ENU诱变草鱼家系生长特性及肌肉生长抑制素基因SNP筛选的研究
2020-03-20王成龙蒋霞云邹曙明
王成龙,陈 杰,蒋霞云,邹曙明
( 上海海洋大学,农业农村部淡水水产种质资源重点试验室,上海 201306 )
N-乙基-N-亚硝基脲(ENU)是一种常用的化学诱变剂,它能对基因组DNA碱基产生烷基化修饰,使DNA在复制时发生错配,导致突变体生成[1-2]。ENU主要诱发不定点的单碱基突变,无任何倾向性,从而使基因发生突变,其诱变模式与自然变异相似[3-4]。同时,ENU诱变效率很高,约为其他诱变手段的10倍。在模式生物中,利用ENU获得大规模突变体进行功能基因的研究已成为常用手段。但在养殖鱼类中的应用报道很少。目前,仅前苏联采用ENU诱变结合雌核发育技术获得了鲤鱼优良突变品系[5]。国内方面,韩启霞等[6]采用ENU浸泡的方法诱变银鲫(Carassiusauratusgibelio)得到突变个体,邹曙明等[7]用ENU诱变处理草鱼(Ctenopharyngodonidellus)精子,产生了生长速度显著不同的F1代。
草鱼肌肉生长抑制素(MSTN)基因是一种肌细胞生长负向调控因子,具有抑制肌肉分化和生长的作用,其表达量与肌肉质量的变化呈负相关[8-10]。目前,针对MSTN基因在哺乳动物方面的功能及调控机理研究较多[11-15],在水产动物方面其研究还处于cDNA克隆与表达阶段,功能研究相对较少[16-19]。鱼类MSTN基因存在两种亚型(MSTN1、MSTN2)并且时空表达具有较大的差异。濮剑威等[20]研究表明,MSTN1基因在草鱼肌肉、脑和眼中的转录量较高,MSTN2基因只在脑和肌肉中有表达。并且通过注射MSTN1型mRNA过表达可导致斑马鱼(Daniorerio)体节发生期胚胎的前后轴拉长、背腹轴变短、脊索轻微扭曲及体节发育受到强烈抑制而不分化等现象。注射MSTN2型 mRNA胚胎早期发育有所延迟并未发生明显变化,但发育至60 h之后尾部明显发生严重弯曲。
单核苷酸多态性(SNP)是指在基因组水平上由于单个核苷酸的变异所引起的DNA序列多态性。SNP作为第三代分子遗传标记在分子育种方面已经得到广泛的应用[21-22]。MSTN基因的SNPs与生长性状的关联研究在畜禽方面已有大量的报道[23-25],在水产动物方面,MSTN1基因在黄颡鱼(Pelteobagrusefulvidraco)[26]、建鲤(Cyprinuscarpiovar.jian)[27]、吉富罗非鱼(OreochromisniloticusGIFT)[28]、草鱼[29]等可作为育种的候选分子标记,而MSTN2基因SNPs与生长性状的关联研究未见报道。笔者通过研究ENU诱变草鱼4个家系的生长,对比筛选出2个具有显著生长差异的家系,通过双向测序在这2个家系中MSTN1、MSTN2基因找出SNP位点,为ENU诱变草鱼生长性状的分子标记辅助育种提供候选标记。
1 材料与方法
1.1 试验材料及管理
ENU诱变草鱼亲本来自于上海海洋大学农业农村部团头鲂(Megalobramaamblycephala)育种中心。挑取6龄优良ENU诱变草鱼亲本进行人工繁殖,雌、雄草鱼各4尾,每尾约25 kg。对亲本进行一雌一雄交配方式干法授精,建立4个ENU诱变草鱼家系,将每个家系的受精卵分别放至孵化桶里进行孵化,待受精卵孵化成平游的鱼苗后,取每个家系鱼苗约2000尾放入6 m×4 m×1.5 m的水泥池中暂养1个月后,再将每个家系鱼苗平均分到2个水泥池中暂养65 d,从鱼苗平游开始计算,第80 d通过剪鳍标记法对家系1、家系2、家系3和家系4分别剪左胸鳍、不剪鳍、右胸鳍和左腹鳍加以区分,从每个家系中随机挑选50尾称量体质量后同池混养,设置3个平行试验组,约每隔30 d对每个家系补剪鳍后放回原水泥池,经过175 d饲养,在每个水泥池各个家系中随机挑选20尾进行生长指标测量,测量完将4个家系草鱼放到同一个土塘进行饲养,放塘之前每个家系各取10尾鱼鳍浸于95%酒精中,-20 ℃保存备用。
