花生抗黄曲霉大果种质的创制与鉴定
2022-03-16蒋艺飞喻博伦丁膺宾陈伟刚郭建斌陈海文罗怀勇刘念黄莉周小静姜慧芳雷永晏立英康彦平姜成红廖伯寿
蒋艺飞,喻博伦,丁膺宾,陈伟刚,郭建斌,陈海文,罗怀勇,刘念,黄莉,周小静,姜慧芳,雷永,晏立英,康彦平,姜成红,廖伯寿*
(1.中国农业科学院油料作物研究所,湖北 武汉,430062;2.宜昌市农业科学研究院,湖北 宜昌,443004)
花生(Arachis hypogeaL.)是重要的油料和经济作物,广泛种植于世界热带和亚热带地区。中国是世界最大的花生生产和消费国,近十年来花生生产总体持续发展,2018年种植面积达到462万公顷,单产3752 kg/hm2,总产达到1733 万吨,总产量和消费量多年持续居世界首位[1]。然而,花生是一种易受黄曲霉毒素污染的农作物,剧毒且强致癌的黄曲霉毒素(aflatoxins)加大了花生及其制品的食品安全风险,严重威胁花生产业的可持续发展、出口贸易[2,3]和消费者健康[4]。而且,随着花生生产规模的扩大、农业生产(收获)方式的改变、全球气候变暖的加剧,花生黄曲霉毒素污染的风险也不断增加[5,6]。黄曲霉毒素是黄曲霉和寄生曲霉等曲霉属真菌产生的一类次生代谢产物,对人及动物肝脏、肠道的危害极大,可引起急性中毒,也可以诱发癌症[7]。强化黄曲霉毒素污染的治理是农业生产和加工贸易中的重要任务,而培育和种植抗黄曲霉的花生品种是消除或降低毒素污染风险最为经济安全的技术措施。
国内外在花生黄曲霉抗性改良研究方面取得了较大进展,发掘和创制出了具有籽仁侵染抗性、籽仁产毒抗性和荚壳侵染抗性的种质资源和品系。Mixon等通过对100份花生材料的抗性鉴定,筛选到PI337394、PI337409 等抗侵染种质[8];黎穗临等通过对270份花生品种资源的鉴定,筛选到梅县红衣、湛秋48 等抗侵染种质[9];庄伟建等通过对32 份花生品系的鉴定,筛选到闽花6 号、IGCV91284 等抗侵染品系[10];Mehan 等通过对502 份材料的鉴定,筛选到了U-4-7-5、VRR245 等抗产毒种质[11];姜慧芳等通过对716 份核心种质的鉴定,筛选到51002-6、ICG12625等抗产毒种质[12];肖达人等通过对1517份种质的鉴定,筛选到了N1211、N1322 等抗产毒种质[13];邱西克等通过对276 份核心种质的鉴定,筛选到W. h8342、W. h8333 等具有荚壳侵染抗性的种质[14]。同时,初步明确了花生种皮物理结构、荚壳及种皮的完整性与侵染抗性有关[15,16],花生若干生化成分如木犀草素、白藜芦醇等具有抑制产毒的功能[17,18]。但是,国内外花生抗黄曲霉育种的进展总体较为缓慢,培育的抗病品种较少,我国仅有的几个抗黄曲霉花生品种由于荚果小产量潜力低而在生产上应用较少,远远不能满足生产需要。因此,迫切需要发掘抗黄曲霉且具高产潜力的大果花生种质并用于抗性育种。
