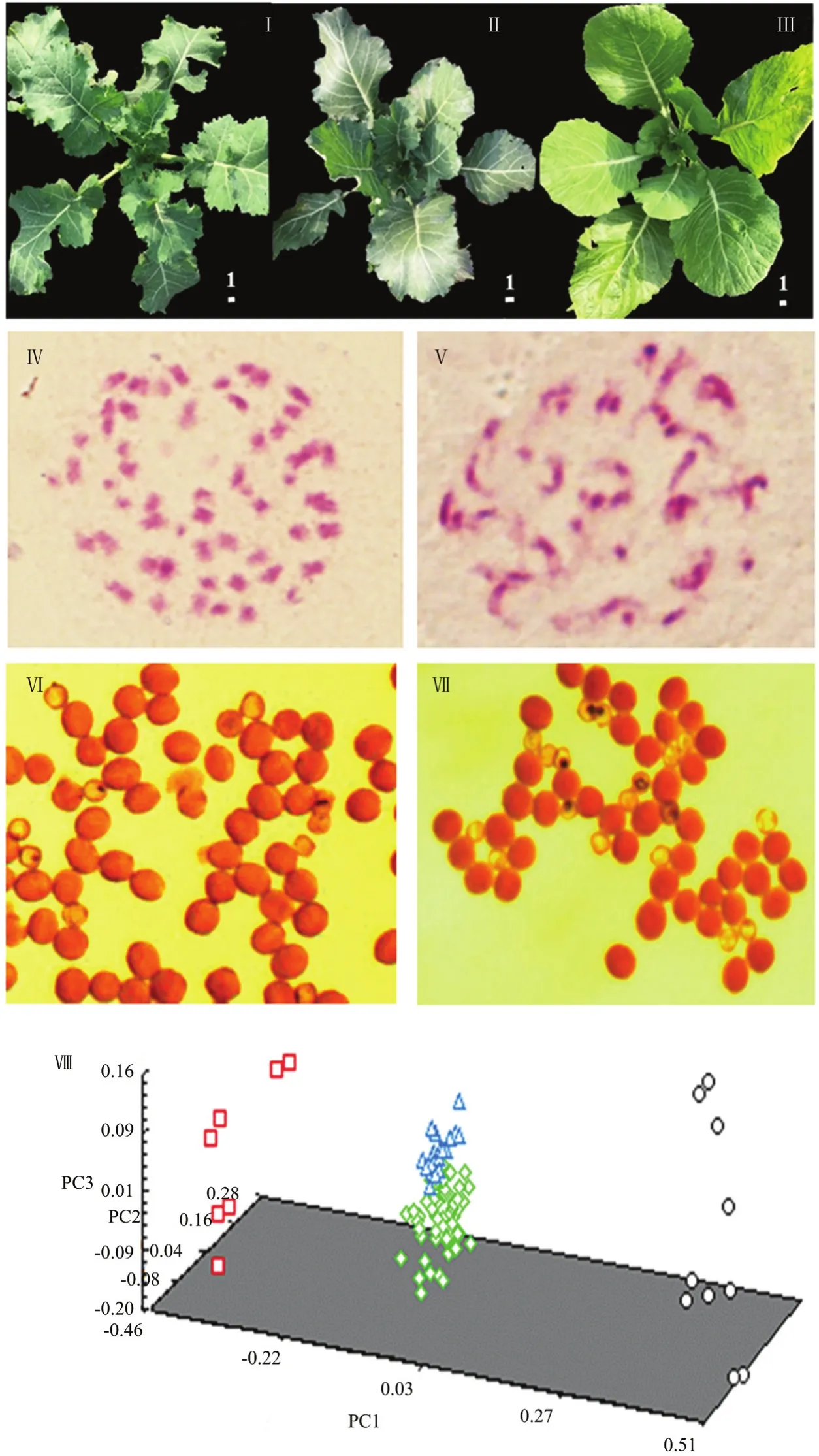
图1 新型甘蓝型油菜的鉴定Fig.1 Identification of new-type B.napus
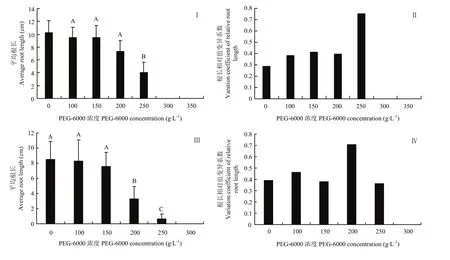
图2 油菜在不同浓度PEG-6000条件下根长的变化Fig.2 Root length of rapeseed treated by PEG-6000 solution with different concentration
2.3 油菜萌发期抗旱性相关指标测定及主成分分析
使用250 g·L-1的PEG-6000溶液对新型甘蓝型油菜和中双11号进行处理,用200 g·L-1的PEG-6000溶液对白菜型油菜进行处理,统计相对萌发势、相对萌发率、相对根长、相对胚轴长以及萌发抗旱指数,构成主成分分析的原始矩阵(表1)。
使用250 g·L-1的PEG-6000溶液对新型甘蓝型油菜进行模拟干旱处理,选用5个指标的相对值进行主成分分析(表 2),第一、第二、第三主成分的贡献率分别为48.62%、35.47%和11.93%。以200 g·L-1的PEG-6000溶液对白菜型油菜进行模拟干旱处理,对5个指标进行主成分分析。