1.2 生长指标测量
用电子称测量草鱼体质量(精确到0.01 g);用直尺、圆规测量全长、体长、头长、体高、尾柄长、尾柄高、体厚形态性状(精确至0.1 cm)。
1.3 基因组DNA提取及SNPs筛选
参照北京天根生物科技有限公司的海洋动物组织基因组DNA提取试剂盒(离心柱型)说明书提取样品基因组DNA。基因组DNA提取完成后,用0.8%的琼脂糖凝胶电泳和分光光度计检测DNA质量和含量,-20 ℃保存备用。
根据美国国立生物技术信息中心GenBank数据库公布的草鱼MSTN1、MSTN2基因序列,采用Primer Premier 5软件设计MSTN1、MSTN2基因所需引物,基因所用引物由生工生物工程(上海)股份有限公司合成,所用引物序列及长度见表1。PCR反应体系为50 μL:模板DNA 60~100 ng,10×Taq Buffer 5μL,dNTP Mixture(2.5 mmol/L) 2 μL,Taq酶(2.5 U/μL)1 μL,上、下游引物(10 μmol/L)各1 μL,0.1% DEPC处理过的去离子水补至50 μL。PCR反应条件为:94 ℃预变性5 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30~90 s,35个循环;72 ℃后延伸10 min,4 ℃终止反应。取3 μL PCR扩增产物,经0.8%琼脂糖凝胶电泳检测PCR产物,电泳条带符合送测要求后,由生工生物工程(上海)股份有限公司进行双向测序。测序的结果利用BioEdit软件确定SNP位点(突变率高于30%)。

表1 ENU诱变草鱼MSTN1、MSTN2基因部分区域扩增SNP位点筛选所用的引物Tab.1 Primers used for screening SNP loci in partial regions of MSTN1 and MSTN2 genes in ENU mutagenized grass carp
1.4 数据统计与分析
使用SPSS 19.0软件[21]中一般线性模型对4个ENU诱变草鱼家系的全长、体长、体高、头长、尾柄长、尾柄高、体厚等性状进行相关性分析。运用美国国立生物技术信息中心、BioEdit对MSTN1、MSTN2基因中SNPs位点及氨基酸翻译进行比对。
2 结 果
2.1 ENU诱变草鱼4个家系生长性状对比
第80 d,平均体质量为家系1>家系4>家系3>家系2,绝对质量增加率为家系1>家系4>家系3>家系2;第175 d,平均体质量为家系4>家系1>家系2>家系3,绝对体质量增加率为家系4>家系2>家系1=家系3(表2)。
4个ENU诱变草鱼家系在第175 d各生长性状情况见表3,除家系1和家系2的体高和头长差异不显著外,其他不同阶段每两个家系间的各个性状均差异显著。

表2 4个ENU诱变草鱼家系不同阶段体质量的增长统计Tab.2 Growth statistics of body weight in four ENU mutagenized grass carp families with different stages

表3 4个ENU诱变草鱼家系各生长性状特征值Tab.3 Characteristics of growth traits of four ENU mutagenized grass carp families
注:同一行数字后的字母不同表示差异显著(P<0.05).
Note: means with different letters in the same line are significant differences(P<0.05).