针对上述问题,本研究以花生高产大果品种中花16 与抗黄曲霉种质J11 杂交构建的RIL 群体为材料,鉴定黄曲霉侵染和产毒抗性,并结合前期对该群体荚果大小的调查结果进行分析,以期获得具有兼抗黄曲霉侵染和产毒的大果新种质,为深化花生黄曲霉抗性改良提供材料支撑。
1 材料与方法
1.1 材料
中花16×J11 重组自交系群体(RIL) F7、F8共188个家系。其中,母本中花16 为感病高产品种,父本J11 为抗病小籽材料。人工接种所用黄曲霉菌株为侵染和产毒能力均强的AF2202[19]。
1.2 方法
RIL 群体及其亲本于2017 和2018 年种植于中国农业科学院油料作物研究所阳逻试验基地,采用常规田间管理。收获晒干后剥壳,选取成熟饱满、大小形状均匀一致、无破损、无病斑、无发芽的籽仁20 粒,浸泡在75%的酒精中3 min,再用无菌水漂洗3 次后进行黄曲霉孢子悬浮液接种,接种浓度为每毫升2×106孢子,接种后振荡使种子表面均匀布满孢子,于30℃培养箱中培养7天后调查侵染指数,依据表面孢子覆盖情况分为四个等级:0、1、2、3 级分别表示孢子覆盖种皮表面积的0、0~25%、25%~50%、50%~100%,计算公式为PSII=(n1+n2×2+n3×3)×100%/(N×3),n1、n2、n3 为各级籽仁数,N 为总粒数。调查侵染指数后的籽仁经酒精消毒后于110℃烘箱中烘60 min,用甲醇提取黄曲霉毒素并用HPLC法检测毒素含量[20]。实验设3次重复。
1.3 数据分析
使用SPSS 26.0 软件计算各性状的平均值、方差、标准差、遗传力及相关性;使用Origin 2018 绘制频率分布图。
2 结果与分析
2.1 花生RIL群体黄曲霉侵染抗性鉴定结果
在2017 年环境中,抗病花生亲本J11 籽仁的侵染指数为56.0%,感病亲本中花16 为87.0%。RIL群体的平均侵染指数为79.4%,变异范围46.9%~98.2%。侵染指数低于50%的家系有3 个,50%~60%的6 个,60%~70%的31 个,70%~80%的55 个,80%~90%的56 个,高于90%的36 个。群体中侵染指数低于抗病亲本J11的家系有5个,高于感病亲本中花16的53个,侵染抗性存在超亲现象。