第一、第二、第三主成分的贡献率分别为71.61%、19.01%和6.43%。由此可见,第一、第二和第三主成分所包含的信息集中反映了甘蓝型油菜和白菜型油菜萌发期抗旱性的各项指标在干旱胁迫环境下的相互关系,由此建立起来的评价体系可用于甘蓝型油菜和白菜型油菜萌发期的抗旱能力评价。
2.4 油菜萌发期抗旱性隶属函数分析
运用综合隶属函数法对新型甘蓝型油菜与白菜型油菜萌发期模拟抗旱检测指标进行分析,并以5个抗旱指标的平均值为指标评价抗旱性(表 3)。结果表明,白菜型油菜8M664、8M655、8M693相比其他白菜型油菜,均具有更好的抗旱性。新型甘蓝型油菜9X018、9X015、9X016的抗旱性优于中双11号,9X111的抗旱性基本与中双11号持平,其他供试材料的抗旱性均低于中双11号。
通过对新型甘蓝型油菜 9X111、9X016、9X015和 9X018及其父本白菜型油菜 8M693、8M664、8M655和8M664的隶属函数值进行相关性分析(表4),发现9X111与其父本8M693呈现正相关(r=0.672),9X016与其父本8M664呈现显著正相关(r=0.864,P<0.05),9X015与其父本 8M655呈现正相关(r=0.372),9X018与其父本 8M664呈现显著正相关(r=0.991,P<0.01)。结果表明,以六倍体为桥梁,可将白菜型油菜的优良抗旱性导入新型甘蓝型油菜,进而获得抗旱性提高的甘蓝型油菜。