对4个ENU诱变草鱼家系在第175 d各形态性状与体质量的相关程度进行了偏相关分析,结果见表4,各形态性状与质量显著性相关由P<0.01判断,因此各家系中与体质量显著性相关的性状分别为:家系1中有全长、体长、尾柄长、体厚,相关系数依次为0.357、0.619、0.608、0.396;家系2中有全长、体长、头长、体厚,相关系数依次为0.348、0.360、0.687、-0.384;家系3中有全长、体长、尾柄长、尾柄高,相关系数依次为0.529、0.449、-0.351、0.384;家系4中有全长、体长、体厚,相关系数依次为0.629、0.543、0.590。
对4个ENU诱变草鱼家系在第175 d的全长、体高、头长、尾柄长、尾柄高、体厚、体质量与体长的比例性状进行了因子分析。通过KMO和Bartlett检验其度量值为0.705接近1,因此适合因子分析,其中第一主成分(P1)和第二主成分(P2)的贡献率分别为39.11%、18.98%,累计贡献率为58.09%,表达式分别为:
P1=-0.113x1+0.376x2+0.436x3+0.228x4+0.006x5+0.335x6-0.033x7;
P2=0.368x1-0.014x2-0.372x3+0.065x4+0.400x5+0.053x6+0.408x7。
式中,x1为全长/体长、x2为体高/体长、x3为头长/体长、x4为尾柄长/体长、x5为尾柄高/体长、x6为体厚/体长、x7为体质量/体长。
各比例性状的分布见图1,7个比例性状可分成两组,第一主成分主要为体高/体长、头长/体长、尾柄长/体长、体厚/体长,第二主成分主要为全长/体长、尾柄高/体长、体质量/体长。
每个家系个体根据第一主成分和第一主成分相关系数得分绘制散点图(图2),家系4在第一主成分、第一主成分得分区域分布明显与其他3个家系不同,家系2和家系3大部分个体分布重叠,家系1与家系2、家系3都有少部分个体质量重叠。
注:P<0.01表示极其显著.
Note:P<0.01 means very significantly different.

图1 比例性状因子主成分得分系数分布Fig.1 The coefficient distribution map of principal component score of proportional trait factor
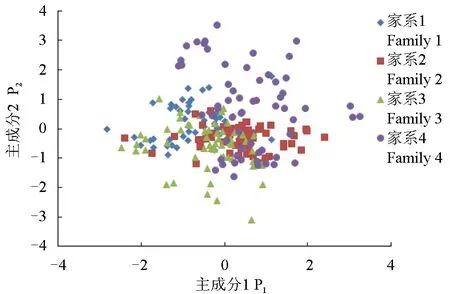