在2018 年环境中,抗病亲本J11 籽仁的侵染指数为51.3%,感病亲本中花16 为91.1%。RIL 家系的平均侵染指数为76.6%,变异范围28.8%~100.0%。侵染指数低于50%的家系有11 个,50%~60%的16个,60%~70%的22个,70%~80%的48个,80%~90%的41 个,高于90%的37 个。RIL 群体中侵染指数低于抗病亲本J11 的家系11 个,高于感病亲本中花16 的32 个,抗性也表现出超亲现象(图1)。
利用SPSS 26.0 软件对RIL 群体的侵染抗性进行方差分析,家系及年份间差异均达显著水平(P<0.05),广义遗传力为63.74%(表1)。由此表明,RIL群体不同材料之间存在抗性的遗传差异,且抗性受环境影响较大。综合分析两年的鉴定结果,8 份抗黄曲霉侵染种质侵染指数低于58.8%,与J11相当。
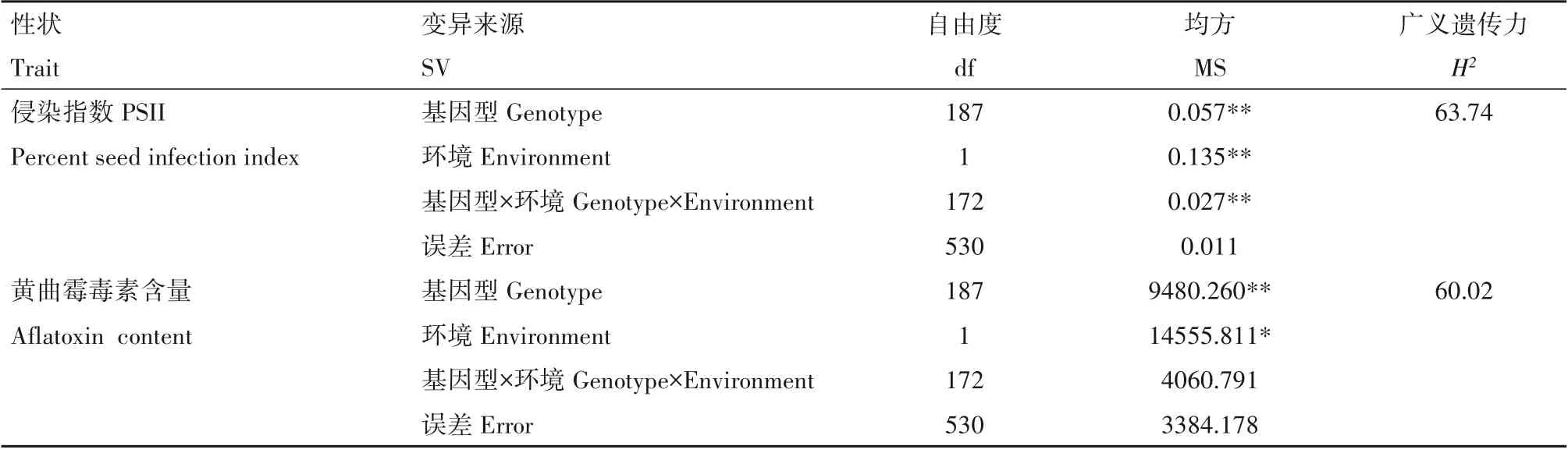
表1 花生RIL群体侵染指数和产毒量的方差及遗传力分析Table 1 Analysis of variance and heritability of PSII and AFT in the peanut RIL
2.2 花生RIL群体的黄曲霉产毒抗性鉴定结果
在2017 年环境中,抗病花生亲本J11 的黄曲霉毒素含量为49.45 μg/g,感病亲本中花16 为158.98 μg/g。 RIL 家系的平均毒素含量138.34 μg/g,变异范围34.82~336.77 μg/g。毒素含量低于80 μg/g 的家系有28 个,80~120 μg/g 的50个,120~160 μg/g 的52 个,160~200 μg/g 的30 个,200~240 μg/g 的15 个,大于240 μg/g 的11 个。群体中黄曲霉毒素含量低于抗病亲本J11的家系有1个,高于感病亲本中花16 的56 个,产毒抗性存在超亲现象。
在2018 年环境中,抗病亲本J11 的毒素含量为76.73 μg/g,感病亲本中花16 为175.67 μg/g。RIL家系的平均毒素含量128.32 μg/g,变异范围39.52~358.84 μg/g。毒素含量低于80 μg/g 的家系有23个,80~120 μg/g 的64 个,120~160 μg/g 的48 个,160~200 μg/g 的26 个,200~240 μg/g 的9 个,大于240 μg/g 的家系5 个。群体中低于抗病亲本J11 的家系19个,高于感病亲本中花16的28个,抗性也表现出超亲现象(图1)。
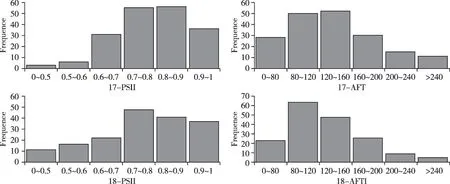