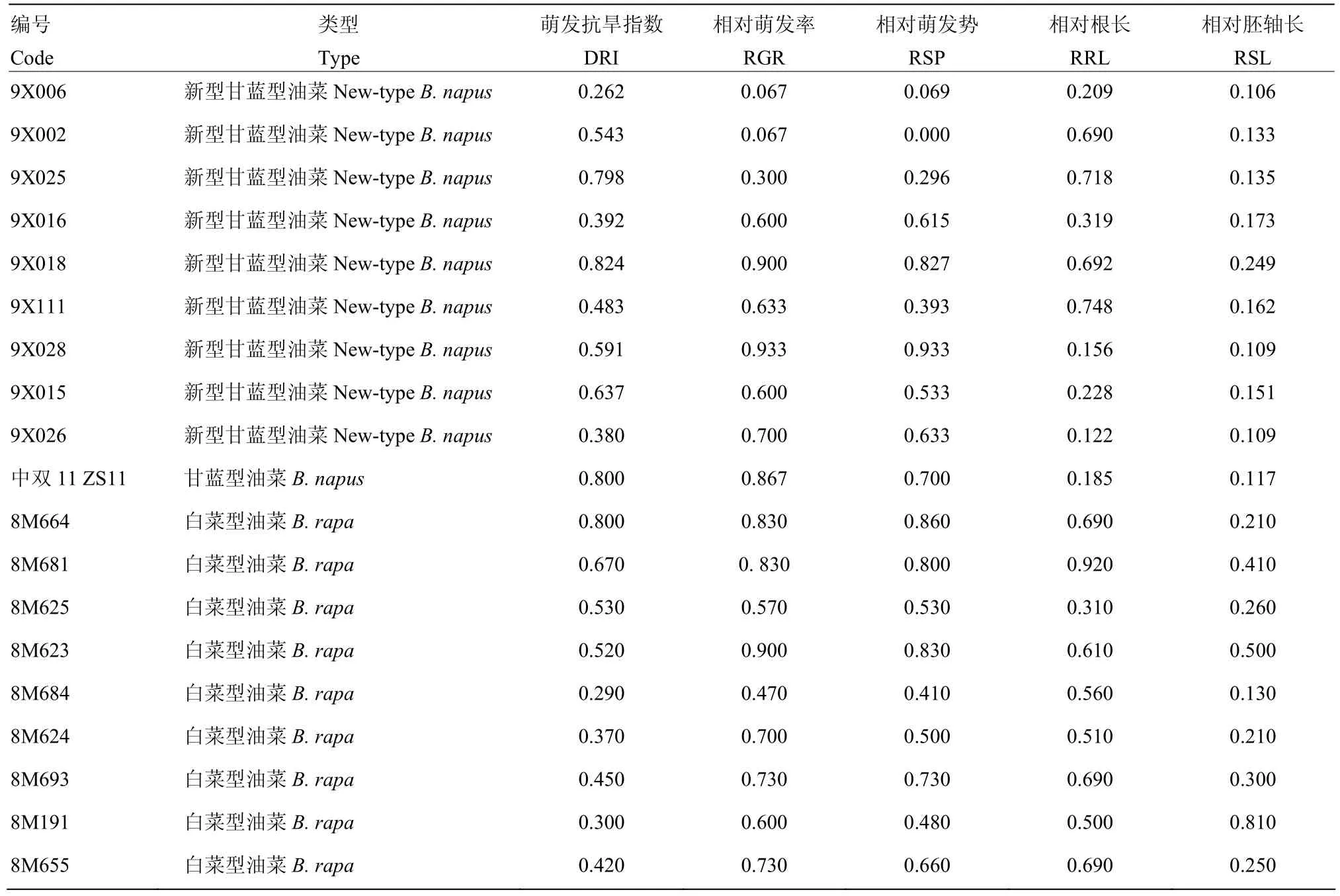
表1 油菜萌发期抗旱性相关指标主成分分析原始矩阵Table 1 Primitive matrix of principal component analysis on drought-resistance indices of rapeseed at germination stage
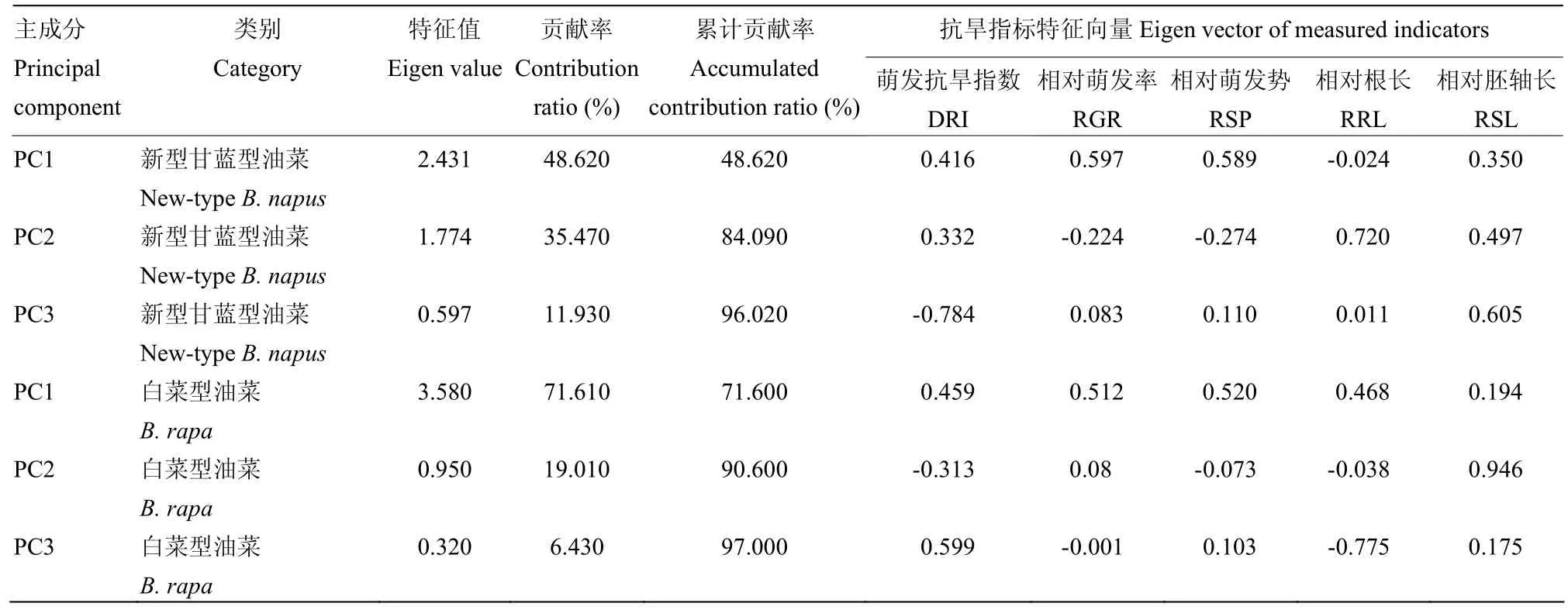
表2 模拟干旱相关指标的主成分分析Table 2 Principal component analysis on drought-resistance-related indicators on rapeseed
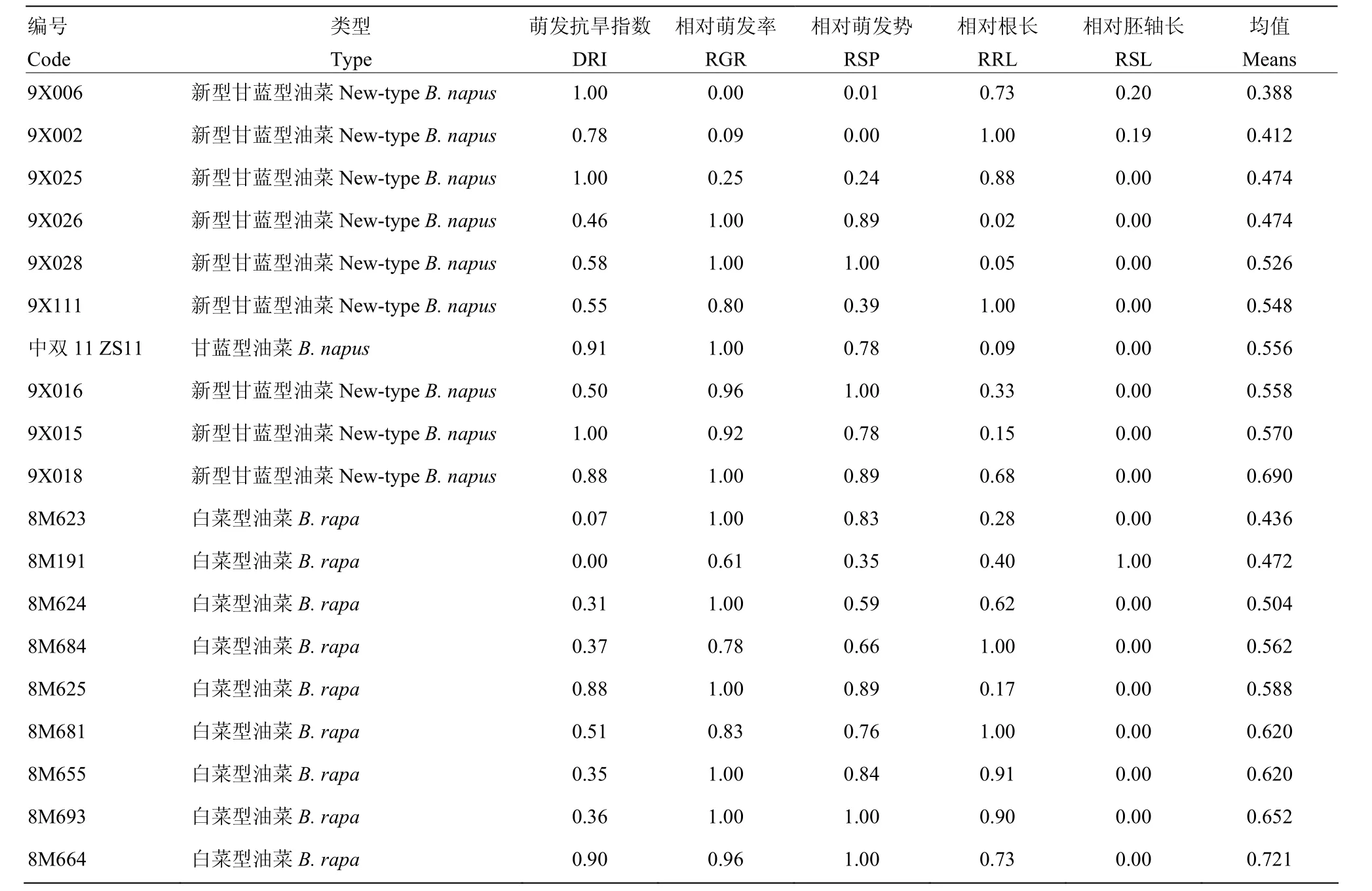
表3 模拟干旱相关指标的隶属函数值Table 3 Membership function value of drought-resistance-related indicators of rapeseed under simulated drought stress