图2 4个ENU诱变草鱼家系的各比例性状特征主因子散点示意Fig.2 The scatter plot map of proportional trait factor in four ENU mutagenized grass carp families
2.2 MSTN1、MSTN2基因序列分析及SNPs位点筛选
本试验获得草鱼MSTN1基因全长共3824 bp,包括2个内含子(长度分别为776、1008 bp)和3个外显子(长度分别为375、370、382 bp),其GenBank为KP719016;对于MSTN2的mRNA全长共1976 bp,其GenBank为KM874827.1。对MSTN基因进行双向测序,碱基在家系内出现2次以上(包括2次)即确定为SNP突变位点。
对于MSTN1基因,家系3、4均在465位点C/G发生错义突变,导致该位点编码的氨基酸由脯氨酸变为精氨酸,家系3还在467位点G/A均发生错义突变,导致该位点编码的氨基酸由天冬氨酸变为天冬酰胺(图3)。
对于MSTN2基因,家系3、4均在912位点C/T发生同义突变,编码的氨基酸为络氨酸,另外家系3在1027位点G/A产生错义突变,导致该位点编码的氨基酸由缬氨酸变为异亮氨酸,而家系4在366位点A/G产生错义突变,导致该位点编码的氨基酸由谷氨酰胺变为精氨酸(图4)。而位于非编码区1390位点A/T和1401位点G/A发现SNP位点(图4)。
3 讨 论
3.1 生长对比设计
在生长对比试验中,试验对象初始规格的差异可以影响到整个试验结束的结果[30],为减少前期生长差异对后期试验的影响,亲鱼催产、人工授精、受精卵孵化、鱼苗放到水泥池暂养的整个过程均同步进行,到了第80 d对每个ENU诱变草鱼家系随机挑取50尾进行剪鳍标记同池混养,设置3个平行试验组,克服不同水泥池对生长对比造成的影响。在整个饲养管理过程中,每日定时投喂2次,每次以水面留有少量饲料为准,每次测量生长性状的操作是在室内进行,并且提前准备足够的冰水,尽量减少高温对草鱼的损伤。鱼类标记一般常用的方法有挂牌、剪鳍、染料标记、荧光标记、电子标记等标记技术,本试验使用的剪鳍标记,相对于其他标记有操作简单、保留时间持久、对鱼伤害小等优点,李思发等[31]研究了剪鳍标记部位对吉富罗非鱼生长的影响,指出剪左胸、右胸、左腹和右腹鳍标记的鱼之间以及剪不同标记的鱼与未剪鳍标记的鱼之间的生长无显著差异。


图3 家系3和家系4 MSTN1基因部分序列SNP位点筛选Fig.3 SNP site selection of partial sequence of MSTN1 in family 3 and family 4