图1 侵染指数和产毒量在2017和2018年的频率分布Fig.1 Frequency distributions of PSII and AFT in 2017 and 2018
利用SPSS 26.0 软件对RIL 群体的产毒抗性进行方差分析,家系及年份间差异均达显著水平(P<0.05),广义遗传力为60.02%(表1)。由此表明,RIL群体不同材料之间存在抗性的遗传差异,且抗性受环境影响较大。综合分析2017 年和2018 年的鉴定结果,获得9份抗黄曲霉产毒种质,毒素含量均低于J11(72.44 μg/g),为高产毒家系的25%以下,具有育种利用价值。
2.3 花生黄曲霉抗性与产量性状的相关性分析
本研究所选用花生RIL群体中的籽仁侵染抗性与产毒抗性显著相关,相关系数为0.193。结合前期[21]RIL 群体的籽仁长(SL)、籽仁宽(SW)及百仁重(HSW)考种数据进行相关性分析(表2),发现PSII和AFT 均与SL、SW 及HSW 显著正相关,其中PSII与三个籽仁性状的相关系数分别为0.284、0.181、0.290,AFT 与三个籽仁性状的相关系数分别为0.249、0.178、0.268。结果表明,一般情况下抗侵染的材料也具有产毒抗性,抗侵染和抗产毒的材料籽仁较小。
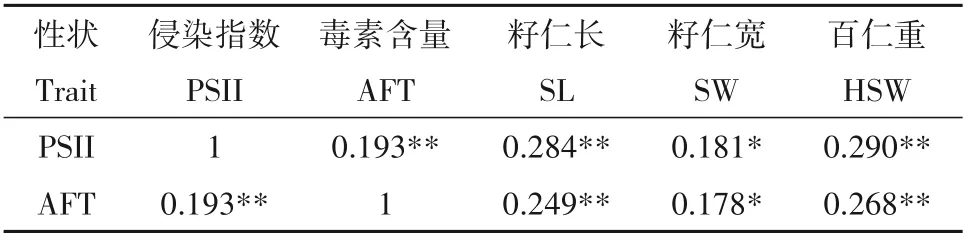
表2 花生黄曲霉抗性与产量性状间的相关性Table 2 Correlation between resistance to A.flavus and yield-related traits
对上述筛选出的抗侵染和抗产毒材料的荚果及籽仁大小分析,百果重平均138.23 g,变异范围为110.65~196.94 g,百仁重平均58.48 g,变异范围为46.68~83.36 g,百果重在150 g 以下的有12 份,190 g 以上的仅1 份,百仁重在60 g 以下的有12 份,80 g 以上的仅1 份,可见这些抗性材料多集中于小果小籽(表3)。其中,QT1107 为兼抗侵染和产毒的大果新种质,百果重达196.94 g,百仁重达83.36 g,达到大果型花生品种的百果重标准,可作为后续抗性育种材料使用。
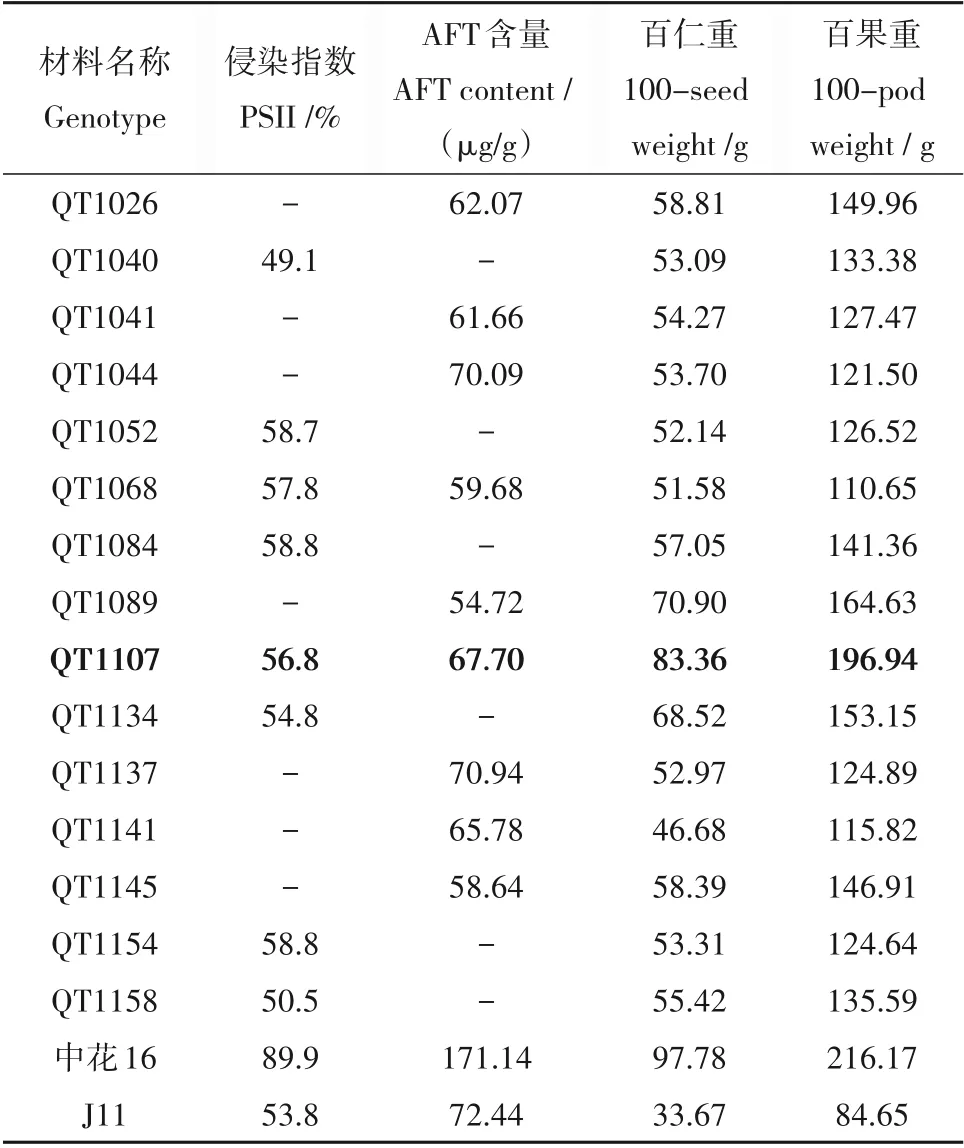
表3 抗黄曲霉花生种质的产量相关性状Table 3 Yield-related traits of peanut lines with resistance to A.flavus
3 讨论与结论