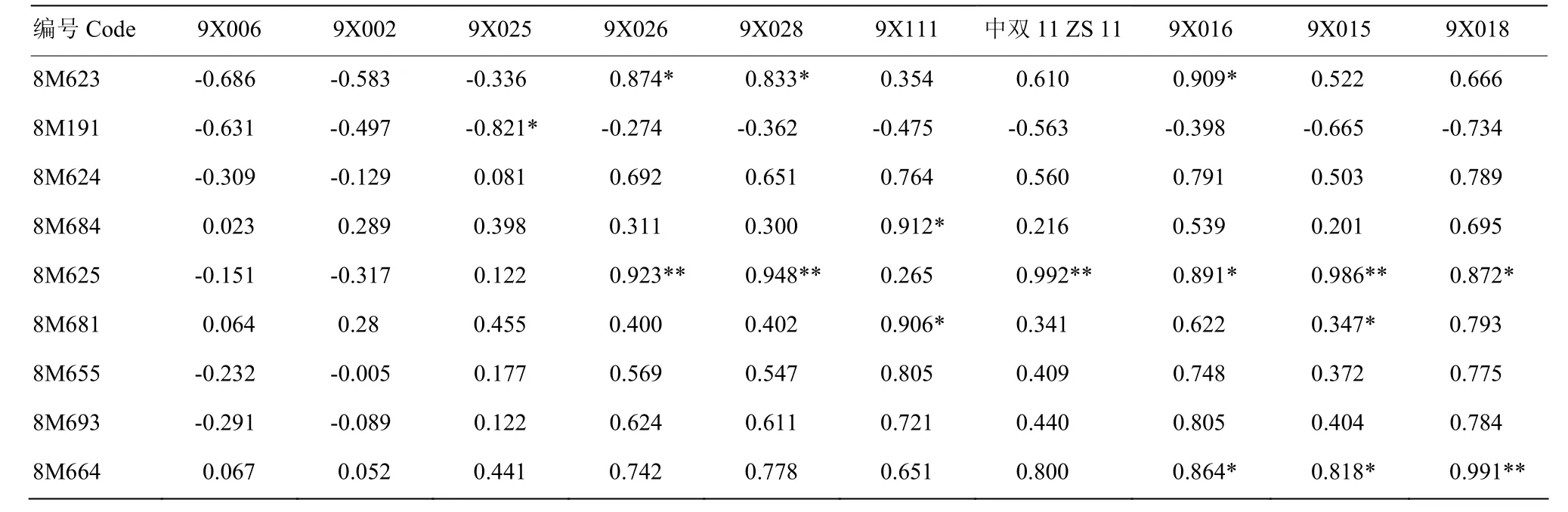
表4 模拟干旱隶属函数值的相关性分析Table 4 Correlation analysis of simulated drought membership function values
3 讨论
甘蓝型油菜是由甘蓝和白菜型油菜杂交,经自然加倍而成。由于其栽培历史短,相对于其亲本种而言,甘蓝型油菜遗传背景较为狭窄[1]。利用甘蓝型油菜亲本种丰富的遗传变异扩展其遗传背景已有不少报道。甘蓝型油菜与甘蓝杂交后不断自交,合成的甘蓝型油菜遗传多样性更丰富[32]。QUAZI等[33]利用甘蓝与甘蓝型油菜杂交,成功将羽衣甘蓝的蚜虫抗性转移到甘蓝型油菜;DING等[34]通过人工合成甘蓝型油菜的途径,将甘蓝亲本的菌核病抗性导入甘蓝型油菜。甘蓝型油菜另一祖先种—白菜型油菜分别独立起源于欧洲和亚洲,具有丰富的遗传变异[18],具备耐贫瘠、耐干旱、抗寒性强、生育期短等特性,为甘蓝型油菜遗传改良提供了丰富的遗传资源[20]。甘蓝型油菜A亚基因组比C亚基因组变异更丰富,白菜型油菜A亚基因组成分对于甘蓝型油菜的遗传变异有很大的影响,其遗传渗入有利于改变甘蓝型油菜遗传背景[16]。