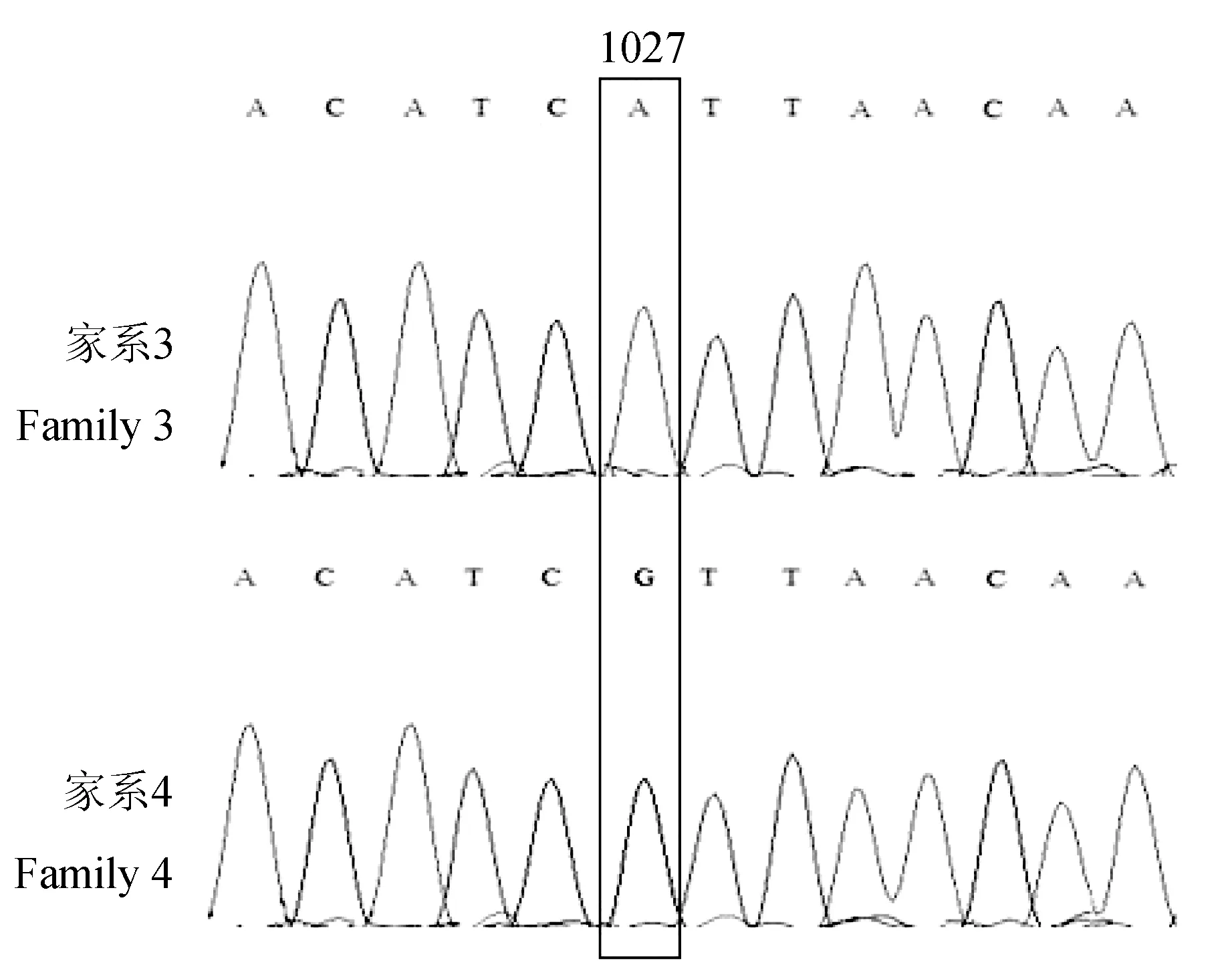
图4 家系3和家系4 MSTN2基因部分序列SNP位点筛选Fig.4 SNP site selection of partial sequence of MSTN2 in family 3 and family 4
3.2 家系生长差异分析
从平均体质量和绝对质量增加率来看,经过175 d的养殖,家系4在这两方面具有明显的优势,而家系3生长优势最弱,从4个家系各个生长性状对比可知,家系4明显处于优势地位。从各个性状与质量的相关程度的角度分析,每个家系的主要相关性状有所区别,家系1中相关系数最高的是尾柄长(0.619),其次是体长(0.619);家系2中相关系数最高的是头长(0.687),明显高于全长、体长、体厚;家系3中相关系数最高的是全长(0.529);家系4中全长、体长、体厚的相关系数相近。形态比例性状在鱼类分类中是一个很重要指标,庆宁等[32]通过形态比例性状对华南沿海地区西部不同水系的中间黄颡鱼(P.intermedius)群体进行了有效分类。本研究通过因子分析将多个相关的形态指标转换为新的、个数较少且相互独立的综合指标,通过4个家系个体在第一、二主成分因子得分系数图可以有效区分家系4和家系3,由此可以看出家系4的生长性状相对于家系3具有明显的生长差异。
3.3 MSTN基因在ENU诱变草鱼家系的SNP多态性
ENU溶液浸泡精子可在草鱼中实现高效诱变,在诱变育种领域具有潜在的应用前景。前期研究显示,ENU主要诱变草鱼基因组DNA产生GC到AT,或者AT到GC的点突变,可造成密码子的无义突变、错义突变和同义突变,ENU对上述草鱼基因组的平均诱变率为0.41%[7]。本试验选取的4个ENU诱变草鱼家系均是具有显著生长优势的个体。通过对MSTN1、MSTN2基因初筛选出不同的SNP位点,对于MSTN1基因,家系3和家系4均在465位点C/G发生错义突变,家系3还在467位点G/A发生错义突变。对于MSTN2基因,家系3、4均在912位点C/T发生同义突变,家系3在1027位点G/A产生错义突变,而家系4在366位点A/G产生错义突变。由此推测,家系3和家系4产生的显著生长差异,与MSTN基因的SNP位点差异导致编码的氨基酸不同,使该基因功能表达差异有联系,因此家系3和家系4可作为试验材料来研究MSTN基因相关分子标记的开发,为后续的ENU诱变草鱼的分子标记辅助育种奠定基础。