黄曲霉毒素污染严重影响花生的产量和品质,是花生产业可持续发展面临的重大问题。培育抗黄曲霉花生品种是防控毒素污染最经济有效的措施,但目前生产上抗性品种应用有限,原因是抗病亲本遗传基础狭窄,抗源种质综合表现尤其是产量潜力较差。本研究通过对源自国际公认的抗黄曲霉花生种质J11 的重组自交系的两年表型数据分析,发现花生侵染抗性和产毒抗性间存在一定的相关性,并鉴定获得了兼抗侵染和产毒的大果新种质,为花生黄曲霉抗性育种提供了潜在的抗源材料。
花生抗黄曲霉种质的筛选多集中于籽仁的侵染和产毒,也有对荚壳侵染抗性的筛选,但并未见对侵染和产毒综合进行筛选的相关报道。黄曲霉抗性鉴定需要较多种子材料,且在低世代群体中进行筛选易丢失高抗重组类型,加之现有的抗性种质鲜有兼抗侵染和产毒的,在很大程度上限制了花生黄曲霉抗性不同机制的系统研究和聚合。关于种质筛选的开展,廖伯寿等利用RIL7-9群体成功创制出3份含油量高、农艺性状优良、高抗青枯病的花生新种质,李威涛等利用F5-F7代RIL 群体筛选出6 份抗青枯病、荚果大和出仁率高的花生新种质[22,23]。本研究通过鉴定中花16×J11 RIL 群体F7-F8代侵染抗性与产毒抗性,共筛选出抗侵染材料8份,抗产毒材料9 份,其中QT1068、QT1107 为兼抗侵染和产毒的材料。这些抗性材料中,QT1040侵染指数比抗病亲本J11 低8 个百分点,QT1158 低6 个百分点,QT1026、QT1041、QT1068、QT1089、QT1145 毒素含量均比J11 低10 个百分点以上,侵染抗性和产毒抗性均表现出超亲现象,表明通过不同抗性品系的杂交,可获得优良抗性育种材料。本研究中抗病亲本J11 为国际公认的抗侵染花生种质,推测由于高抗侵染使得黄曲霉菌无法定殖于花生内部从而无法获得营养物质,最终产生少量黄曲霉毒素,这一机理有待进一步研究。
通过对籽仁侵染指数、毒素含量和籽仁大小的相关性分析,三者间存在一定程度的正相关,本研究中的抗病亲本J11 侵染指数低、产毒量低,但籽仁小,产量性状指标过低,多年鉴定百果重约84.65g、百仁重约33.08g。若与经济性状好的大果杂交组合则更易在后代家系中创制出大果材料。前人研究发现花生黄曲霉抗性和荚果大小为数量性状,受多基因控制[24,25],而本研究中感病亲本中花16 为高产品种,二者荚果大小差异倍数为2.55 倍,侵染指数1.67 倍,产毒量2.36 倍,可见两亲本的遗传背景有较大差异,是聚合创制抗黄曲霉的大果种质的良好组合。结合对已筛选的抗黄曲霉种质的荚果大小进行考察,成功在抗性稳定的高世代RIL 家系中鉴定出兼抗侵染和产毒的优良大果新种质。
连续两年对高世代花生RIL群体进行黄曲霉侵染抗性和产毒抗性的表型鉴定,获得抗侵染种质8份,抗产毒种质9 份,其中兼抗侵染和产毒种质2份,兼抗侵染和产毒的大果种质1份。