利用白菜型油菜与甘蓝型油菜杂交创建新型甘蓝型油菜的策略已在甘蓝型油菜育种中得到应用[35],但此策略中仍需要进行胚挽救,且从后代中选择获得甘蓝型油菜的周期长,效率低。因此,利用其他策略转移白菜型油菜的优良性状到甘蓝型油菜具有重要意义。XIAO等[35]利用埃塞俄比亚芥(B.carinata,BBCC,2n=34)与白菜型油菜杂交,经染色体加倍获得六倍体AABBCC,再与甘蓝型油菜杂交获得五倍体AABCC,而后进行自交,可获得新型甘蓝型油菜。近些年,许多研究者提出了利用六倍体为桥梁改良甘蓝型油菜的方法。钱伟等[27]发明了一种以甘蓝型油菜与甘蓝杂交创建六倍体AnAnCnCnCoCo的方法,并以此六倍体材料为桥梁与大量白菜型油菜杂交,同时导入甘蓝和白菜型油菜的遗传成分,有效拓宽了现有甘蓝型油菜的遗传背景。随后有许多的研究者运用此方法,以六倍体材料为桥梁合成新型甘蓝型油菜。MEI等[28]、刘瑶等[36]利用抗菌核病的甘蓝与甘蓝型油菜杂交获得六倍体,利用此六倍体与甘蓝型油菜回交,创建了菌核病抗性改良的新型甘蓝型油菜。以此六倍体与白菜型油菜杂交,进一步拓宽了甘蓝型油菜遗传背景,且获得了具有菌核病抗性、含白菜型油菜遗传成分的新型甘蓝型油菜[29]。本研究利用的六倍体材料(AnAnCnCnCoCo),前期导入了甘蓝的遗传成分,有效增加了遗传多样性,且以此六倍体与具备优良抗旱性的白菜型油菜(ArAr)杂交,可以快速获得新型甘蓝型油菜AnArCnCo,避免了不断地自交或者回交过程,最大程度快速拓宽了甘蓝型油菜的抗旱性遗传背景,为甘蓝型油菜抗旱性育种提供了优良的种质资源。
植物的抗旱性是由多个基因决定的遗传性状,受基因型和水分胁迫的双重制约[37-39]。卢坤等[40]以RNA-Seq技术在甘蓝型油菜叶片中鉴定了部分干旱胁迫应答基因。CHEN等[41]对盐胁迫响应的甘蓝型油菜叶和根进行比较转录组分析,发现分别有582个和438个转录因子在盐胁迫响应中受到差异调节。ZHANG等[42]鉴定了79个在甘蓝型油菜水分胁迫耐受中起作用的候选基因,其中8个基因与干旱耐受性相关。本研究将白菜的抗旱性进行了有效转移,创建了抗旱性新型甘蓝型油菜,可为甘蓝型油菜抗旱性育种提供优质的种质资源,但是对其分子基础仍不清楚,因此还有待更深入的研究。后续研究中,不仅要深入挖掘白菜型油菜产生抗旱性产生的关键基因,还要深入探讨白菜型油菜A亚基因组的导入对于甘蓝型油菜基因组结构、基因组表达特性产生的深远影响。
4 结论
以六倍体材料为桥梁,与抗旱的白菜型油菜杂交,创制、筛选获得了具有优良抗旱性的新型甘蓝型油菜,可为甘蓝型油菜抗旱育种提供优良的基因资源。
